Phenotypic Analysis and Gene Cloning of a New Allelic Mutant of SPL5 in Rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Investigation of Chlorophyll Content
2.3. Determination of Various Antioxidant Indexes
2.4. Real-Time PCR Analysis
2.5. Histochemical Label Staining
2.6. Gene Mapping
2.7. Subcellular Localization
3. Results
3.1. Phenotypic Characteristics of Mutant lm5
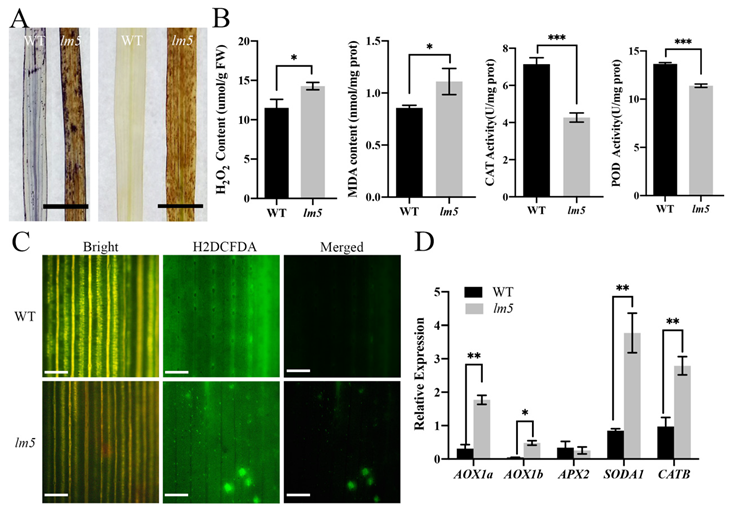
3.2. ROS Accumulation Occurred in lm5 Mutants
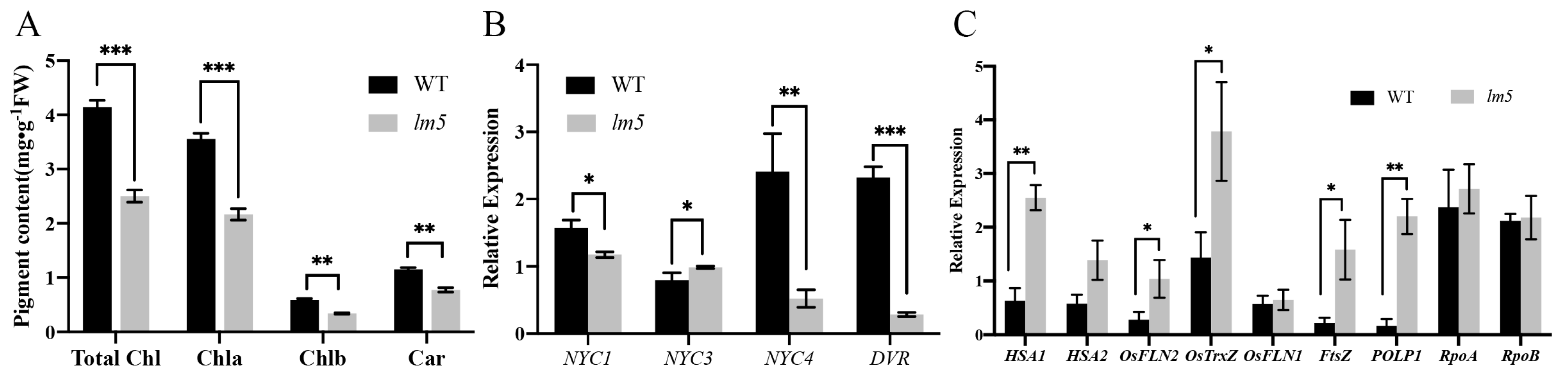
3.3. The Chlorophyll Content of the lm5 Mutant Decreased
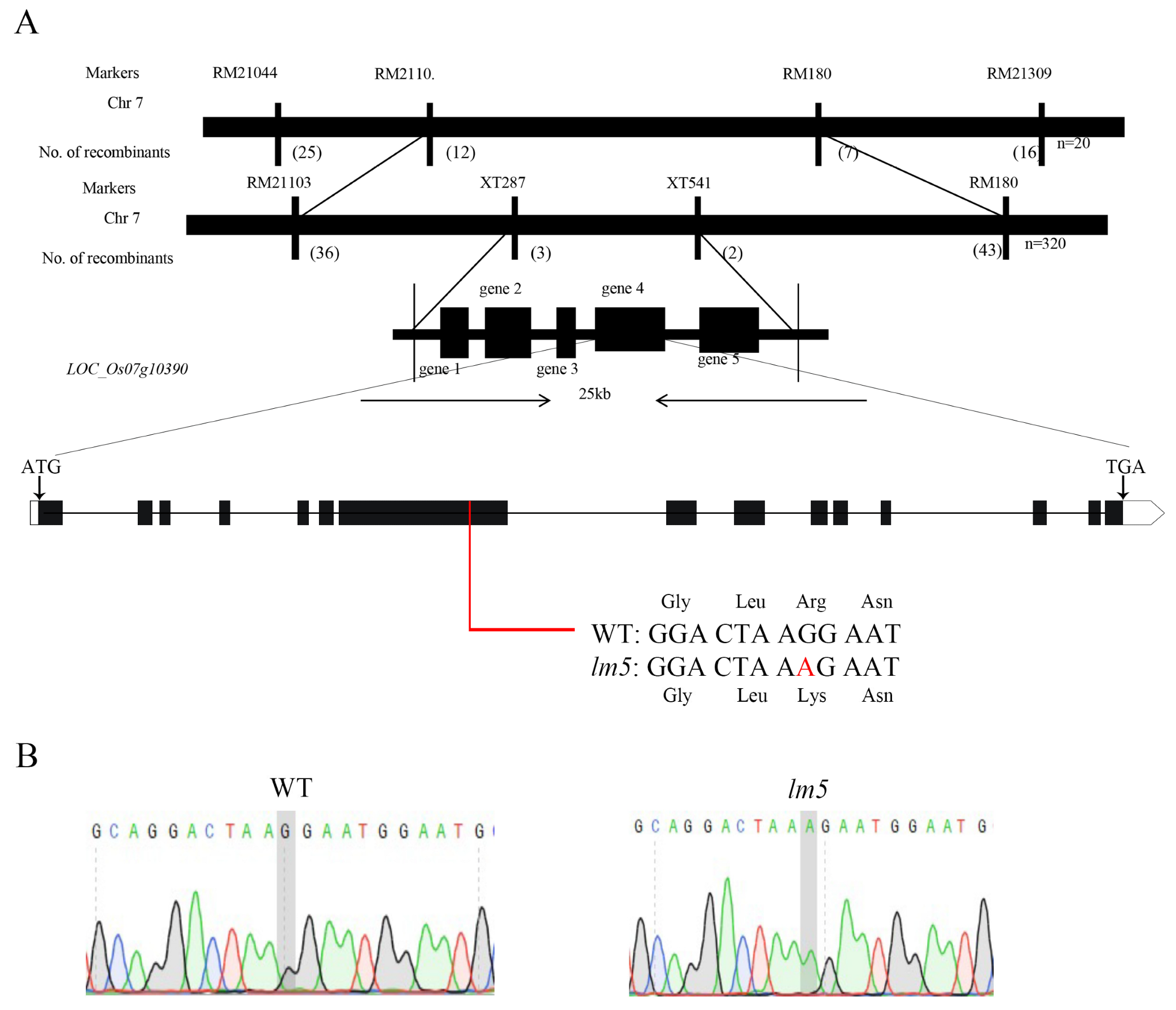
3.4. Genetic Analysis and Candidate Gene Mapping of Macular Mutant lm5
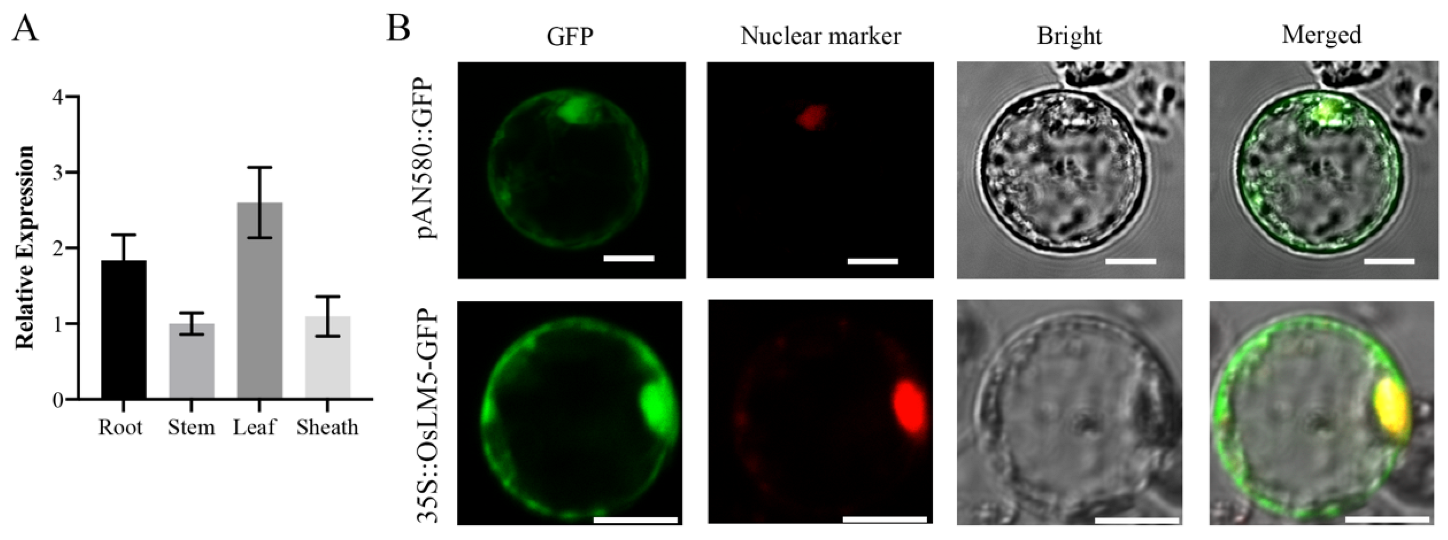
3.5. OsLM5 Expression Pattern Analysis
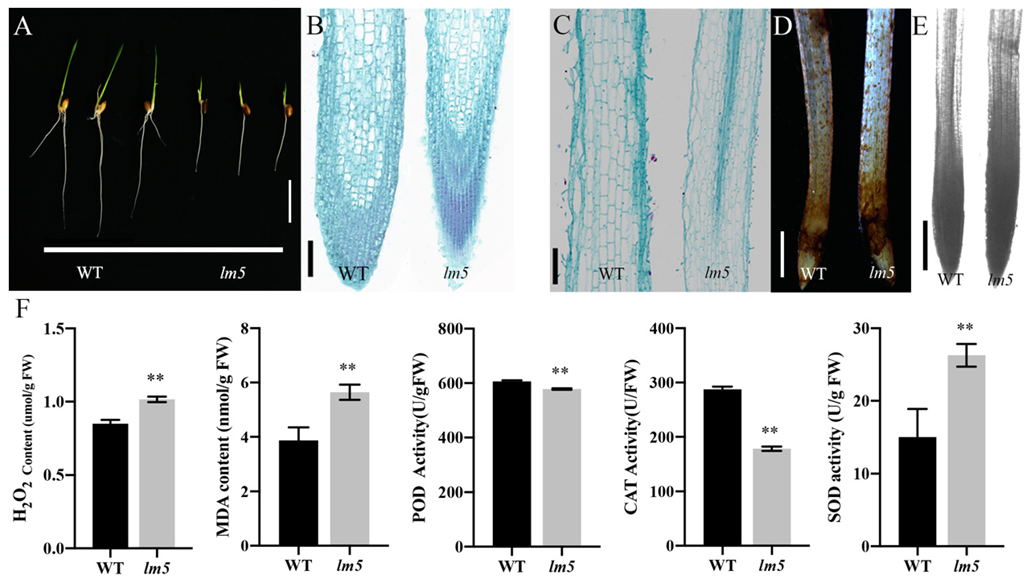
3.6. OsLM5 Mutations Affect Root Development
4. Discussion
4.1. The Formation of Leaf Lesions Affected the Agronomic Traits of Mutant lm5
4.2. ROS Accumulation Led to the Formation of Disease-like Spots in Mutant lm5
4.3. Effects of Mutant lm5 on Photosynthetic Function in Leaves
4.4. OsLM5 Is a New Allele of SPL5
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gross, B.L.; Zhao, Z. Archaeological and genetic insights into the origins of domesticated rice. Proc. Natl. Acad. Sci. USA 2014, 111, 6190–6197. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Peng, Y.; Zhang, Q.; Xia, S.; Ruan, B.; Xu, Q.; Yu, X.; Zhou, T.; Liu, H.; Zeng, D.; et al. Disruption of EARLY LESION LEAF 1, encoding a cytochrome P450 monooxygenase, induces ROS accumulation and cell death in rice. Plant J. 2020, 105, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Richter, T.E.; Hulbert, S.H.; Pryor, T. Disease Lesion Mimicry Caused by Mutations in the Rust Resistance Gene rp1. Plant Cell 1996, 8, 1367–1376. [Google Scholar] [CrossRef]
- Wu, C.; Bordeos, A.; Madamba, M.R.S.; Baraoidan, M.; Ramos, M.; Wang, G.-L.; Leach, J.E.; Leung, H. Rice lesion mimic mutants with enhanced resistance to diseases. Mol. Genet. Genom. 2008, 279, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Yan, M.; Yun, H.; Tan, J.; Du, D.; Sun, H.; Guo, Y.; Sang, X.; Zhang, C. Identification and fine mapping of lesion mimic mutant spl36 in rice (Oryza sativa L.). Breed. Sci. 2021, 71, 510–519. [Google Scholar] [CrossRef]
