
Effects of TaMTL-Edited Mutations on Grain Phenotype and Storage Component Composition in Wheat


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wheat Materials and Planting
2.2. Embryo Rescue from Endospermless Grains
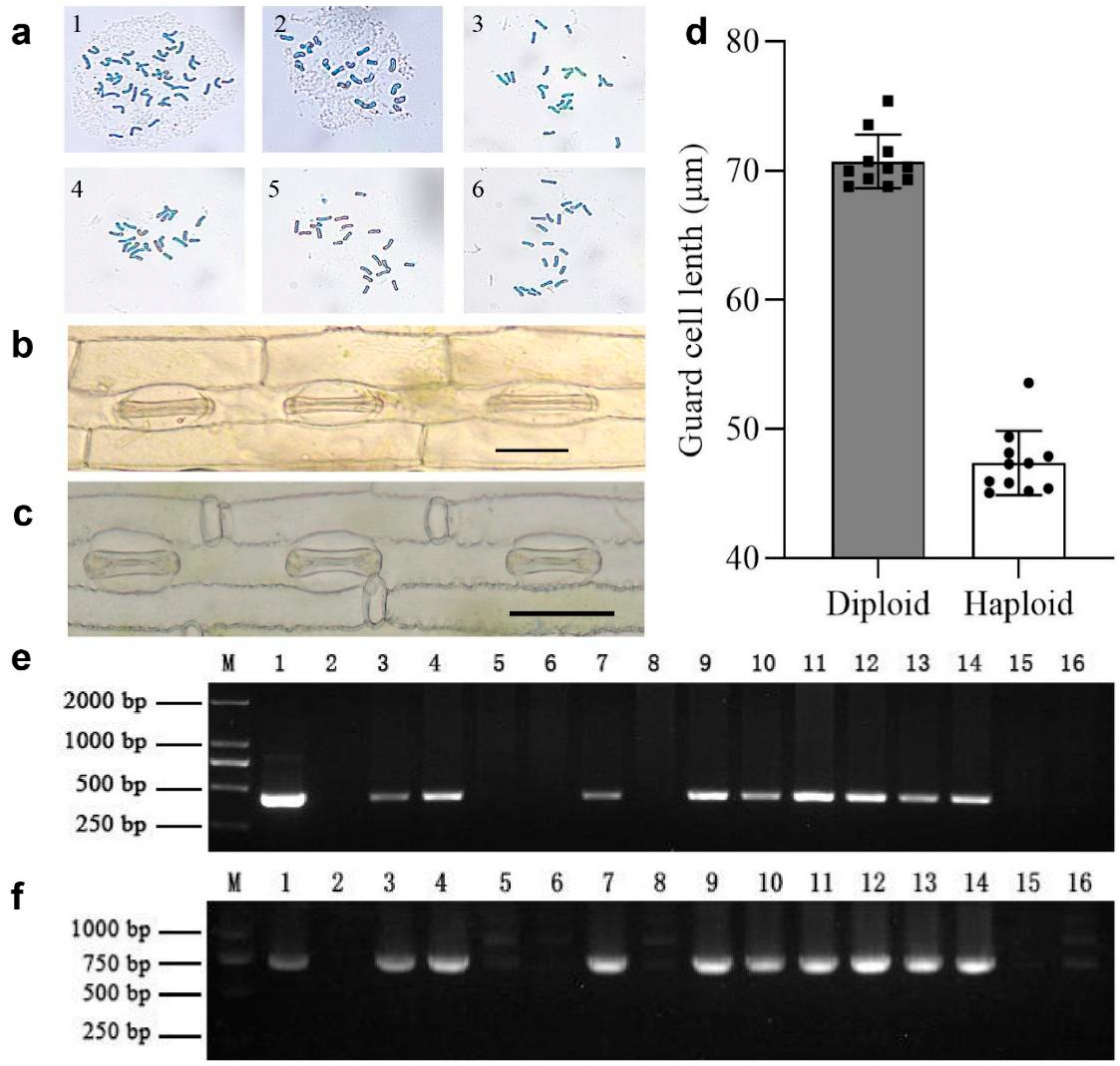
2.3. Determination of Wheat Guard Cell Length
2.4. Chromosome Counting in Wheat Root Tips
2.5. Marker-Assisted Haploid Confirmation in Hybrid F1 Seedlings
2.6. Grain Trait Examination of the mtl Mutants
2.7. Observation of Starch Granules and Protein Bodies in Wheat Grains
2.8. Determination of Total Starch and Amylose Contents
2.9. Investigation of Starch Granule Morphology
2.10. Quantification of the Gluten and Non-Gluten Contents
2.11. Statistical Analysis
3. Results
3.1. Induction of Wheat Haploid Seeds in mtl Mutants
3.2. Wheat Grain Measurments in mtl Mutants
3.3. Wheat Embryo Abortion in the mtl-ABD Mutant
3.4. Effect of Embryo Abortion on Endosperm Structure
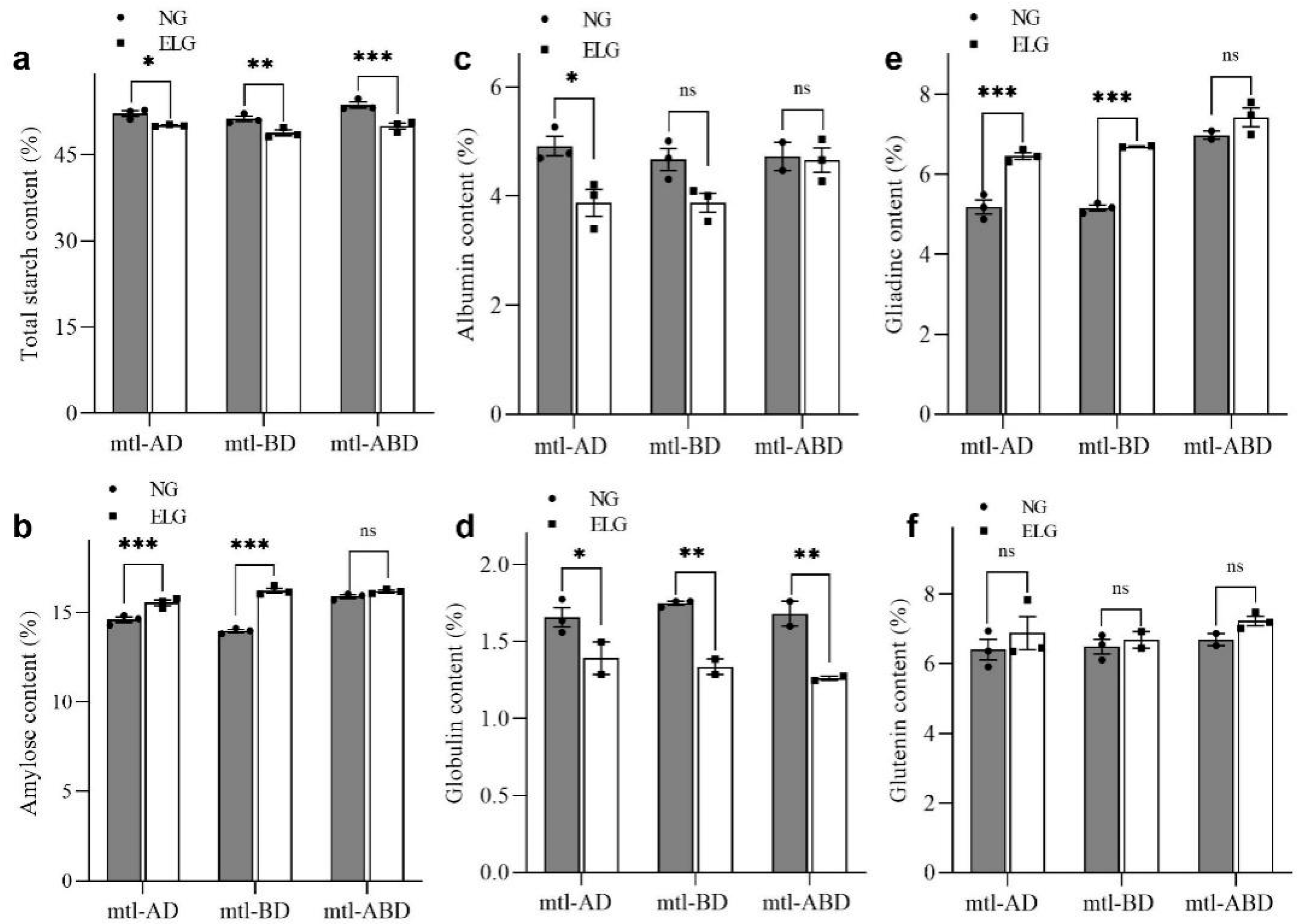
3.5. Effect of Wheat Embryo Abortion on Starch Content and Starch Granule Composition
3.6. Effect of Wheat Embryo Abortion on Grain Storage Protein Composition
4. Discussion
4.1. The mtl-A Mutation Has No Effect on Wheat Grain Phenotype
4.2. Independence between Wheat Embryo and Endosperm Development
4.3. Endosperm Origin in Wheat Embryoless Grains of the mtl Mutants
4.4. Rapid Screening of the Wheat Haploids Induced by the mtl Mutations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Berger, F.; Hamamura, Y.; Ingouff, M.; Higashiyama, T. Double fertilization-caught in the act. Trends Plant Sci. 2008, 13, 437–443. [Google Scholar] [PubMed]
- Evers, T.; Millar, S. Cereal Grain Structure and Development: Some Implications for Quality. J. Cereal Sci. 2002, 36, 261–284. [Google Scholar] [CrossRef]
- Sabelli, P.A.; Larkins, B.A. The Development of Endosperm in Grasses. Plant Physiol. 2009, 149, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, I.; Raff, M. Size Control in Animal Development. Cell 1999, 96, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Crickmore, M.A.; Mann, R.S. The control of size in animals: Insights from selector genes. BioEssays 2008, 30, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations (FAO). World Food Situation: FAO Cereal Supply and Demand Brief; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2016. [Google Scholar]
