
Cross-Resistance Pattern and Genetic Studies in Spirotetramat-Resistant Citrus Red Mite, Panonychus citri (Acari: Tetranychidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acaricides and Other Chemicals
2.2. Citrus Red Mite
2.3. Larval and Egg Bioassays
2.4. Cross-Resistance Studies
2.5. Crossing Experiment and Toxicological Test (Bioassay–Reciprocal Crossing LS-FJ♀ × ST-NK♂ and ST-NK♀ × LS-FJ♂)
2.5.1. Crossing Experiment
2.5.2. Toxicological Test
2.6. Data Analysis
3. Results
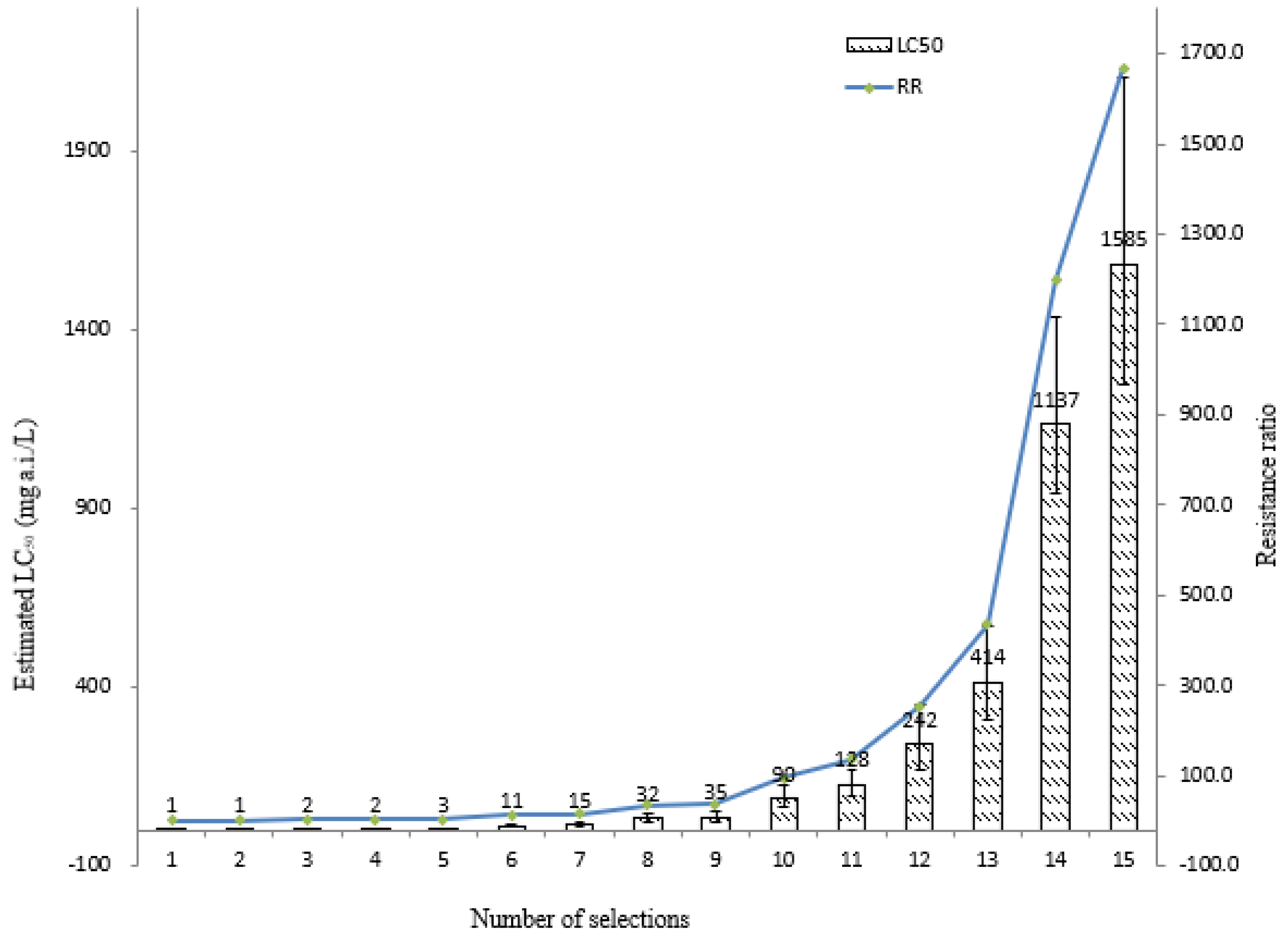
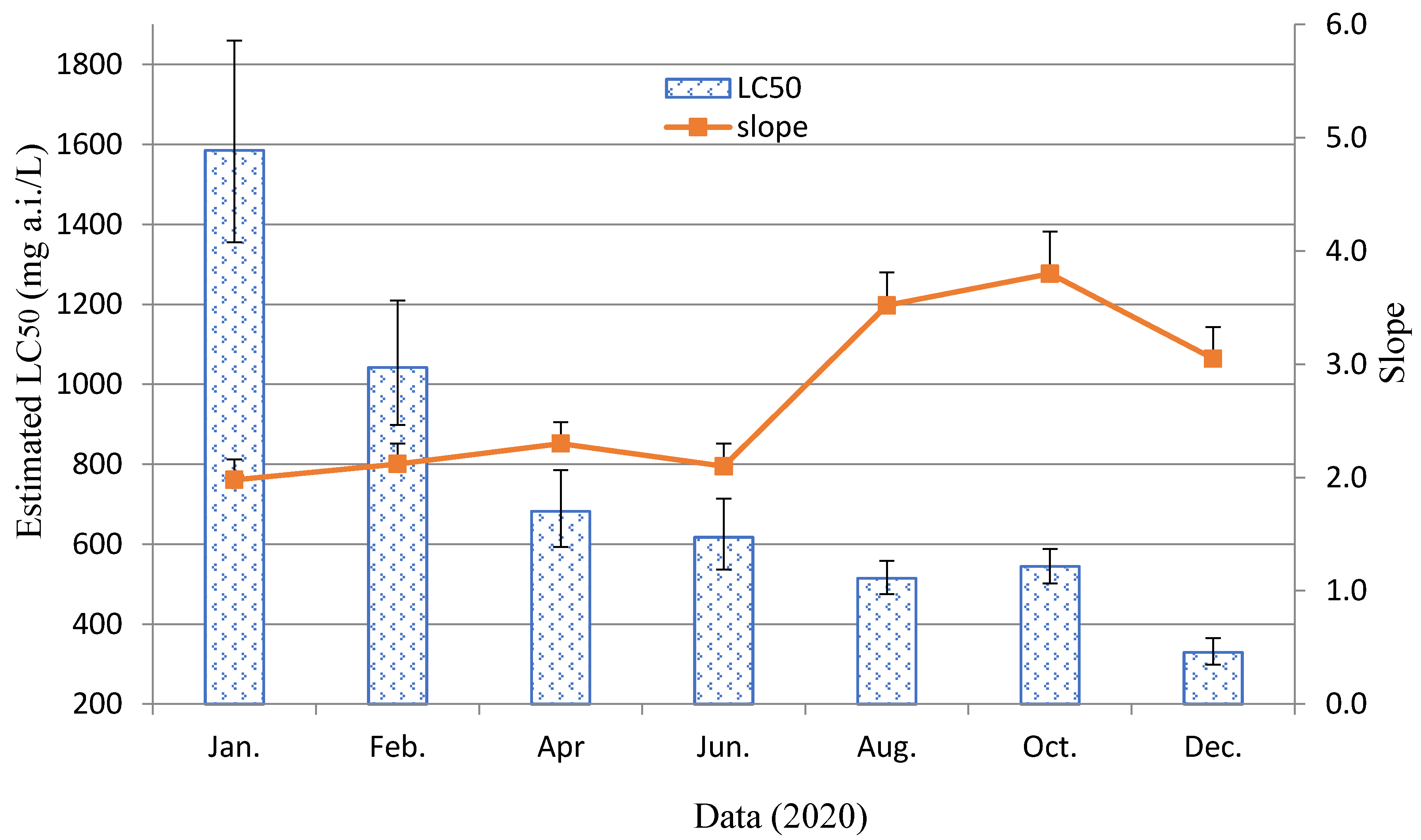
3.1. Susceptibilities of LS-FJ and Selected ST-NK Strain Stability
3.2. Cross-Resistance Studies
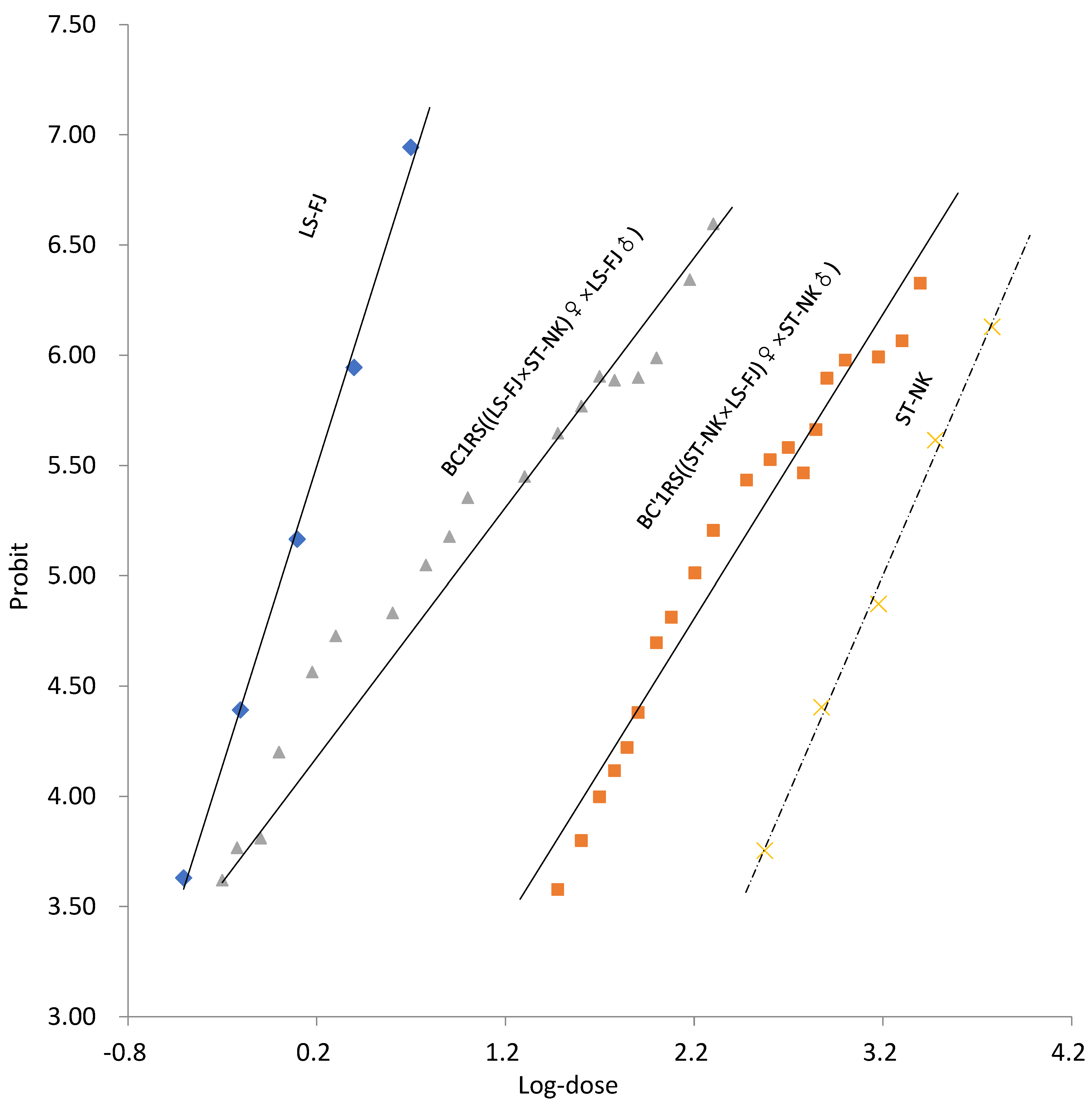
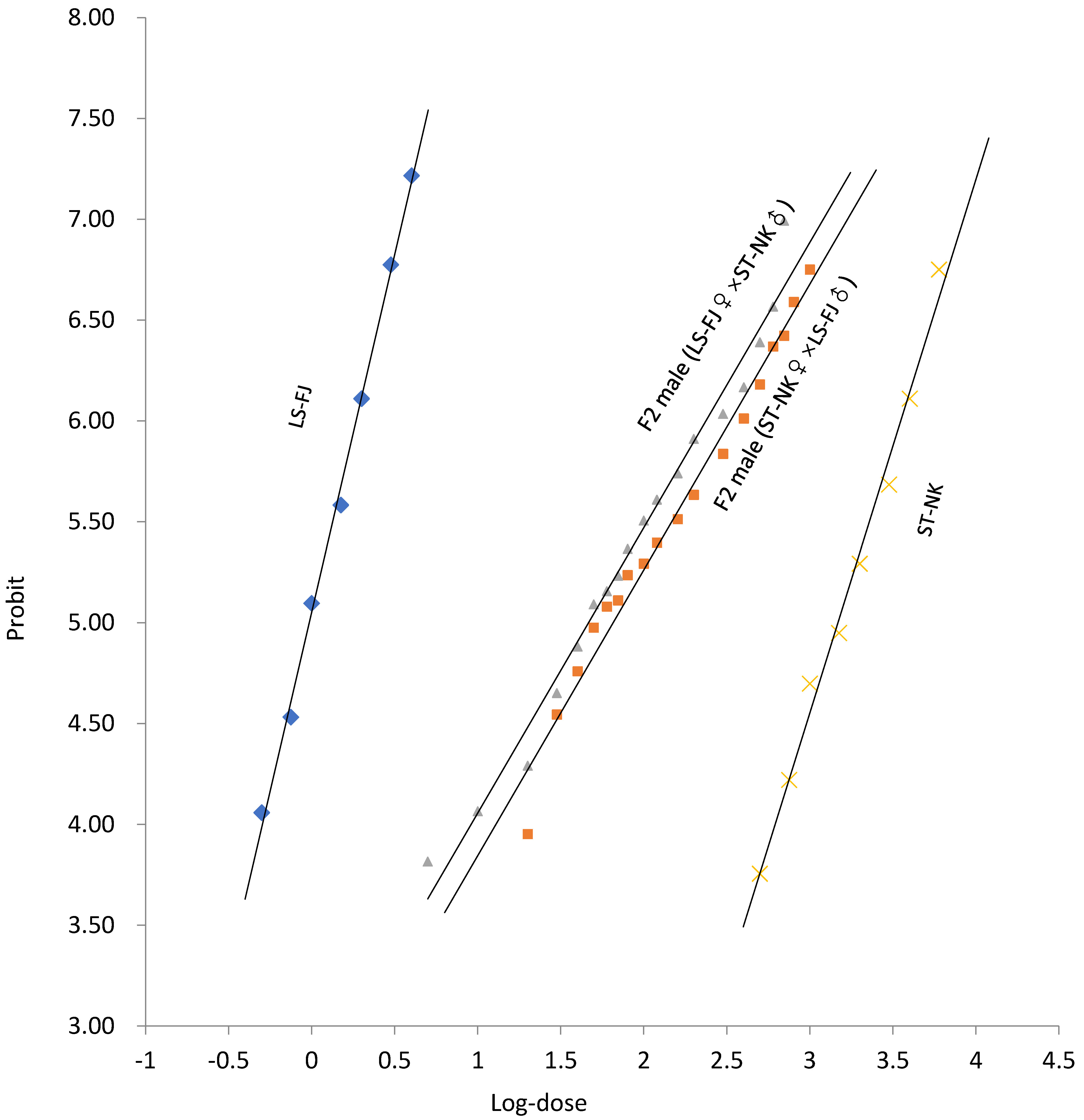
3.3. Mode of Inheritance of Spirotetramat Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China In China Agricultural Yearbook 2020; China Agricultural Publishing House: Beijing, China, 2021.
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef] [PubMed]
- ICAMA (Institute for the Control of Agrochemicals, Ministry of Agriculture). Pesticide Registration Public Announcement. 2021. Available online: http://www.chinapesticide.org.cn/hysj/index.jhtml. (accessed on 6 June 2021).
