1. Introduction
An ever-increasing world population and climate change are among the major contributors to famine. Crop yields must continue to increase to attain the Sustainable Development Goals and ensure safe, nutritious, and adequate food for everyone. Cereals and derivatives should play a key role in this challenge since they are the staple food of most countries of the world and the primary food for livestock. Cereal grains constitute more than half of the food energy and half of the protein consumed on Earth [
1]. With the pressure on resources and the climate change scenario, future food supplies not only need to be increased but also enriched especially in nutrients and protein to address food and nutrient security. Nutrition deficiencies have a significant impact on a country’s productivity and can reduce its gross domestic product (GDP) by as much as 7% globally and up to 9–10% in African and South Asian countries [
2]. In this context, there is a need to develop new sustainable sources of protein and nutrients to avoid the 150 more people expected to be at risk of protein deficiencies by 2050 [
3]. It is evident that protein plays an essential role within the human body, including building muscle and bone and brain development. Its components, amino acids, are the building blocks of this development and are critically important in the first 1000 days of life and throughout the lifecycle [
4]. Animal-based foods are excellent sources of protein but contribute 14.5% of all greenhouse gas emissions [
5]. In this view, development of new nutrient-rich crops such as high-protein oat,
Avena magna ssp.
domestica, becomes one of the pathways to diversify the sources of protein and help attack the dual challenge of increasing supply in a sustainable manner.
Oat ranks sixth in world cereal production after wheat, maize, rice, barley, and sorghum [
6]. The most widespread cultivated species of oat is common oat (
Avena sativa), with Abyssinian oat (
A. abyssinica), diploid naked or hulless oats (
A. nuda), and lopsided oat (
A. strigosa) being of relatively minor importance [
7]. Morocco is the center of origin and a major center of diversity of the
Avena genus [
8]; consequently, it represents an essential country for expanding genetic resource conservation efforts and an interesting location for expanding oat production.
Oat is a cereal that has long been prized for its nutritional attributes, particularly in terms of mixed-linkage soluble beta-glucan, protein, oil, and antioxidants. Oat grain is also free-gluten and has the highest protein content among cereals; as a consequence, it is tolerated by most celiac disease patients [
9]. Several previous studies have shown oat’s health-promoting effects in preventing obesity, type II diabetes, gastrointestinal diseases, coronary heart disease, and certain types of cancer [
10]. For human consumption, an increasing supply of oat-based products has been exhibited in the market, including a diverse array of breads, oatcakes, biscuits, granola bars, and ever more novel foods such as yogurt-type products and oat-based drinks [
11,
12,
13].
The newly domesticated Moroccan tetraploid species
A. magna (2n = 4x = 28, CCDD genome) was developed via sexual transfer of the domestication syndrome from the hexaploid common oat
A. sativa (2n = 6x = 42, AACCDD; [
14]). Subsequently, Jackson [
7] produced a set of
A. magna ssp.
domestica lines via mutagenesis within a population of F
2:8 recombinant inbred lines (RILs) from a cross between a stable domesticated backcross line, Ba13-13, and a wild
A. magna parent [
15]. Oliver et al. [
15] also mapped three of the domestication syndrome genes, for shattering (
Ba), geniculate awn (
Awn), and lemma pubescence (
Lp), and measured the linkage of
Awn and
Ba with a terminal knob on chromosome 2C at a distance of 2.1 cM.
While the lines developed by Jackson [
7] are suitable for production under standard cultivation practices, they also possess much higher protein levels than common oats, exceeding 25% in the relatively new cultivar ‘Avery’.
Avena magna ssp.
domestica also showed promise in improving dietary iron and zinc in comparison to other cereals, thus potentially reducing anemia [
16,
17]. These characteristics make
A. magna a good grain resource for the development of high-quality protein products to address developing-world malnutrition, developed-world obesity, and high-quality livestock feed [
11]. Since
A. magna is already adapted to the semi-arid conditions of Morocco, it is a promising source of sustainable grain for a world increasingly affected by climate change. Nevertheless, there is still a need to continue breeding under Moroccan conditions for traits such as resistance to seed dehiscence, lodging, dormancy, productivity, and stability, as well as tolerance of diseases.
The main objective of this study was to assess the diversity and performance of A. magna ssp. domestica lines using ‘Avery’ as the control in seven diverse environments in Morocco. The evaluation included morphological and yield parameters, disease tolerance (crown rust, BYDV), and yield stability.