- Kelly, D.; Vatsa, A.; Mayham, W.; Kazic, T. Extracting complex lesion phenotypes in Zea mays. Mach. Vis. Appl. 2015, 27, 145–156. [Google Scholar] [CrossRef]
- Shirsekar, G.S.; Vega-Sanchez, M.E.; Bordeos, A.; Baraoidan, M.; Swisshelm, A.; Fan, J.; Park, C.H.; Leung, H.; Wang, G.-L. Identification and characterization of suppressor mutants of spl11-mediated cell death in rice. Mol. Plant-Microbe Interact. 2014, 27, 528–536. [Google Scholar] [CrossRef]
- Hoisington, D.; Neuffer, M.; Walbot, V. Disease lesion mimics in maize: I. Effect of genetic background, temperature, developmental age, and wounding on necrotic spot formation with Les1. Dev. Biol. 1982, 93, 381–388. [Google Scholar] [CrossRef]
- Wolter, M.; Hollricher, K.; Salamini, F.; Schulze-Lefert, P. The mlo resistance alleles to powdery mildew infection in barley trigger a developmentally controlled defence mimic phenotype. Mol. Genet. Genom. 1993, 239, 122–128. [Google Scholar] [CrossRef]
- Yao, Q.; Zhou, R.; Fu, T.; Wu, W.; Zhu, Z.; Li, A.; Jia, J. Characterization and mapping of complementary lesion-mimic genes lm1 and lm2 in common wheat. Theor. Appl. Genet. 2009, 119, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Spassieva, S.; Hille, J. A lesion mimic phenotype in tomato obtained by isolating and silencing an Lls1 homologue. Plant Sci. 2002, 162, 543–549. [Google Scholar] [CrossRef]
- Wang, S.-H.; Lim, J.-H.; Kim, S.-S.; Cho, S.-H.; Yoo, S.-C.; Koh, H.-J.; Sakuraba, Y.; Paek, N.-C. Mutation of SPOTTED LEAF3 (SPL3) impairs abscisic acid-responsive signalling and delays leaf senescence in rice. J. Exp. Bot. 2015, 66, 7045–7059. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef]
- Nurnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef]
- Yuchun, R.A.O.; Ran, J.I.A.O.; Sheng, W.A.N.G.; Xianmei, W.U.; Hanfei, Y.E.; Chenyang, P.A.N.; Sanfeng, L.I.; Dedong, X.; Weiyong, Z.H.O.U.; Gaoxing, D.A.I.; et al. SPL36 Encodes a Receptor-like Protein Kinase that Regulates Programmed Cell Death and Defense Responses in Rice. Rice 2021, 14, 34. [Google Scholar] [CrossRef]
- Sun, H.; Mao, J.; Lan, B.; Zhang, C.; Zhao, C.; Pan, G.; Pan, X. Characterization and mapping of a spotted-leaf genotype, spl (Y181) that confers blast susceptibility in rice. Eur. J. Plant Pathol. 2014, 140, 407–417. [Google Scholar] [CrossRef]