- Barron, C.; Surget, A.; Rouau, X. Relative amounts of tissues in mature wheat (Triticum aestivum L.) grain and their carbohydrate and phenolic acid composition. J. Cereal Sci. 2007, 45, 88–96. [Google Scholar] [CrossRef]
- Hurkman, W.J.; McCue, K.; Altenbach, S.B.; Korn, A.; Tanaka, C.K.; Kothari, K.M.; Johnson, E.L.; Bechtel, D.B.; Wilson, J.D.; Anderson, O.D.; et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 2003, 164, 873–881. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins-structures and biosynthesis. Plant Cell. 1995, 7, 945–956. [Google Scholar]
- Ball, S.G.; Morell, M.K. From Bacterial Glycogen to Starch: Understanding the Biogenesis of the Plant Starch Granule. Annu. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Botticella, E.; Sestili, F.; Hernandez-Lopez, A.; Phillips, A.; Lafiandra, D. High resolution melting analysis for the detection of EMS induced mutations in wheat SbeIIa genes. BMC Plant Biol. 2011, 11, 156. [Google Scholar]
- Li, J.; Jiao, G.; Sun, Y.; Chen, J.; Zhong, Y.; Yan, L.; Xia, L. Modification of starch composition, structure and properties through editing of TaSBEIIa in both winter and spring wheat varieties by CRISPR/Cas9. Plant Biotechnol. J. 2021, 19, 937–951. [Google Scholar]
- Evers, A.D. Scanning Electron Microscopy of Wheat Starch. III. Granule Development in the Endosperm. Starch 1971, 23, 157–162. [Google Scholar] [CrossRef]
- Evers, A.D. The Size Distribution Among Starch Granules in Wheat Endosperm. Starch 1973, 25, 303–304. [Google Scholar] [CrossRef]
- Morrison, W.; Gadan, H. The amylose and lipid contents of starch granules in developing wheat endosperm. J. Cereal Sci. 1987, 5, 263–275. [Google Scholar] [CrossRef]
- Shevkani, K.; Singh, N.; Bajaj, R.; Kaur, A. Wheat starch production, structure, functionality and applications-a review. Int. J. Food Sci. Technol. 2016, 52, 38–58. [Google Scholar] [CrossRef]
- Faridi, H.; Finley, J.W.; D’Appolonia, B. Improved wheat for baking. Crit. Rev. Food Sci. Nutr. 1989, 28, 175–209. [Google Scholar] [CrossRef]