4. Discussion
The goal of this present work was to determine the agro-morphological variability of 11
A. magna ssp.
domestica advanced lines and a control cultivar named Avery (ATC).
Avena sativa (2n = 6x = 42) is the principal species of oat cultivated in Morocco, usually for fodder [
25]. At the beginning of the local oat research program of the National Institute of Agricultural Research, varieties derived from the native hexaploid
A. byzantina (red oat), which has a low water requirement and the ability to adjust its growth cycle to Moroccan environmental variability, were also released [
26]. The array of wild
Avena species native to Morocco—the center of origin of the genus—unfortunately lacks the required traits for broad cultivation such as resistance to shattering, semi-dwarfism, lodging resistance, erect growth habit, resistance to seed dormancy, etc. The neo-domesticated
A. magna material first described by Ladizinsky [
14] and later modified by Jackson [
7] should have the potential to diversify the cultivated germplasm of oats. This new oat diversity contributes to the conservation of genetic resources of the genus and plays a positive role in maintaining crop diversity [
6,
17]. In addition, the significantly higher protein content of ‘Avery’ previously reported by Jackson [
7] could encourage farmers and manufacturers to harness this asset to improve the crop’s profitability for both human and animal feed. Over the course of the past 40 years, interest in oats for human consumption has gradually increased, concomitant with mounting clinical evidence regarding its nutritional benefits for cardiovascular health [
27].
The disease assessment study had focused only on RC and BYDV, since there were no noticeable symptoms of powdery mildew at the heading stage when scoring was carried out. Several authors agree that RC (
Puccinia coronata) is the major disease threatening global oat production [
28]. This pathogen can drastically affect oat production by causing 10–40% yield loss [
29]. Crown rust (RC) was only recorded at Bouchane in the 2018–2019 cropping season because at the other locations there were no noticeable symptoms in the experimental plots. The cause may be due to relatively dry and cool conditions at the experimental sites during 2020–2021. Nazareno et al. [
28] reported that crown rust epidemics usually occur in warm areas as temperatures fluctuate between 20 and 25 °C and humidity surpasses 50–60%. At the El Kbab high-altitude site and in Alnif’s dry environment, weather conditions would be expected to suppress crown rust development. The incidence of BYDV was more common than RC across the experimental locations. BYDV is probably the most economically important small-grain virus transmitted by insects, with 25 aphid species being involved as potential BYDV vectors [