- Yin, Z.; Chen, J.; Zeng, L.; Goh, M.; Leung, H.; Khush, G.S.; Wang, G.-L. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol. Plant-Microbe Interact. 2000, 13, 869–876. [Google Scholar] [CrossRef]
- Shang, H.; Li, P.; Zhang, X.; Xu, X.; Gong, J.; Yang, S.; He, Y.; Wu, J.-L. The Gain-of-Function Mutation, OsSpl26, Positively Regulates Plant Immunity in Rice. Int. J. Mol. Sci. 2022, 23, 14168. [Google Scholar] [CrossRef]
- Wang, L.; Han, S.; Zhong, S.; Wei, H.; Zhang, Y.; Zhao, Y.; Liu, B. Characterization and fine mapping of a necrotic leaf mutant in maize (Zea mays L.). J. Genet. Genom. 2013, 40, 307–314. [Google Scholar] [CrossRef]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Tomita, C.; Sugimoto, K.; Hasegawa, M.; Hayashi, N.; Dubouzet, J.G.; Ochiai, H.; Sekimoto, H.; Hirochika, H.; Kikuchi, S. Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol. Biol. 2007, 63, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, Z.; Liu, T.; Gao, B.; Xiong, X. Identification and Map-Based Cloning of the Light-Induced Lesion Mimic Mutant 1 (LIL1) Gene in Rice. Front. Plant Sci. 2017, 8, 2122. [Google Scholar] [CrossRef] [PubMed]
- Kojo, K.; Yaeno, T.; Kusumi, K.; Matsumura, H.; Fujisawa, S.; Terauchi, R.; Iba, K. Regulatory mechanisms of ROI generation are affected by rice spl mutations. Plant Cell Physiol. 2006, 47, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-L.; Sun, A.-Z.; Chen, S.-T.; Chen, L.-S.; Guo, F.-Q. SPL6 represses signalling outputs of ER stress in control of panicle cell death in rice. Nat. Plants 2018, 4, 280–288. [Google Scholar] [CrossRef]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The Roles of Mitochondrial Reactive Oxygen Species in Cellular Signaling and Stress Response in Plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef]
- Dickman, M.B.; Fluhr, R. Centrality of host cell death in plant-microbe interactions. Annu. Rev. Phytopathol. 2013, 51, 543–570. [Google Scholar] [CrossRef]
- Wang, L.; Pei, Z.; Tian, Y.; He, C. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol. Plant-Microbe Interact. 2005, 18, 375–384. [Google Scholar] [CrossRef]
