
Pathways and Drivers of Gross N Transformation in Different Soil Types under Long-Term Chemical Fertilizer Treatments


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. 15N Tracing Experiment
2.3. Determination of Mineral N Concentration and 15N Enrichment
2.4. Quantification of Gross N Transformation Rates
2.5. Calculations and Statistical Analyses
3. Results
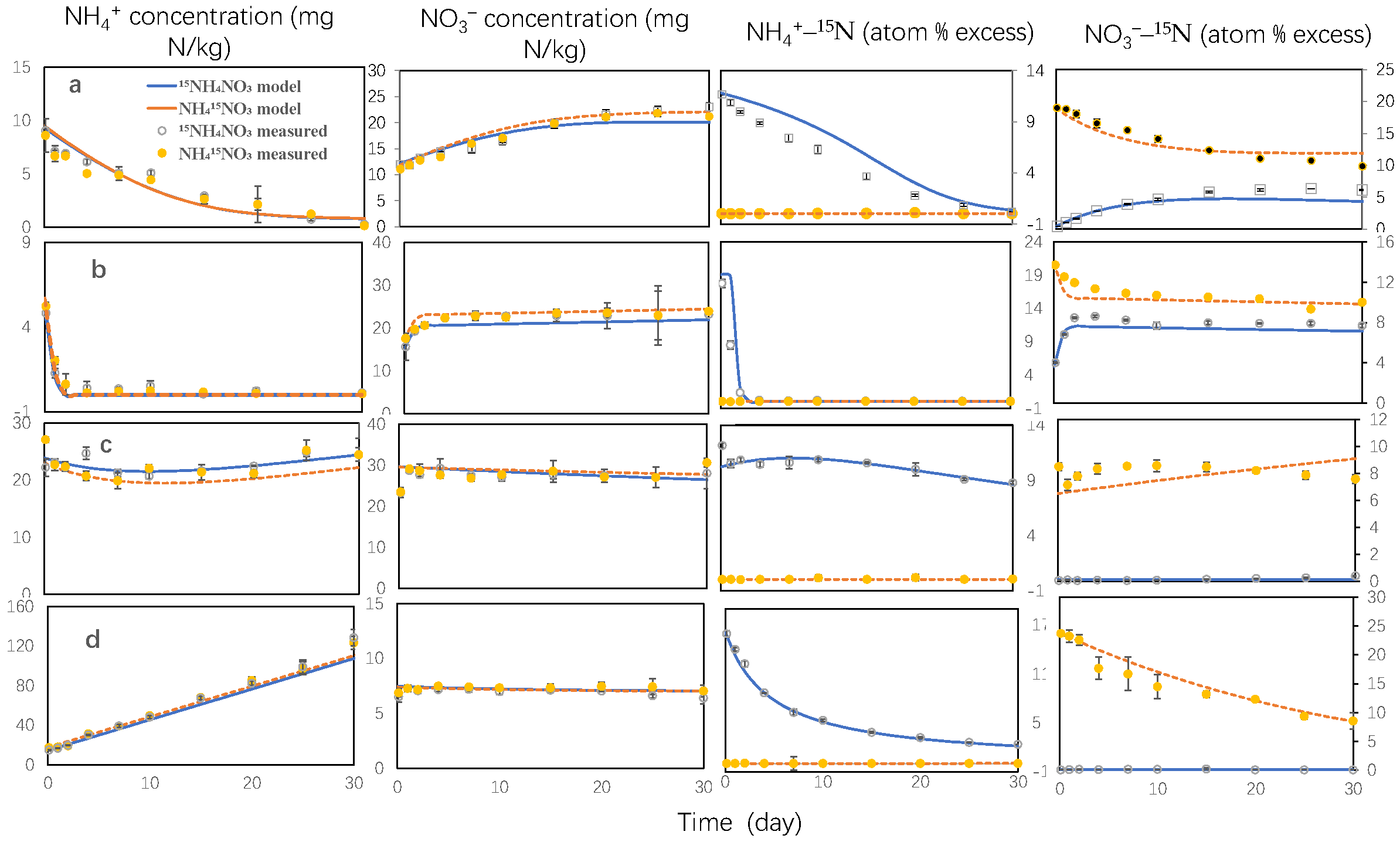
3.1. Inorganic Nitrogen Pool Sizes and 15N Enrichment
3.2. Gross N Transformation Rates
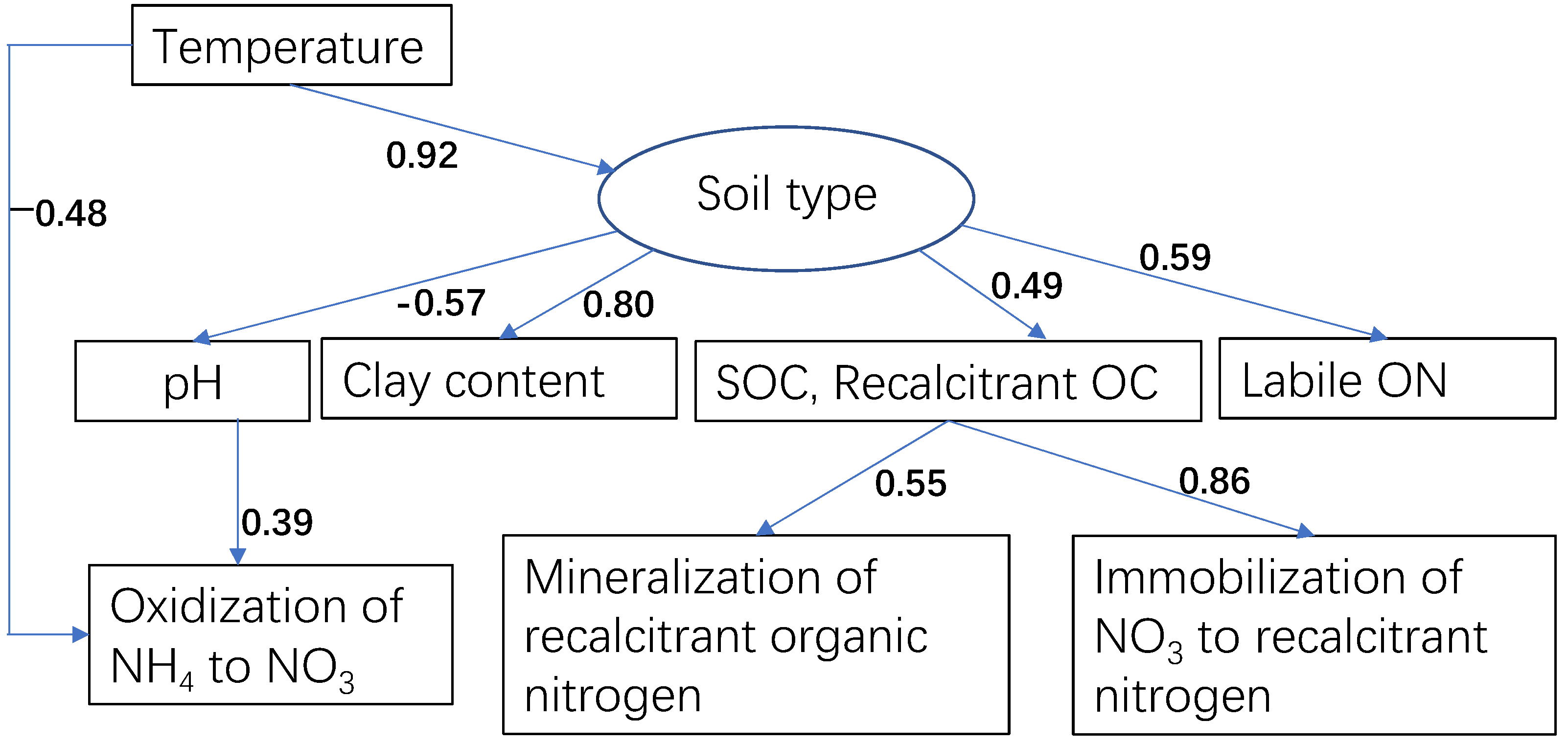
3.3. PLS-PM Analysis
4. Discussion
4.1. Patterns of Gross N Mineralization Rates and Affecting Factors
4.2. Patterns of Gross N Nitrification Rate and Affecting Factors
4.3. Factors Affecting the Immobilization of NO3−
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elrys, A.S.; Ali, A.; Zhang, H.; Cheng, Y.; Zhang, J.; Cai, Z.; Müller, C.; Chang, S.X. Patterns and drivers of global gross nitrogen mineralization in soils. Glob. Chang. Biol. Glob. Chang. Biol. 2021, 27, 5950–5962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhou, M.; Zhu, B.; Xiao, Q.; Zheng, X.; Zhang, J.; Müller, C.; Butterbach-Bahl, K. Soil clay minerals: An overlooked mediator of gross N transformations in Regosolic soils of subtropical montane landscapes. Soil Biol. Biochem. 2022, 168. [Google Scholar] [CrossRef]
- Bossio, D.; Scow, K.; Gunapala, N.; Graham, K. Determinants of Soil Microbial Communities: Effects of Agricultural Management, Season, and Soil Type on Phospholipid Fatty Acid Profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, X.; Li, S.; Yin, X. Long-Term Flooding Paddy Affects Inorganic Nitrogen Supply and Conservation Processes in Subtropical Soils of Southwest China. J. Soil Sci. Plant Nutr. 2022, 22, 2049–2059. [Google Scholar] [CrossRef]
- Hou, H.; Liu, X.; Zhou, W.; Ji, J.; Lan, X.; Lv, Z.; Liu, Y.; Zhang, J.; Müller, C. N transformation mechanisms and N dynamics of organic fertilisers as partial substitutes for chemical fertilisers in paddy soils. J. Soils Sediments 2022, 22, 2516–2529. [Google Scholar] [CrossRef]