30]. High light intensity and low temperatures (15–18 °C) also promote increased intensity of BYDV symptoms [
31]. All of these considerations and
A. magna’s well-known susceptibility to BYDV [
14] may explain the high incidence of the virus observed in our experiments. The trials’ results did not show consistent tolerance to diseases of tested lines at all sites, except for line AT6, which appeared to be more tolerant to BYDV and RC infections. Combining data from all sites, PCA revealed that the control variety Avery and AT6 were relatively less susceptible to the diseases, which was not the case for AT7 and AT9. Jellen et al. [
19] had already noted A02 (line AT7 in this study) among the lines showing RC symptoms in the Lahri experiment in 2018.
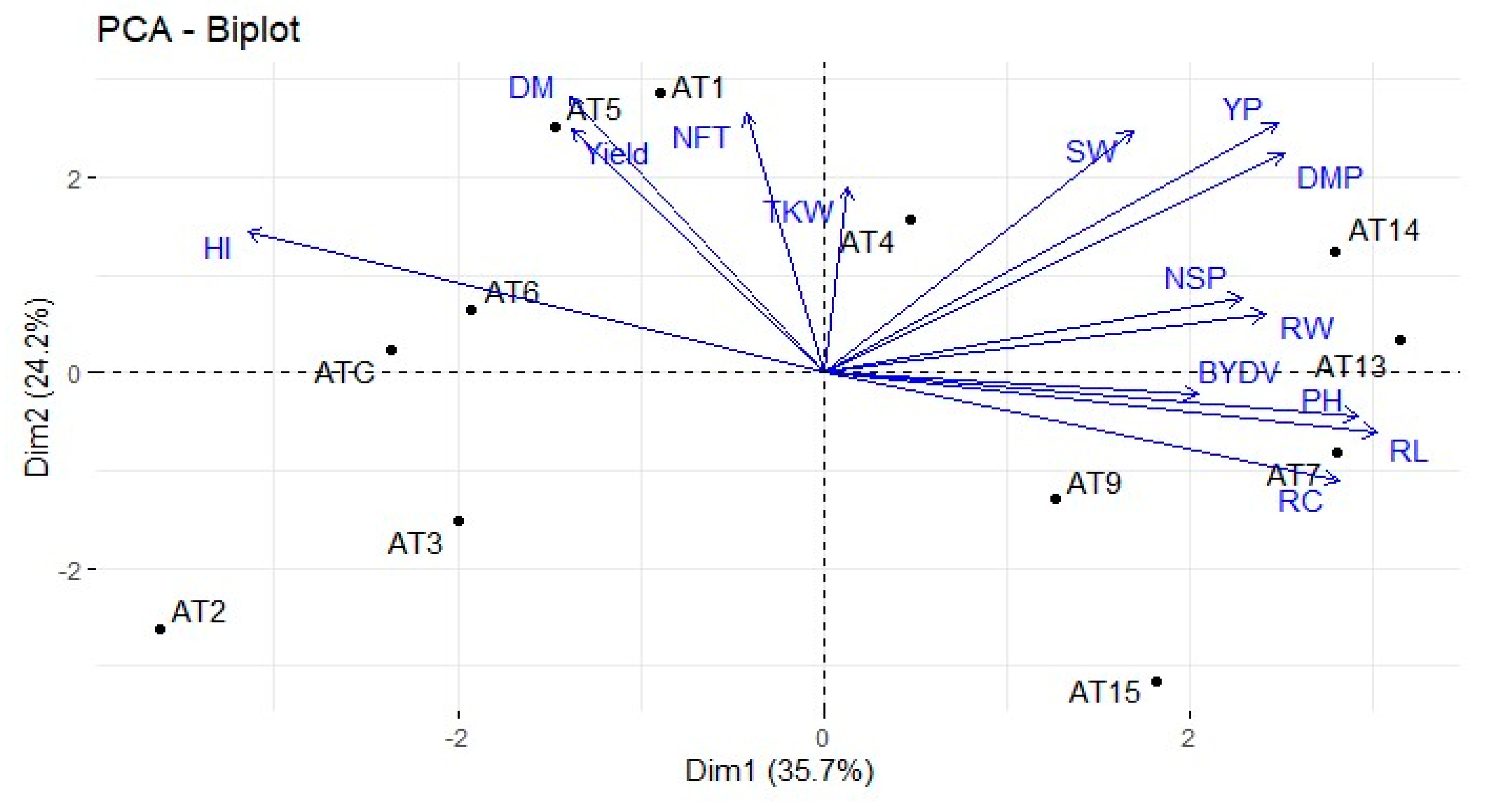
Data analyses revealed significant agro-morphological diversity between the domesticated
A. magna lines. Plant height (PH) varied between 48.93 and 120.47 cm across the sites. Through the different sites, PH fluctuated between 84.07 cm for AT2 and 94.92 cm for AT15. Previous studies reported similar plant height values. Dumlupinar et al. [
32], who studied
A. sativa pure-line varieties, reported variation for plant height between 46 and 112 cm. Among our investigated traits, PH gave the lowest CV, between 9.97 and 21.18% across the five sites. This finding is in line with Solange et al. [
33] and Dumlupinar et al. [
32], who reported CVs of 11.16 and 8.7%, respectively, for this trait. A possible explanation for these observations is that plant height is affected by a relatively small number of major genes, which in this case might be largely fixed through selection. PCA analysis showed that PH was positively correlated with RL (0.72) and NSP (0.60); on the other hand, PH was significantly negatively correlated with HI. Other authors reported a significant negative correlation between PH and grain yield and explained the relationship as being due to lodging [
34,
35]. Plant size has to be examined closely in varietal selection to avoid lodging, in our case with lines AT13 and AT15.
Root length across locations ranged from 11.87 to 16.16 cm at Oukaimeden_21 and Bouchane_19, respectively. The longest roots were observed in Bouchane in three successive seasons. This behavior is likely due to the sandy soil at Bouchane, which facilitates and encourages deep root penetration as capillary water drains downward through the soil profile. Finer roots were also more easily retained as the plants were uprooted in the light-textured soil to take RL measurements. The higher clay content at the other experimental sites allowed for better soil water retention. This supports the previous observation that A.
magna ssp.
domestica has the ability to deeply penetrate the soil under water-stress conditions [
19].