- Zeng, L.-R.; Qu, S.; Bordeos, A.; Yang, C.; Baraoidan, M.; Yan, H.; Xie, Q.; Nahm, B.H.; Leung, H.; Wang, G.-L. Spotted leaf11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell 2004, 16, 2795–2808. [Google Scholar] [CrossRef]
- Hu, P.; Tan, Y.; Wen, Y.; Fang, Y.; Wang, Y.; Wu, H.; Wang, J.; Wu, K.; Chai, B.; Zhu, L.; et al. LMPA Regulates Lesion Mimic Leaf and Panicle Development Through ROS-Induced PCD in Rice. Front. Plant Sci. 2022, 13, 875038. [Google Scholar] [CrossRef]
- Pei, Z.-M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Photosensitized oxidation of membrane lipids: Reaction pathways, cytotoxic effects, and cytoprotective mechanisms. J. Photochem. Photobiol. B Biol. 2001, 63, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Rebeiz, C.A.; Montazer-Zouhoor, A.; Mayasich, J.M.; Tripathy, B.C.; Wu, S.; Rebeiz, C.C.; Friedmann, H.C. Photodynamic herbicides. Recent developments and molecular basis of selectivity. Crit. Rev. Plant Sci. 1988, 6, 385–436. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yu, Y.; Chen, Q.; Mu, G.; Shen, Z.; Zheng, L. OsMYB45 plays an important role in rice resistance to cadmium stress. Plant Sci. 2017, 264, 1–8. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Chai, T.; Zhou, J.; Liu, J.; Xing, D. LSD1 and HY5 antagonistically regulate red light induced-programmed cell death in Arabidopsis. Front. Plant Sci. 2015, 6, 292. [Google Scholar] [CrossRef]
- Abou-Attia, M.A.; Wang, X.; Al-Attala, M.N.; Xu, Q.; Zhan, G.; Kang, Z. TaMDAR6 acts as a negative regulator of plant cell death and participates indirectly in stomatal regulation during the wheat stripe rust-fungus interaction. Physiol. Plant. 2015, 156, 262–277. [Google Scholar] [CrossRef]
- Xiao, G.; Zhou, J.; Lu, X.; Huang, R.; Zhang, H. Excessive UDPG resulting from the mutation of UAP1 causes programmed cell death by triggering reactive oxygen species accumulation and caspase-like activity in rice. New Phytol. 2017, 217, 332–343. [Google Scholar] [CrossRef]
- Yang, C.; Li, W.; Cao, J.; Meng, F.; Yu, Y.; Huang, J.; Jiang, L.; Liu, M.; Zhang, Z.; Chen, X.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2017, 89, 338–353. [Google Scholar] [CrossRef]