- Fiaz, S.; Sheng, Z.; Zeb, A.; Barman, H.N.; Shar, T.; Ali, U.; Tang, S. Analysis of genomic regions for crude protein and frac-tions of protein using a recombinant inbred population in rice (Oryza sativa L.). J. Taibah Univ. Sci. 2021, 15, 579–588. [Google Scholar]
- Snyder, H. The Proteins of the Wheat Kernel; American Association for the Advancement of Science: Washington, DC, USA, 1907. [Google Scholar]
- Dupont, F.M.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Altenbach, S.B. Deciphering the complexities of the wheat flour proteome using quantitative two-dimensional electrophoresis, three proteases and tandem mass spectrometry. Proteome Sci. 2011, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Loussert, C.; Popineau, Y.; Mangavel, C. Protein bodies ontogeny and localization of prolamin components in the developing endosperm of wheat caryopses. J. Cereal Sci. 2008, 47, 445–456. [Google Scholar] [CrossRef]
- Vensel, W.H.; Tanaka, C.K.; Altenbach, S.B. Protein composition of wheat gluten polymer fractions determined by quantitative two-dimensional gel electrophoresis and tandem mass spectrometry. Proteome Sci. 2014, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Tao, H.P.; Kasarda, D.D. Two-Dimensional Gel Mapping and N-Terminal Sequencing of LMW-Glutenin Subunits. J. Exp. Bot. 1989, 40, 1015–1020. [Google Scholar] [CrossRef]
- Bean, S.R.; Lookhart, G.L. Factors Influencing the Characterization of Gluten Proteins by Size-Exclusion Chromatography and Multiangle Laser Light Scattering (SEC-MALLS). Cereal Chem. 2001, 78, 608–618. [Google Scholar] [CrossRef]
- Don, C.; Mann, G.; Bekes, F.; Hamer, R. HMW-GS affect the properties of glutenin particles in GMP and thus flour quality. J. Cereal Sci. 2006, 44, 127–136. [Google Scholar] [CrossRef]
- Shewry, P.R.; D’Ovidio, R.; Lafiandra, D.; Jenkins, J.A.; Mills, E.C.; Békés, F. Wheat grain proteins. In Wheat: Chemistry and Technology; American Association of Cereal Chemists: St. Paul, MN, USA, 2009; pp. 223–298. [Google Scholar]
- Kelliher, T.; Starr, D.; Richbourg, L.; Chintamanani, S.; Delzer, B.; Nuccio, M.L.; Green, J.; Chen, Z.; McCuiston, J.; Wang, W.; et al. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature 2017, 542, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, X.; Meng, D.; Zhong, Y.; Chen, C.; Dong, X.; Xu, X.; Chen, B.; Li, W.; Li, L.; et al. A 4-bp Insertion at ZmPLA1 Encoding a Putative Phospholipase A Generates Haploid Induction in Maize. Mol. Plant 2017, 10, 520–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Zhang, Y.; Liu, C.; Liu, Y.; Wang, Y.; Liang, D.; Liu, J.; Sahoo, G.; Kelliher, T. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants 2018, 4, 530–533. [Google Scholar] [CrossRef]