The three-way ANOVA detected significant differences among lines according to their tillering ability: 4.03 NFT/plant for AT15 and 5.93 for AT1 on average (
Table 7). The genetic control of tillering capacity has been discussed previously in oat genotypes [
36,
37]. The mean NFT was mostly equivalent among our test locations except for Alnif_21, where it reached an average of 11.09. Good crop management practices such as irrigation and fertilization have a positive influence on the tillering ability of plants; this may explain the significantly higher NFT value at this irrigated site.
The NSP varied among lines, ranging from 24.85 to 36.06 for AT2 and AT7, respectively. Solange et al. [
33] observed an NSP mean of 26.63 for
A. sativa, which is close to our findings. However, Jan et al. [
6] counted 66 spikelets per panicle on average in their experiment. The number of spikelets is influenced by soil fertility, humidity, and photoperiod.
Thousand-kernel weight (TKW) is an important productivity and grain-quality trait for oats. According to our three-way ANOVA, TKW was more significantly influenced by growing location than genotype. Considering separately the experimental sites, two-way ANOVA showed a significant effect of the accessions at all testing sites except Bouchane_19. Our line TKW values, similar to previous results in
A. sativa, varied between 32.83 and 49.73 g for AT3 and AT5, respectively. Our results correspond well with
A. sativa TKW values [
6,
32,
33].
Seed partitioning on the total biomass is evaluated through the HI. Statistical analyses showed that HI was influenced by both the line and the environment; it varied between 20.43 and 31.33% across the five testing locations. Regarding the effect of the lines, the HI varied between 19.88% for AT13 and 34.76% for AT5. Jan et al. [
6] and Li et al. [
38] reported on variation for HI among
A. sativa genotypes and between environments and years; their HI values were comparable to ours.
It has historically been a major challenge for plant breeders to consistently identify superior-yielding genotypes across environments when GEI is highly significant. Several researchers testing crops in multi-location conditions have reported larger environmental than genotypic effects on grain yield [
39,
40,
41,
42]. In our case, the environmental effect represented 78% of the total grain yield variation, while the genotype explained only 4%. Grain yield varied extensively among our sites: from 6.89 q/ha at Bouchane_19 to 85.50 q/ha at Alnif_21. Previously, within a larger set of
A. magna lines, Jellen et al. [
19] identified the environment as the most important factor influencing yield, with genotype being the second most significant factor. Our AMMI analysis allowed classification of the
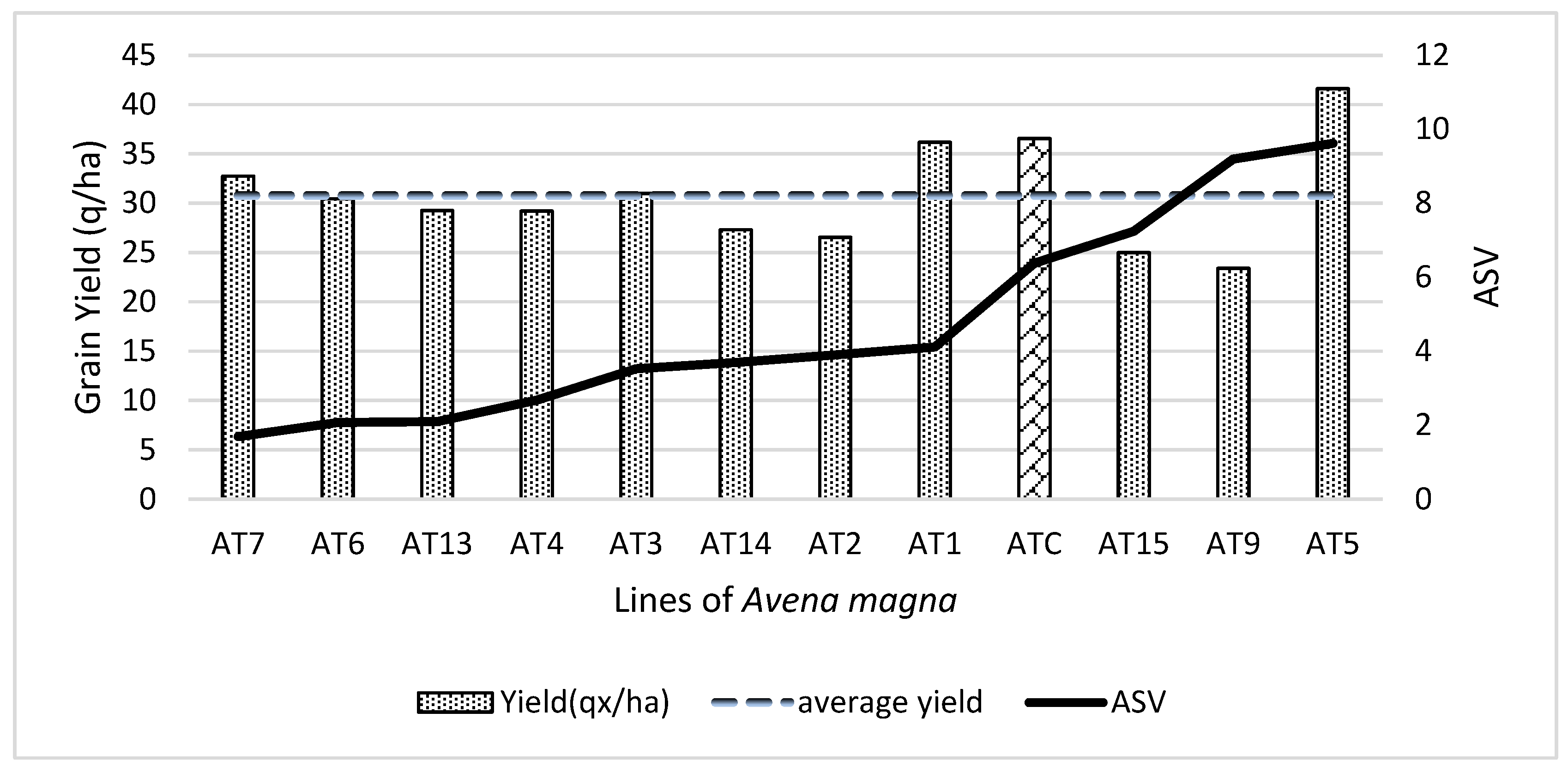
A. magna lines based on their ASVs. Combining ASV and yield into a single index should represent a better way to evaluate a genotype’s potential across locations. The yield stability index has been used by several authors to rank their germplasm [