- Wang, S.; Lei, C.; Wang, J.; Ma, J.; Tang, S.; Wang, C.; Zhao, K.; Tian, P.; Zhang, H.; Qi, C.; et al. SPL33, encoding an eEF1A-like protein, negatively regulates cell death and defense responses in rice. J. Exp. Bot. 2017, 68, 899–913. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef]
- Silva-Navas, J.; Moreno-Risueno, M.A.; Manzano, C.; Téllez-Robledo, B.; Navarro-Neila, S.; Carrasco, V.; Pollmann, S.; Gallego, F.J.; del Pozo, J.C. Flavonols Mediate Root Phototropism and Growth through Regulation of Proliferation-to-Differentiation Transition. Plant Cell 2016, 28, 1372–1387. [Google Scholar] [CrossRef] [PubMed]
- Manzano, C.; Pallero-Baena, M.; Casimiro, I.; De Rybel, B.; Orman-Ligeza, B.; Van Isterdael, G.; Beeckman, T.; Draye, X.; Casero, P.; del Pozo, J.C. The Emerging Role of Reactive Oxygen Species Signaling during Lateral Root Development. Plant Physiol. 2014, 165, 1105–1119. [Google Scholar] [CrossRef] [PubMed]
- Clore, A.M.; Doore, S.M.; Tinnirello, S.M.N. Increased levels of reactive oxygen species and expression of a cytoplasmic aconitase/iron regulatory protein 1 homolog during the early response of maize pulvini to gravistimulation. Plant Cell Environ. 2007, 31, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.H.; Bae, Y.S.; Lee, J.S. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 2001, 126, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, L.; Pan, J.; Zheng, X.; Jiang, G.; Jin, Y.; Gu, Z.; Qian, Q.; Zhai, W.; Ma, B. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol. Breed. 2011, 30, 939–949. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Han, S.-H.; Sakuraba, Y.; Koh, H.-J.; Paek, N.-C. Leaf variegation in the rice zebra2 mutant is caused by photoperiodic accumulation of tetra-Cis-lycopene and singlet oxygen. Mol. Cells 2011, 33, 87–97. [Google Scholar] [CrossRef]
- Leshem, Y.; Melamed-Book, N.; Cagnac, O.; Ronen, G.; Nishri, Y.; Solomon, M.; Cohen, G.; Levine, A. Suppression of Arabidopsis vesicle-SNARE expression inhibited fusion of H2O2-containing vesicles with tonoplast and increased salt tolerance. Proc. Natl. Acad. Sci. USA 2006, 103, 18008–18013. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Liu, L.; Liu, Q.; Bi, Z.; Yu, N.; Cheng, S.; Cao, L. Fine mapping of the lesion mimic and early senescence 1 (lmes1) in rice (Oryza sativa). Plant Physiol. Biochem. 2014, 80, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Hess, W.R.; Börner, T. Organellar RNA polymerases of higher plants. Int. Rev. Cytol. 1999, 190, 1–59. [Google Scholar] [CrossRef] [PubMed]