- Liu, C.; Zhong, Y.; Qi, X.; Chen, M.; Liu, Z.; Chen, C.; Tian, X.; Li, J.; Jiao, Y.; Wang, D.; et al. Extension of the in vivo haploid induction system from diploid maize to hexaploid wheat. Plant Biotechnol. J. 2019, 18, 316–318. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.Y.; Wang, K.; Jia, Z.M.; Gong, Q.; Lin, Z.S.; Du, L.P.; Ye, X. Editing TaMTL gene induces haploid plants efficiently by optimized Agrobac-terium-mediated CRISPR system in wheat. J. Exp. Bot. 2020, 71, 1337–1349. [Google Scholar]
- Du, L.P.; Xu, H.J.; Zhao, L.L.; Zhang, C.X.; Ye, X.G. Identification of ploidy of wheat pollen plants using length of guard cell. Beijing Agric. Sci. 1996, 14, 10–12. (In Chinese) [Google Scholar]
- Li, S.; Wang, J.; Wang, K.; Chen, J.; Wang, K.; Du, L.; Ye, X. Development of PCR markers specific to Dasypyrum villosum genome based on transcriptome data and their application in breeding Triticum aestivum-D. villosum#4 alien chromosome lines. BMC Genom. 2019, 20, 289. [Google Scholar]
- Yuan, J.; Guo, X.; Hu, J.; Lv, Z.; Han, F. Characterization of two CENH 3 genes and their roles in wheat evolution. New Phytol. 2014, 206, 839–851. [Google Scholar] [CrossRef]
- Zhu, J.; Fang, L.; Yu, J.; Zhao, Y.; Chen, F.; Xia, G. 5-Azacytidine treatment and TaPBF-D over-expression increases glutenin accumulation within the wheat grain by hypomethylating the Glu-1 promoters. Theor. Appl. Genet. 2018, 131, 735–746. [Google Scholar]
- Wang, H.; Yang, J.; Zhang, M.; Fan, W.; Firon, N.; Pattanaik, S.; Yuan, L.; Zhang, P. Altered Phenylpropanoid Metabolism in the Maize Lc-Expressed Sweet Potato (Ipomoea batatas) Affects Storage Root Development. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Zhou, Q.; Li, E.; Yuan, L.; Wang, W.; Zhang, H.; Liu, L.; Wang, Z.; Yang, J.; Gu, J. Effects of nitrogen fertilizer on structure and physicochemical properties of ‘super’ rice starch. Carbohydr. Polym. 2020, 239, 116237. [Google Scholar] [CrossRef]