40,
43,
44]. Stable lines are usually less productive than low-stability ones. Stable genotypes have large adaptation and intermediate yield potential, while low-stability lines yield the maximum in the environment in which they are best adapted while ranking among the lowest in other locations. In this study, we presented a hybrid graph to display the
A. magna line performance and separated them into two groups according to their ASV and grain yield. The first group included
A. magna lines whose grain yield across locations exceeded the average yield of 30.76 q/ha. Two of the genotypes produced relatively higher grain yields, but they were unstable: AT5 (mean yield of 43.31 q/ha) Avery (35.66 q/ha). Line AT5 performed the best at El Kbab_19 while Avery produced the most at Alnif_21, with respective yields of 103.17 q/ha and 114.9 q/ha. Gadisa et al. [
40], while investigating germplasm consisting of 15 bread wheat lines, noticed that the two highest-yielding genotypes were among the most unstable. Each of these two wheat lines had interacted positively at a given site. These results are in agreement with our findings. Among the tested
A. magna lines, AT1 was moderately stable, while AT7 had the highest stability in the collection. Using the yield stability index, AT7 ranked first as being the superior genotype among the tested germplasm in terms of stability and grain yield. This accession, however, appeared to be less tolerant to diseases. Among the
A. magna lines that produced below the mean grain yield, AT6 exhibited good yield stability and will be further scrutinized for its apparent tolerance to the main common diseases and its yield, which approached 30.44 q/ha.
The average grain yield of this present study across five sites and three years of experimentation was 30,76 q/ha. It may be recalled when Tam [
45] tested 101 oat varieties of
A. sativa from Germany, Sweden, Russia, Canada, the USA, and other countries, where the average yields ranged between 32.88 and 58.24 q/ha. In our trials, Bouchane’s relatively low average yield of 6.89 q/ha is explained by the semi-arid climate conditions; moreover, Morocco is an arid to semi-arid country, unlike Estonia where Tam carried out his experiment. At the most favorable Moroccan site, Ain Itto_21, rainfall barely reached 609.8 mm during the cropping season. However, it should be noted that with certain lines at certain locations, grain yield greatly exceeded the above-mentioned yield range: for example, AT5 at El Kbab_19 and Avery at Alnif_21. Tests of various
A. sativa varieties selected for northern Morocco reached 45 q/ha under experimental conditions and 24 q/ha in farmers’ production fields [
46]; these previous results confirm that grain yield tends to decrease under working farm conditions, and some
A. magna lines might express their best yield potential under good cultivation and environmental conditions.




 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}