- Shiina, T.; Tsunoyama, Y.; Nakahira, Y.; Khan, M.S. Plastid RNA polymerases, promoters, and transcription regulators in higher plants. Int. Rev. Cytol. 2005, 244, 1–68. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, S.; Qiu, Z.; Zhao, J.; Nie, W.; Lin, H.; Zhu, Z.; Zeng, D.; Qian, Q.; Zhu, L. FRUCTOKINASE-LIKE PROTEIN 1 interacts with TRXz to regulate chloroplast development in rice. J. Integr. Plant Biol. 2018, 60, 94–111. [Google Scholar] [CrossRef]
- Takeuchi, R.; Kimura, S.; Saotome, A.; Sakaguchi, K. Biochemical properties of a plastidial DNA polymerase of rice. Plant Mol. Biol. 2007, 64, 601–611. [Google Scholar] [CrossRef]
- Qiao, Y.; Jiang, W.; Lee, J.; Park, B.; Choi, M.; Piao, R.; Woo, M.; Roh, J.; Han, L.; Paek, N.; et al. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit μ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol. 2009, 185, 258–274. [Google Scholar] [CrossRef]
- Zhao, M.; Guo, Y.; Sun, H.; Dai, J.; Peng, X.; Wu, X.; Yun, H.; Zhang, L.; Qian, Y.; Li, X.; et al. Lesion mimic mutant 8 balances disease resistance and growth in rice. Front. Plant Sci. 2023, 14, 1189926. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Y.; Ma, X.; Meng, L.; Jing, R.; Wang, F.; Wang, S.; Cheng, Z.; Zhang, X.; Jiang, L.; et al. Disruption of gene SPL35, encoding a novel CUE domain-containing protein, leads to cell death and enhanced disease response in rice. Plant Biotechnol. J. 2019, 17, 1679–1693. [Google Scholar] [CrossRef]
- Yang, Y.; Qi, M.; Mei, C. Endogenous salicylic acid protects rice plants from oxidative damage caused by aging as well as biotic and abiotic stress. Plant J. 2004, 40, 909–919. [Google Scholar] [CrossRef]
- Kang, S.G.; Lee, K.E.; Singh, M.; Kumar, P.; Matin, M.N. Rice Lesion Mimic Mutants (LMM): The Current Understanding of Genetic Mutations in the Failure of ROS Scavenging during Lesion Formation. Plants 2021, 10, 1598. [Google Scholar] [CrossRef]