- Bretschneider, T.; Fischer, R.; Nauen, R. Inhibitors of Lipid Synthesis (Acetyl-CoA-carboxylase Inhibitors). In Modern Crop Protection Compounds; Kramer, W., Schirmer, U., Eds.; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2007; pp. 909–925. [Google Scholar]
- Brück, E.; Elbert, A.; Fischer, R.; Krueger, S.; Kühnhold, J.; Michael, K.A.; Nauen, R.; Niebes, J.F.; Reckmann, U.; Schnorbach, H.J.; et al. Movento®, an innovative ambimobile insecticide for sucking insect pest control in agriculture: Biological profile and field performance. Crop Prot. 2009, 10, 838–844. [Google Scholar] [CrossRef]
- Ouyang, Y.; Montez, G.H.; Liu, L.; Grafton-Cardwell, E.E. Spirodiclofen and spirotetramat bioassays for monitoring resistance in citrus redmite, Panonychus citri (Acari: Tetranychidae). Pest Manag. Sci. 2012, 68, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Bielza, P.; Moreno, I.; Belando, A.; Gravalos, C.; Izquierdo, J.; Nauen, R. Spiromesifen and spirotetramat resistance in field populations of Bemisia tabaci Gennadius in Spain. Pest Manag. Sci. 2019, 75, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.K.; Zheng, H.X.; Xie, W.; Wang, S.L.; Wu, Q.J.; Zhang, Y.J. Field