- Zheng, T.; Qi, P.-F.; Cao, Y.-L.; Han, Y.-N.; Ma, H.-L.; Guo, Z.-R.; Wang, Y.; Qiao, Y.-Y.; Hua, S.-Y.; Yu, H.-Y.; et al. Mechanisms of wheat (Triticum aestivum) grain storage proteins in response to nitrogen application and its impacts on processing quality. Sci. Rep. 2018, 8, 11928. [Google Scholar] [CrossRef]
- Xu, X.; Li, L.; Dong, X.; Jin, W.; Melchinger, A.E.; Chen, S. Gametophytic and zygotic selection leads to segregation dis-tortion through in vivo induction of a maternal haploid in maize. J. Exp. Bot. 2013, 64, 1083–1096. [Google Scholar]
- Neuffer, M.G.; Sheridan, W.F. Defective kernel mutants of maize. I. genetic and lethality studies. Genetics 1980, 95, 929–944. [Google Scholar] [CrossRef]
- Kranz, E.; Von Wiegen, P.; Quader, H.; Lörz, H. Endosperm development after fusion of isolated, single maize sperm and central cells in vitro. Plant Cell. 1998, 10, 511–524. [Google Scholar]
- Ohad, N.; Margossian, L.; Hsu, Y.C.; Williams, C.; Repetti, P.; Fischer, R.L. A mutation that allows endosperm development without fertilization. Proc. Natl. Acad. Sci. USA 1996, 93, 5319–5324. [Google Scholar] [CrossRef] [Green Version]
- Grossniklaus, U.; Vielle-Calzada, J.P.; Hoeppner, M.A.; Gagliano, W.B. Maternal control of embryogenesis by medea, a Polycomb group gene in Arabidopsis. Science 1998, 280, 446–450. [Google Scholar]
- Ohad, N. Plant development: Parental conflict overcome. Nature 2007, 447, 275–276. [Google Scholar] [CrossRef]
- Haig, D.; Westoby, M. Parent-Specific Gene Expression and the Triploid Endosperm. Am. Nat. 1989, 134, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Liu, C.; Qi, X.; Jiao, Y.; Wang, D.; Wang, Y.; Liu, Z.; Chen, C.; Chen, B.; Tian, X.; et al. Mutation of ZmDMP enhances haploid induction in maize. Nat. Plants 2019, 5, 575–580. [Google Scholar] [CrossRef]
- Kuppu, S.; Ron, M.; Marimuthu, M.P.; Li, G.; Huddleson, A.; Siddeek, M.H.; Terry, J.; Buchner, R.; Shabek, N.; Comai, L.; et al. A variety of changes, including CRISPR/Cas9-mediated deletions, in CENH3 lead to haploid induction on outcrossing. Plant Biotechnol. J. 2020, 18, 2068–2080. [Google Scholar] [CrossRef]