- Yamada, M.; Han, X.; Benfey, P.N. RGF1 controls root meristem size through ROS signalling. Nature 2020, 577, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Berthold, D.A.; Voevodskaya, N.; Stenmark, P.; Gräslund, A.; Nordlund, P. EPR studies of the mitochondrial alternative oxidase: Evidence for a diiron carboxylate center. Perspect. Surg. 2002, 277, 43608–43614. [Google Scholar] [CrossRef]
- Affourtit, C.; Albury, M.S.; Crichton, P.G.; Moore, A.L. Exploring the molecular nature of alternative oxidase regulation and catalysis. FEBS Lett. 2001, 510, 121–126. [Google Scholar] [CrossRef]
- Li, H.; Zhang, H.; Yang, Y.; Fu, G.; Tao, L.; Xiong, J. Effects and oxygen-regulated mechanisms of water management on cadmium (Cd) accumulation in rice (Oryza sativa). Sci. Total Environ. 2022, 846, 157484. [Google Scholar] [CrossRef]
- Murakami, Y.; Toriyama, K. Enhanced high temperature tolerance in transgenic rice seedlings with elevated levels of alternative oxidase, OsAOX1a. Plant Biotechnol. 2008, 25, 361–364. [Google Scholar] [CrossRef]
- Vanlerberghe, G.C.; Cvetkovska, M.; Wang, J. Is the maintenance of homeostatic mitochondrial signaling during stress a physiological role for alternative oxidase? Physiol. Plant. 2009, 137, 392–406. [Google Scholar] [CrossRef]
- Saika, H.; Ohtsu, K.; Hamanaka, S.; Nakazono, M.; Tsutsumi, N.; Hirai, A. AOX1c, a novel rice gene for alternative oxidase; Comparison with rice AOX1a and AOX1b. Genes Genet. Syst. 2002, 77, 31–38. [Google Scholar] [CrossRef]
- Guan, Q.; Takano, T.; Liu, S. Genetic transformation and analysis of rice OsAPx2 gene in Medicago sativa. PLoS ONE 2012, 7, e41233. [Google Scholar] [CrossRef]
- Lin, A.; Wang, Y.; Tang, J.; Xue, P.; Li, C.; Liu, L.; Hu, B.; Yang, F.; Loake, G.J.; Chu, C. Nitric oxide and protein S-nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol. 2011, 158, 451–464. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, J.; Wang, F.; Tang, Y.; Wei, Z.; Ji, Z.; Wang, C.; Zhao, K. Mutation Types of CYP71P1 Cause Different Phenotypes of Mosaic Spot Lesion and Premature Leaf Senescence in Rice. Front. Plant Sci. 2021, 12, 641300. [Google Scholar] [CrossRef] [PubMed]
- Das, B.K.; Xia, L.; Palandjian, L.; Gozani, O.; Chyung, Y.; Reed, R. Characterization of a protein complex containing spliceosomal proteins SAPs 49, 130, 145, and 155. Mol. Cell. Biol. 1999, 19, 6796–6802. [Google Scholar] [CrossRef] [PubMed]