resistance monitoring of the immature stages of the whitefly Bemisia tabaci to spirotetramat in China. Crop Prot. 2017, 98, 243–247. [Google Scholar] [CrossRef]
- Pan, Y.O.; Yang, C.; Gao, X.W.; Peng, T.F.; Bi, R.; Xi, J.H.; Xin, X.C.; Zhu, E.; Wu, Y.Q.; Shang, Q.L. Spirotetramat resistance adaption analysis of Aphis gossypii Glover by transcriptomic survey. Pestic. Biochem. Physiol. 2015, 124, 73–80. [Google Scholar] [CrossRef]
- Ejaz, M.; Shad, S.A. Spirotetramat Resistance Selected in the Phenacoccus solenopsis (Homoptera: Pseudococcidae): Cross-Resistance Patterns, Stability, and Fitness Costs Analysis. J. Econ. Entomol. 2017, 110, 1226–1234. [Google Scholar] [CrossRef]
- Zanardi, O.Z.; Bordini, G.P.; Franco, A.A.; de Morais, M.R.; Yamamoto, P.T. Development and reproduction of Panonychus citri (Prostigmata: Tetranychidae) on different species and varieties of citrus plants. Exp. Appl. Acarol. 2015, 67, 565–581. [Google Scholar] [CrossRef]
- Huang, M.D. Integrated Management of Citrus Red Mite; Chinese Science Press: Beijing, China, 1979; pp. 361–369. [Google Scholar]