- Lv, J.; Yu, K.; Wei, J.; Gui, H.; Liu, C.; Liang, D.; Wang, Y.; Zhou, H.; Carlin, R.; Rich, R.; et al. Generation of paternal haploids in wheat by genome editing of the centromeric histone CENH3. Nat. Biotechnol. 2020, 38, 1397–1401. [Google Scholar] [CrossRef]
- Zhong, Y.; Chen, B.; Li, M.; Wang, D.; Jiao, Y.; Qi, X.; Wang, M.; Liu, Z.; Chen, C.; Wang, Y.; et al. A DMP-triggered in vivo maternal haploid induction system in the dicotyledonous Arabidopsis. Nat. Plants 2020, 6, 466–472. [Google Scholar] [CrossRef]
- Cheng, Z.; Sun, Y.; Yang, S.; Zhi, H.; Yin, T.; Ma, X.; Sui, Y. Establishing in planta haploid inducer line by edited SiMTL in foxtail millet (Setaria italica). Plant Biotechnol. J. 2021, 19, 1089–1091. [Google Scholar]
- Li, Y.; Lin, Z.; Yue, Y.; Zhao, H.; Fei, X.; Liu, C.; Chen, S.; Lai, J.; Song, W. Loss-of-function alleles of ZmPLD3 cause haploid induction in maize. Nat. Plants 2021, 7, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Chen, B.; Wang, D.; Zhu, X.; Li, M.; Zhang, J.; Chen, M.; Wang, M.; Riksen, T.; Liu, J.; et al. In vivo maternal haploid induction in tomato. Plant Biotechnol. J. 2021, 20, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Bohanec, B.; Jakše, M. Variations in gynogenic response among long-day onion (Allium cepa L.) accessions. Plant Cell Rep. 1999, 18, 737–742. [Google Scholar] [CrossRef]
- Dong, L.; Li, L.; Liu, C.; Liu, C.; Geng, S.; Li, X.; Xie, C. Genome editing and dou-ble-fluorescence proteins enable robust maternal haploid induction and identification in Maize. Mol. Plant. 2018, 11, 1214–1217. [Google Scholar] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ → 3′) | Product Size (bp) | Application |
|---|---|---|---|
| bar-F bar-R | ACCATCGTCAACCACTACATCG GCTGCCAGAAACCCACGTCATG | 430 | Detecting CRISPR/Cas9 construct |
| Cas9-F Cas9-R | AGGAGACTATCACCCCTTGGAAC TTGAAGGTAAGAGAGTCATCGTGG | 706 | Detecting CRISPR/Cas9 construct |
| Female Parents | Crossing Type or Male Parent | Seed Setting Rate (%) | Haploid Induction Rate (%) | Number of Normal Grains | Number of Embryoless Grains | Frequency for Embryoless Grains (%) |
|---|---|---|---|---|---|---|
| mtl-A | ⓧ a | 96.22 | 0 | 1289 | 0 | 0 |
| mtl-AD | ⓧ | 52.94 | 7.8 | 814 | 252 | 23.6 |
| mtl-BD | ⓧ | 39.47 | 15.6 | 934 | 261 | 21.8 |
| mtl-ABD | ⓧ | 32.41 | 13.9 | 1124 | 356 | 24.1 |
| Fielder | mtl-A | 89.97 | 0 | 529 | 1 | 0.2 |