- Lan, T.; Han, Y.; Cai, Z. Comparison of Gross N Transformation Rates in Two Paddy Soils Under Aerobic Condition. Pedosphere 2017, 27, 112–120. [Google Scholar] [CrossRef]
- Nishio, T.; Sekiya, H.; Toriyama, K.; Kogano, K. Changes in gross rates of nitrogen transformations in soil caused by con-version of paddy fields to upland fields. Soil Sci. Plant Nutr. 1994, 40, 301–309. [Google Scholar] [CrossRef]
- Kader, M.A.; Sleutel, S.; Begum, S.A.; Moslehuddin, A.Z.M.; De Neve, S. Nitrogen mineralization in sub-tropical paddy soils in relation to soil mineralogy, management, pH, carbon, nitrogen and iron contents. Eur. J. Soil Sci. 2013, 64, 47–57. [Google Scholar] [CrossRef]
- Zhang, J.B.; Cai, Z.C.; Yang, W.Y.; Zhu, T.B.; Yu, Y.J.; Yan, X.Y. Long-term field fertilization affects soil nitrogen transformations in a rice-wheat-rotation cropping system. J. Plant Nutr. Soil Sci. 2012, 175, 939–946. [Google Scholar] [CrossRef]
- Lang, M.; Li, P.; Han, X.; Qiao, Y.; Miao, S. Gross nitrogen transformations in black soil under different land uses and management systems. Biol. Fertil. Soils 2015, 52, 233–241. [Google Scholar] [CrossRef]
- Li, P.; Lang, M. Gross nitrogen transformations and related N2O emissions in uncultivated and cultivated black soil. Biol. Fertil. Soils 2013, 50, 197–206. [Google Scholar] [CrossRef]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Wang, B.; Wang, S.; Meng, F.; Lu, C. Characteristics of soil labile organic carbon and carbon management index under different long-term fertilization systems in four typical soils of China. Soil Fertil. Sci. China 2018, 2, 29–34. [Google Scholar]
- Lteif, A.; Whalen, J.K.; Bradley, R.L.; Camiré, C. Nitrogen transformations revealed by isotope dilution in an organically fertilized hybrid poplar plantation. Plant Soil 2010, 333, 105–116. [Google Scholar] [CrossRef]
- Müller, C.; Rütting, T.; Kattge, J.; Laughlin, R.; Stevens, R. Estimation of parameters in complex 15N tracing models by Monte Carlo sampling. Soil Biol. Biochem. 2007, 39, 715–726. [Google Scholar] [CrossRef]
- Ai, C.; Zhang, S.; Zhang, X.; Guo, D.; Zhou, W.; Huang, S. Distinct responses of soil bacterial and fungal communities to changes in fertilization regime and crop rotation. Geoderma 2018, 319, 156–166. [Google Scholar] [CrossRef]