- Chen, N.C.; Huang, H.Z.; Peng, Y.L. Monitoring of pesticide-resistance and preliminary studies on main resistance mechanism of citric spider mite (Panonychus citri McGregor). Pesticides 1996, 35, 15–18. [Google Scholar]
- Chen, G.H.; Wang, A.Z. Test on the resistance of Panonychus citri to Dicofol and pyridaben. Hunan Agri. Sci. 2001, 4, 48. [Google Scholar]
- Hu, J.; Wang, C.F.; Wang, J.; You, Y.; Chen, F. Monitoring of resistance to spirodiclofen and five other acaricides in Panonychus citri collected from Chinese citrus orchards. Pest Manag. Sci. 2010, 66, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Döker, İ.; Kazak, C.; Ay, R. Resistance status and detoxification enzyme activity in ten populations of Panonychus citri (Acari: Tetranychidae) from Turkey. Crop Prot. 2021, 141, 105488. [Google Scholar] [CrossRef]
- Shelton, A.M.; Sances, F.V.; Hawley, J.; Tang, D.J.; Boune, M.; Jungers, D.; Collins, H.L.; Farias, J. Assessment of insecticide resistance after the outbreak of diamondback moth (Lepidoptera: Plutellidae) in California in 1997. J. Econ. Entomol. 2000, 93, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.Z.; Li, Y.X.; Collins, H.L.; Gusukuma-Minuto, L.; Mau, R.F.L.; Thompson, G.D.; Shelton, A.M. Monitoring and characterization of diamondback moth resistance to spinosad. J. Econ. Entomol. 2002, 95, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, A.; Yoneda, H.; Hatano, R.; Asada, M. Genetic analysis of hexythiazox resistance in the citrus red mite, Panonychus citri (McGregor). J. Pestic. Sci. 1995, 20, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Van Pottelberge, S.; Van Leeuwen, T.; Jahangir, K.; Luc, T. Genetic and biochemical analysis of a laboratory-selected spirodiclofen-resistant strain of Tetranychus urticae Koch (Acari: Tetranychidae). Pest Manag. Sci. 2009, 65, 358–366. [Google Scholar] [CrossRef]
- Potter, C. An improved apparatus for applying direct sprays and surface films with data on the electrostatic charge on atomized spray fluids. Ann. Appl. Biol. 1952, 39, 1–28. [Google Scholar] [CrossRef]
- Sabine, G.; Philippe, A.; Serge, K. The acaricidal effect of sulfur on Tetranychus urticae (Acari: Tetranychidae) under laboratory conditions. 11. International Congress of Acarology, Sep 2002, Mexico, Mexico. Universidad nacional autonoma de Mexico, 2007. Available online: https://hal.archives-ouvertes.fr/hal-01231316/document. (accessed on 3 April 2021).