| Fielder | mtl-AD | 28.06 | 9.1 | 142 | 47 | 24.9 |
| Fielder | mtl-BD | 27.82 | 13.0 | 158 | 55 | 25.8 |
| Fielder | mtl-ABD | 25.71 | 14.1 | 193 | 73 | 27.4 |
| Female Parents | Grain Length of Normal Grains (mm) | Grain Length of Embryoless Grains (mm) | Grain Width of Normal Grains (mm) | Grain Width of Embryoless Grains (mm) | TKW for Normal Grains (g) | TKW for Embryoless Grains (g) |
|---|---|---|---|---|---|---|
| Fielder | 6.9 ± 0.14 | --- | 3.5 ± 0.02 | --- | 42.53 | --- |
| mtl-A | 7.0 ± 0.06 | --- | 3.5 ± 0.05 | --- | 42.62 | --- |
| mtl-AD | 7.3 * ± 0.01 | 7.2 * ± 0.01 | 3.9 * ± 0.06 | 4.0 * ± 0.08 | 48.92 | 48.25 |
| mtl-BD | 7.3 * ± 0.01 | 7.2 * ± 0.03 | 4.0 * ± 0.01 | 3.9 * ± 0.04 | 47.32 | 47.07 |
| mtl-ABD | 7.3 * ± 0.04 | 7.2 * ± 0.06 | 3.9 * ± 0.02 | 3.8 * ± 0.01 | 47.27 | 46.53 |
| Days Post Anthesis (d) | Number of Normal Grains | Number of Embryoless Grains | Frequency for Embryoless Grains (%) |
|---|---|---|---|
| 9 | 287 | 112 | 28.1 |
| 12 | 221 | 95 | 30.1 |
| 15 | 230 | 103 | 30.9 |
| 18 | 229 | 91 | 28.4 |
| 23 | 234 | 84 | 26.4 |
| 28 | 243 | 93 | 27.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, H.; Zhang, S.; Yu, M.; Wang, K.; Yu, Y.; Qiu, Y.; Chang, Y.; Lin, Z.; Du, L.; Fu, D.; et al. Effects of TaMTL-Edited Mutations on Grain Phenotype and Storage Component Composition in Wheat. Agriculture 2022, 12, 587. https://doi.org/10.3390/agriculture12050587
Tang H, Zhang S, Yu M, Wang K, Yu Y, Qiu Y, Chang Y, Lin Z, Du L, Fu D, et al. Effects of TaMTL-Edited Mutations on Grain Phenotype and Storage Component Composition in Wheat. Agriculture. 2022; 12(5):587. https://doi.org/10.3390/agriculture12050587
Chicago/Turabian StyleTang, Huali, Shuangxi Zhang, Mei Yu, Ke Wang, Yang Yu, Yuliang Qiu, Yanan Chang, Zhishan Lin, Lipu Du, Daolin Fu, and et al. 2022. "Effects of TaMTL-Edited Mutations on Grain Phenotype and Storage Component Composition in Wheat" Agriculture 12, no. 5: 587. https://doi.org/10.3390/agriculture12050587
APA StyleTang, H., Zhang, S., Yu, M., Wang, K., Yu, Y., Qiu, Y., Chang, Y., Lin, Z., Du, L., Fu, D., Li, Z., & Ye, X. (2022). Effects of TaMTL-Edited Mutations on Grain Phenotype and Storage Component Composition in Wheat. Agriculture, 12(5), 587. https://doi.org/10.3390/agriculture12050587