- Barrett, J.; Burke, I. Potential nitrogen immobilization in grassland soils across a soil organic matter gradient. Soil Biol. Biochem. 2000, 32, 1707–1716. [Google Scholar] [CrossRef]
- Müller, C.; Laughlin, R.J.; Christie, P.; Watson, C.J. Effects of repeated fertilizer and cattle slurry applications over 38 years on N dynamics in a temperate grassland soil. Soil Biol. Biochem. 2011, 43, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Rütting, T.; Clough, T.J.; Müller, C.; Lieffering, M.; Newton, P.C.D. Ten years of elevated atmospheric carbon dioxide alters soil nitrogen transformations in a sheep-grazed pasture. Glob. Chang. Biol. 2009, 16, 2530–2542. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, B.; Zhang, J.; Müller, C.; Cai, Z. Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China. Soil Biol. Biochem. 2015, 91, 222–231. [Google Scholar] [CrossRef]
- Huang, X.; Zhu-Barker, X.; Horwath, W.R.; Faeflen, S.J.; Luo, H.; Xin, X.; Jiang, X. Effect of iron oxide on nitrification in two agricultural soils with different pH. Biogeosciences 2016, 13, 5609–5617. [Google Scholar] [CrossRef] [Green Version]
- Elrys, A.S.; Wang, J.; Metwally, M.A.S.; Cheng, Y.; Zhang, J.; Cai, Z.; Chang, S.X.; Müller, C. Global gross nitrification rates are dominantly driven by soil carbon-to-nitrogen stoichiometry and total nitrogen. Glob. Chang. Biol. 2021, 27, 6512–6524. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, X.M.; Deng, J.Q.; Lin, J.; Xia, W. Charge properties in three kinds of red soils from different parent materials. Soils 2011, 43, 481–486, (In Chinese with English abstract). [Google Scholar]
- Bannert, A.; Kleineidam, K.; Wissing, L.; Müller-Niggemann, C.; Vogelsang, V.; Welzl, G.; Cao, Z.H.; Schloter, M. Changes in diversity and functional gene abundances of microbial communities involved in nitrogen fixation, nitrification and denitri-fication comparing a tidal wetland to paddy soils cultivated for different time periods. Appl. Environ. Microbiol. 2011, 77, 6109–6116. [Google Scholar] [CrossRef] [Green Version]