- Stone, B.F. A formula for determining degree of dominance in cases of mono factorial inheritance of resistance to chemicals. Bull. WHO 1968, 38, 325–326. [Google Scholar]
- Georghiou, G.P. Genetics of resistance to insecticides in house flies and mosquitoes. Exp. Parasitol. 1969, 26, 224–255. [Google Scholar] [CrossRef]
- Abbott, S.W. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Fukami, J.; Uesugi, Y.; Ishizuka, K. Pest Resistance to Pesticides; Soft Science Inc.: Tokyo, Japan, 1983. [Google Scholar]
- Yu, D.Y.; Wang, C.F.; Yu, Y.; Huang, Y.Q.; Yao, J.A.; Hu, J.F. Laboratory selection for spirodiclofen resistance and cross-resistance in Panonychus citri. Afr. J. Biotechnol. 2011, 17, 24–34. [Google Scholar]
- Brown, T.M.; Payne, G.T. Experimental Selection for Insecticide Resistance. J. Econ. Entomol. 1988, 1, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Mário, E.S.; Bruce, V.; Rafael, S.M.S.; Maria, C.V.Q.; Rafaela, G. Spiromesifen resistance in Tetranychus urticae (Acari: Tetranychidae): Selection, stability, and monitoring. Crop Prot. 2016, 89, 278–283. [Google Scholar]
- Yamamoto, A.; Yoneda, H.; Hatano, R.; Asada, M. Stability of hexythiazox resistance in the citrus red mite, Panonychus citri (McGregor) under laboratory and field conditions. J. Pestic. Sci. 1996, 20, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Demaeght, P.; Dermauw, W.; Tsakireli, D.; Khajehali, J.; Nauen, R.; Tirry, L.; Vontas, J.; Lümmen, P.; Van Leeuwen, T. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae: CYP392E10 metabolizes spirodiclofen, but not its corresponding enol. Insect Biochem. Mol. Biol. 2013, 43, 544–554. [Google Scholar] [CrossRef]
- Kramer, T.; Nauen, R. Monitoring of spirodiclofen susceptibility in field populations of European red mites, Panonychus ulmi (Koch) (Acari: Tetranychidae), and the cross-resistance pattern of a laboratory-selected strain. Pest Manag. Sci. 2011, 67, 1285–1293. [Google Scholar] [CrossRef]
- Dekeyser, M.A. Acaricide mode of action. Pest Manag. Sci. 2005, 61, 103–110. [Google Scholar] [CrossRef]
- Elbert, A.; Nauen, R.; Salmon, E. Resistance management guidelines for the new ketoenol insecticide Movento®. Bayer Crop Sci. J. 2008, 2, 403–416. [Google Scholar]
- Sayyed, A.H.; Wright, D.J. Fipronil resistance in the diamondback moth (Lepidoptera: Plutellidae): Inheritance and number of genes involved. J. Econ. Entomol. 2004, 97, 2043–2050. [Google Scholar] [CrossRef]
- Uesugi, R.; Goka, K.; Osakabe, M.H. Genetic basis of resistance to chlorfenapyr and etoxazole in the two-spotted spider mite (Acari: Tetranychidae). J. Econ. Entomol. 2002, 95, 1267–1274. [Google Scholar] [CrossRef]