- Golas, M.M.; Sander, B.; Will, C.L.; Lührmann, R.; Stark, H. Molecular architecture of the multiprotein splicing factor SF3b. Science 2003, 300, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Tsuge, T.; Dohmae, N.; Takio, K.; Wei, N. Association of SAP130/SF3b-3 with Cullin-RING ubiquitin ligase complexes and its regulation by the COP9 signalosome. BMC Biochem. 2008, 9, 1. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ishikawa, E.; Sakuma, M.; Hara, H.; Ogata, K.; Saito, T. Mincle is an ITAM-coupled activating receptor that senses damaged cells. Nat. Immunol. 2008, 9, 1179–1188. [Google Scholar] [CrossRef]
- Jincho, Y.; Sotomaru, Y.; Kawahara, M.; Ono, Y.; Ogawa, H.; Obata, Y.; Kono, T. Identification of genes aberrantly expressed in mouse embryonic stem cell-cloned blastocysts. Biol. Reprod. 2008, 78, 568–576. [Google Scholar] [CrossRef]
- Ge, C.-W.; E, Z.-G.; Pan, J.-J.; Jiang, H.; Zhang, X.-Q.; Zeng, D.-L.; Dong, G.-J.; Hu, J.; Xue, D.-W. Map-based cloning of a spotted-leaf mutant gene OsSL5 in Japonica rice. Plant Growth Regul. 2014, 75, 595–603. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Xu, N.; Shui, Y.; Zhang, J.; Yin, W.; Tian, M.; Guo, F.; Bai, D.; Qi, P.; Huang, Q.; et al. Phenotypic Analysis and Gene Cloning of a New Allelic Mutant of SPL5 in Rice. Agriculture 2023, 13, 1875. https://doi.org/10.3390/agriculture13101875
Li P, Xu N, Shui Y, Zhang J, Yin W, Tian M, Guo F, Bai D, Qi P, Huang Q, et al. Phenotypic Analysis and Gene Cloning of a New Allelic Mutant of SPL5 in Rice. Agriculture. 2023; 13(10):1875. https://doi.org/10.3390/agriculture13101875
Chicago/Turabian StyleLi, Ping, Nana Xu, Yang Shui, Jie Zhang, Wuzhong Yin, Min Tian, Faping Guo, Dasong Bai, Pan Qi, Qingxiong Huang, and et al. 2023. "Phenotypic Analysis and Gene Cloning of a New Allelic Mutant of SPL5 in Rice" Agriculture 13, no. 10: 1875. https://doi.org/10.3390/agriculture13101875
APA StyleLi, P., Xu, N., Shui, Y., Zhang, J., Yin, W., Tian, M., Guo, F., Bai, D., Qi, P., Huang, Q., Li, B., Li, Y., Hu, Y., & Peng, Y. (2023). Phenotypic Analysis and Gene Cloning of a New Allelic Mutant of SPL5 in Rice. Agriculture, 13(10), 1875. https://doi.org/10.3390/agriculture13101875