- Robertson, G.P.; Groffman, P.M. Nitrogen transformation. In Soil Microbiology, Biochemistry, and Ecology; Paul, E.A., Ed.; Springer: New York, NY, USA, 2007; pp. 341–364. [Google Scholar]
- Cookson, W.; Osman, M.; Marschner, P.; Abaye, D.; Clark, I.; Murphy, D.; Stockdale, E.; Watson, C. Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol. Biochem. 2007, 39, 744–756. [Google Scholar] [CrossRef]
- Gubry-Rangin, C.; Hai, B.; Quince, C.; Engel, M.; Thomson, B.C.; James, P.; Schloter, M.; Griffiths, R.I.; Prosser, J.I.; Nicol, G.W. Niche specialization of terrestrial archaeal ammonia oxidizers. Proc. Natl. Acad. Sci. USA 2011, 108, 21206–21211. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.J.; Ling, N.; Chen, H.; Zhu, C.; Kong, Y.L.; Wang, M.; Shen, Q.R.; Guo, S.W. Distinct drivers of activity, abundance, diversity and composition of ammonia–oxidizers: Evidence from a long-term field experiment. Soil Biol. Biochem. 2017, 115, 403–414. [Google Scholar] [CrossRef]
- Shen, J.P.; Zhang, L.M.; Di, H.J.; He, J.Z. A review of ammonia-oxidizing bacteria and archaea in Chinese soils. Front. Microbiol. 2012, 3, 00296. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.Y.; Liu, Q.; Zhao, J.; Zhang, J.B. Ecological niche differentiation of ammonia-oxidizing archaea and bacteria in acidic soils due to land use change. Soil Res. 2018, 56, 71–79. [Google Scholar] [CrossRef]
- Daebeler, A.; Bodelier, P.L.; Hefting, M.M.; Rütting, T.; Jia, Z.; Laanbroek, H.J. Soil warming and fertilization altered rates of nitrogen transformation processes and selected for adapted ammonia-oxidizing archaea in sub-arctic grassland soil. Soil Biol. Biochem. 2017, 107, 114–124. [Google Scholar] [CrossRef]
- Brierley, E.; Wood, M. Heterotrophic nitrification in an acid forest soil: Isolation and characterisation of a nitrifying bacterium. Soil Biol. Biochem. 2001, 33, 1403–1409. [Google Scholar] [CrossRef]