- Demaeght, P.; Osborne, E.J.; J Odman-Naresh, O.; Grbić, M.; Nauen, R.; Merzendorfer, H.; Clark, R.M.; Van Leeuwen, T. High resolution genetic mapping uncovers chitin synthase-1 as the target-site of the structurally diverse mite growth inhibitors clofentezine, hexythiazox and etoxazole in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 51, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, B.A.; Van De Baan, H.E. Ecological and genetic factors influencing evolution of pesticide resistance in tetranychid and phytoseiid mites. Exp. Appl. Acarol. 1988, 4, 277–300. [Google Scholar] [CrossRef]
- Roush, R.T.; McKenzie, J.A. Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol. 1987, 32, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Helle, W.; Pijnacker, L.P. Parthenogenesis, chromosomes and sex. In Spider Mites. Their Biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1A, pp. 129–139. [Google Scholar]
- Crozier, R.H. Adaptive consequences of male-haploidy. In Spider Mites. Their Biology, Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; Volume 1A, pp. 201–222. [Google Scholar]
- Van Leeuwen, T.; Stillatus, V.; Tirry, L. Genetic analysis and cross-resistance spectrum of a laboratory-selected chlorfenapyr resistant strain of two-spotted spider mite (Acari: Tetranychidae). Exp. Appl. Acarol. 2004, 32, 249–261. [Google Scholar] [CrossRef]
- Herron, G.A.; Rophail, J. Genetics of hexythiazox resistance in two spotted spider mite, Tetranychus urticae Koch. Exp. Appl. Acarol. 1993, 17, 423–431. [Google Scholar] [CrossRef]
- Asahara, M.; Uesugi, R.; Osakabe, M.H. Linkage between one of the polygenic hexythiazox resistance genes and an etoxazole resistance gene in the two spotted spider mite (Acari: Tetranychidae). J. Econ. Entomol. 2008, 101, 1704–1710. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
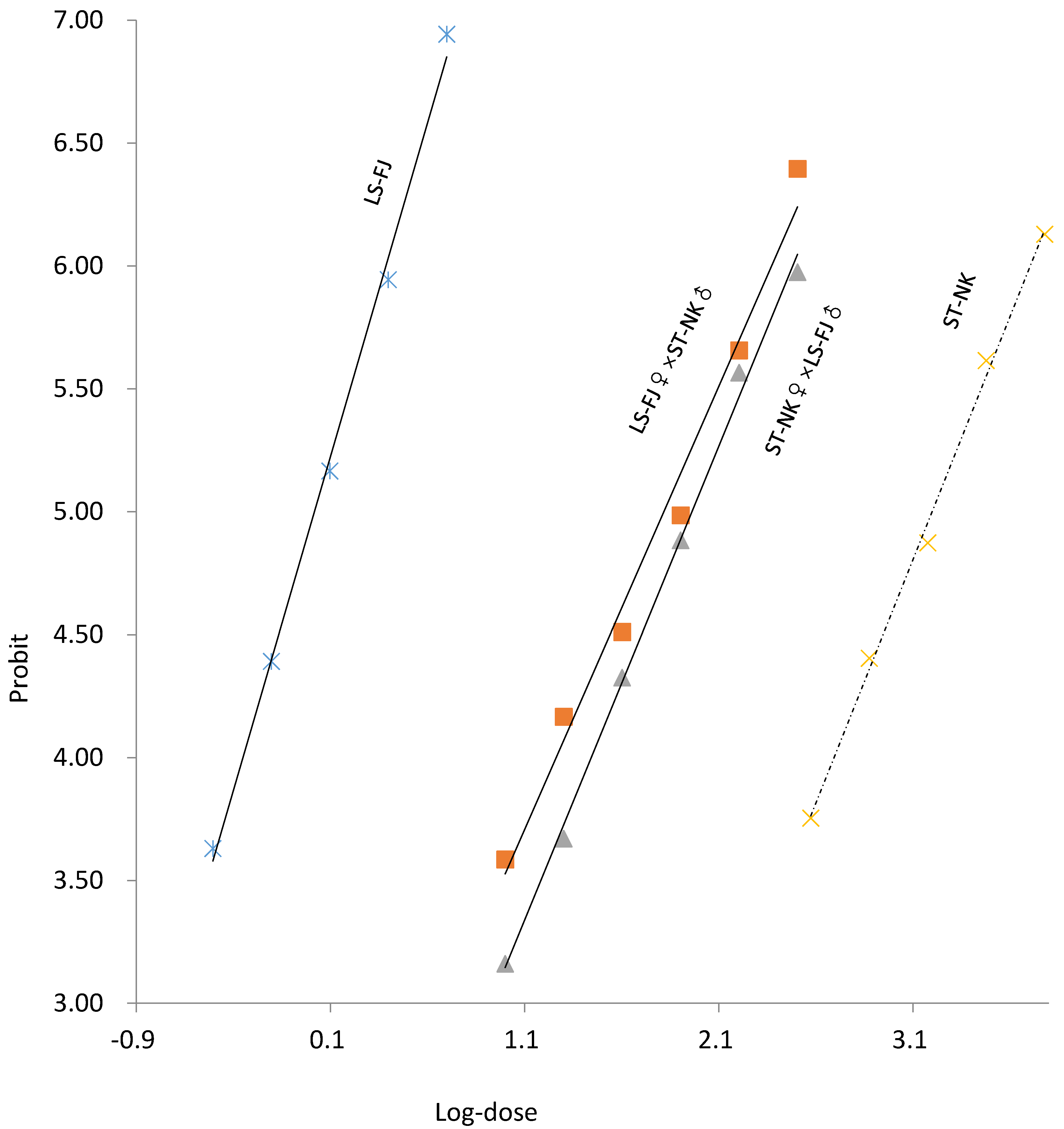{kind=link}
{kind=link}
{kind=link}
| LS-FJ | ST-NK | |||||
|---|---|---|---|---|---|---|
| Acaricides | Slope (±SE) | LC50 (95%CI) (mg L−1) | Slope (±SE) | LC50 (95%CI) (mg L−1) | RR | |
| Spirotetramat | Eggs | 3.41 (±0.25) | 74.15 (71.10–77.33) | 1.9 (0.22) | 10057.19 (8637.30–11,977.19) | 135.6 |
| Larvae | 5.25 (0.43) | 0.95 (0.89–1.01) | 1.98 (0.18) | 1585.01 (1355.94–1860.78) | 1668.4 | |
| Spirodiclofen | Eggs | 3.51 (0.28) | 4.57 (4.38–4.76) | 2.03 (0.18) | 163.02 (139.78–189.82) | 35.7 |
| Larvae | 4.42 (±0.32) | 1.2 (1.12–1.29) | 2.15 (±0.19) | 90.04 (77.43–104.14) | 75.0 | |
| Spiromesifen | Eggs | 2.91 (±0.23) | 2.69 (2.54–2.83) | 2.06 (±0.18) | 43.33 (37.22–50.36) | 16.1 |
| Larvae | 4.1 (±0.31) | 0.9 (0.84–0.97) | 2.68 (±0.21) | 28.55 (24.36–33.51) | 31.7 | |