- Zhao, B.; Li, X.; Liu, H.; Wang, B.; Zhu, P.; Huang, S.; Bao, D.; Li, Y.; So, H. Results from long-term fertilizer experiments in China: The risk of groundwater pollution by nitrate. NJAS-Wagening. J. Life Sci. 2011, 58, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Rice, C.W.; Tiedje, J.M. Regulation of nitrate assimilation by ammonium in soils and in isolated soil microorganisms. Soil Biol. Biochem. 1989, 21, 597–602. [Google Scholar] [CrossRef]
- Burger, M.; Jackson, L.E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems. Soil Biol. Biochem. 2003, 35, 29–36. [Google Scholar] [CrossRef]
- Elrys, A.S.; Chen, Z.; Wang, J.; Uwiragiye, Y.; Helmy, A.M.; Desoky, E.M.; Cheng, Y.; Zhang, J.; Cai, Z.; Müller, C. Global patterns of soil gross immobilization of ammonium and nitrate in terrestrial ecosystems. Glob. Chang. Biol. 2022, 28, 4472–4488. [Google Scholar] [CrossRef] [PubMed]
- Niboyet, A.; Barthes, L.; Hungate, B.A.; LE Roux, X.; Bloor, J.M.G.; Ambroise, A.; Fontaine, S.; Price, P.M.; Leadley, P.W. Responses of soil nitrogen cycling to the interactive effects of elevated CO2 and inorganic N supply. Plant Soil 2009, 327, 35–47. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Black Soil Chernozem (WRB) * | Fluvo-Aquic Soil Calcaric-Fluvisols | Upland Red Soil Ferralic Cambisol | Paddy Red Soil Ferralic Cambisol | |
|---|---|---|---|---|
| Parent material | Quaternary sediments | River Alluvium | Quaternary red clay | Quaternary red clay |
| Clay mineral type # | Montmorillonite | Hydromica Montmorillonite | Kaolinite | Kaolinite |
| Clay content (%) | 31.1 | 20 | 43.9 | 38.7 |
| SOC (g kg−1) | 12.6 | 7.3 | 8.7 | 21.3 |
| Total N (g kg−1) | 1.34 | 0.67 | 1.02 | 2.11 |
| pH | 5.9 | 7.9 | 4.6 | 5.4 |
| Temperature (°C) | 4.5 | 14.4 | 18 | 17 |
| Rainfall (mm) | 525 | 700 | 1255 | 1370 |
| N Treatments | Black Soil | Fluvo-Aquic Soil | Upland Red Soil | Paddy Red Soil | |||
|---|---|---|---|---|---|---|---|
| Maize | Wheat | Maize | Wheat | Maize | Early Rice | Late Rice | |
| N (urea) | 165 | 165 | 188 | 90 | 210 | 165 | 165 |
| P (calcium superphosphate) | 36 | 36 | 41 | 16 | 37 | 45 | 45 |
| K (potassium chloride) | 68 | 68 | 78 | 30 | 70 | 120 | 120 |
| Black Upland Soil | Fluvo-Aquic Upland Soil | Red Upland Soil | Red Paddy Soil | |||||
|---|---|---|---|---|---|---|---|---|
| K * | Mean | K | Mean | K | Mean | K | Mean | |
| MNrec | 0 | 0.145 ± 0.005 c | 0 | 0.037 ± 0.003 d | 0 | 0.724 ± 0.035 b | 0 | 3.096 ± 0.020 a |
| INH4 | 1 | 0.003 ± 0.001 b | 1 | 0.000 ± 0 b | 1 | 0.272 ± 0.086 a | 1 | 0.003 ± 0.002 b |
| MNlab | 1 | 0.054 ± 0.009 a | 1 | 0.012 ± 0.002 b | 1 | 0.036 ± 0.297 b | 1 | 0.017 ± 0.009 b |
| ONrec | 0 | 0.000 ± 0.000 b | 0 | 0.007 ± 0.000 b | 0 | 0.002 ± 0.008 b | 0 | 0.238 ± 0.004 a |
| INO3 | 1 | 0.193 ± 0.014 b | 1 | 0.000 ± 0.000 c | 2 | 0.010 ± 0.006 c | 1 | 0.259 ± 0.005 a |
| ONH4 | 2 | 0.467 ± 0.012 a | 2 | 0.223 ± 0.016 b | 2 | 0.000 ± 0.000 c | 1 | 0.010 ± 0.001 c |
| ANH4 | 1 | 0.000 ± 0.000 b | 1 | 0 ± 0 b | 1 | 1.204 ± 0.128 a | 1 | 0.000 ± 0.005 b |
| RNH4 | 1 | 0 ± 0 b | 1 | 0 ± 0 b | 1 | 0.715 ± 0.045 a | 1 | 0 ± 0 b |
| ANO3 | 1 | 0.029 ± 0.009 b | 1 | 0 ± 0 c | 1 | 0.110 ± 0.013 a | 1 | 0 ± 0 c |
| RNO3 | 1 | 0.039 ± 0.002 a | 1 | 0 ± 0 b | 1 | 0.040 ± 0.003 a | 1 | 0 ± 0 b |
| NC (%) | 235 | 485 | 1 | 8 | ||||
| AIC | 5359 | 3600 | 583 | 207 | ||||
| R2 | 0.96 | 0.99 | 0.96 | 0.99 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Yu, W.; Meng, F.; Zhang, J.; Lu, C. Pathways and Drivers of Gross N Transformation in Different Soil Types under Long-Term Chemical Fertilizer Treatments. Agriculture 2023, 13, 300. https://doi.org/10.3390/agriculture13020300
Li G, Yu W, Meng F, Zhang J, Lu C. Pathways and Drivers of Gross N Transformation in Different Soil Types under Long-Term Chemical Fertilizer Treatments. Agriculture. 2023; 13(2):300. https://doi.org/10.3390/agriculture13020300
Chicago/Turabian StyleLi, Guihua, Weishui Yu, Fanhua Meng, Jianfeng Zhang, and Changai Lu. 2023. "Pathways and Drivers of Gross N Transformation in Different Soil Types under Long-Term Chemical Fertilizer Treatments" Agriculture 13, no. 2: 300. https://doi.org/10.3390/agriculture13020300
APA StyleLi, G., Yu, W., Meng, F., Zhang, J., & Lu, C. (2023). Pathways and Drivers of Gross N Transformation in Different Soil Types under Long-Term Chemical Fertilizer Treatments. Agriculture, 13(2), 300. https://doi.org/10.3390/agriculture13020300