| Etoxazole | Eggs | 2.56 (±0.19) | 1.38 (1.30–1.46) | 1.98 (±0.19) | 1.49 (1.26–1.76) | 1.1 |
| Larvae | 3.71 (±0.29) | 1.27 (1.17–1.38) | 2.06 (±0.16) | 1.28 (1.10–1.48) | 1.0 | |
| Abamectin | Eggs | 3.94 (±0.31) | 3.04 (2.92–3.16) | 0.92 (±0.08) | 4.24 (3.67–4.91) | 1.4 |
| Larvae | 3.72 (±0.34) | 1.31 (1.20–1.42) | 2.07 (±0.18) | 1.46 (1.25–1.70) | 1.1 | |
| Pyridaben | Eggs | 2.15 (±0.19) | 4.54 (3.89–5.27) | 1.83 (±0.17) | 8.4 (7.06–9.93) | 1.9 |
| Larvae | 2.08 (±0.18) | 5.51 (4.76–6.40) | 2.27 (±0.19) | 10.03 (8.71–11.56) | 1.8 | |
| Hexythiazox | Eggs | 2.5 (±0.2) | 9.46 (8.90–10.05) | 1.04 (±0.09) | 23.01 (20.12–26.33) | 2.4 |
| Larvae | 2.03 (±0.18) | 17.72 (15.21–20.56) | 2.1 (±0.18) | 102.33 (88.18–118.80) | 5.8 | |
| Clofentezine | Eggs | 2.41 (±0.18) | 25.13 (23.66–26.61) | 1.09 (±0.09) | 50.3 (44.11–57.60) | 2.0 |
| Larvae | 1.65 (±0.13) | 19.32 (16.28–23.02) | 2.04 (±0.18) | 95.67 (82.13–111.18) | 5.0 | |
| Strain | n | Slope (±SE) | LC50 (95%CI) (mg L−1) | LC90 (95%CI) (mg L−1) | D | |
|---|---|---|---|---|---|---|
| Diploid F1 progeny | LS-FJ♀ × ST-NK♂ | 2355 | 1.83 (±0.17) | 68.81 (58.18–81.05) | 345.79 (260.70–509.88) | 0.15 |
| ST-NK♀ × LS-FJ♂ | 2445 | 1.98 (±0.17) | 91.15 (78.22–106.85) | 416.67 (316.38–604.73) | 0.23 | |
| Haploid F2 progeny | LS-FJ♀ × ST-NK♂ | 3882 | 1.24 (±0.05) | 49.91 (45.00–55.06) | 544.85 (461.98–657.54) | 0.07 |
| ST-NK♀ × LS-FJ♂ | 3759 | 1.38 (±0.06) | 63.06 (56.83–69.36) | 527.91 (458.06–620.86) | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Wang, J.; Yu, Y.; Rao, W.; Chen, F.; Wang, C.; Fan, G. Cross-Resistance Pattern and Genetic Studies in Spirotetramat-Resistant Citrus Red Mite, Panonychus citri (Acari: Tetranychidae). Agriculture 2022, 12, 737. https://doi.org/10.3390/agriculture12050737
Hu J, Wang J, Yu Y, Rao W, Chen F, Wang C, Fan G. Cross-Resistance Pattern and Genetic Studies in Spirotetramat-Resistant Citrus Red Mite, Panonychus citri (Acari: Tetranychidae). Agriculture. 2022; 12(5):737. https://doi.org/10.3390/agriculture12050737
Chicago/Turabian StyleHu, Jinfeng, Jun Wang, Yun Yu, Wenhua Rao, Feng Chen, Changfang Wang, and Guocheng Fan. 2022. "Cross-Resistance Pattern and Genetic Studies in Spirotetramat-Resistant Citrus Red Mite, Panonychus citri (Acari: Tetranychidae)" Agriculture 12, no. 5: 737. https://doi.org/10.3390/agriculture12050737
APA StyleHu, J., Wang, J., Yu, Y., Rao, W., Chen, F., Wang, C., & Fan, G. (2022). Cross-Resistance Pattern and Genetic Studies in Spirotetramat-Resistant Citrus Red Mite, Panonychus citri (Acari: Tetranychidae). Agriculture, 12(5), 737. https://doi.org/10.3390/agriculture12050737