Sorption to Biochar Impacts β-Glucosidase and Phosphatase Enzyme Activities


Abstract
:1. Introduction
2. Materials and Methods
2.1. Concentrated Enzymes
2.2. Solid Phases
2.3. Enzyme Sorption Assay
2.4. Enzyme Activity Assay
2.5. Assay of Substrate Stability and Sorption
2.6. Statistics
3. Results
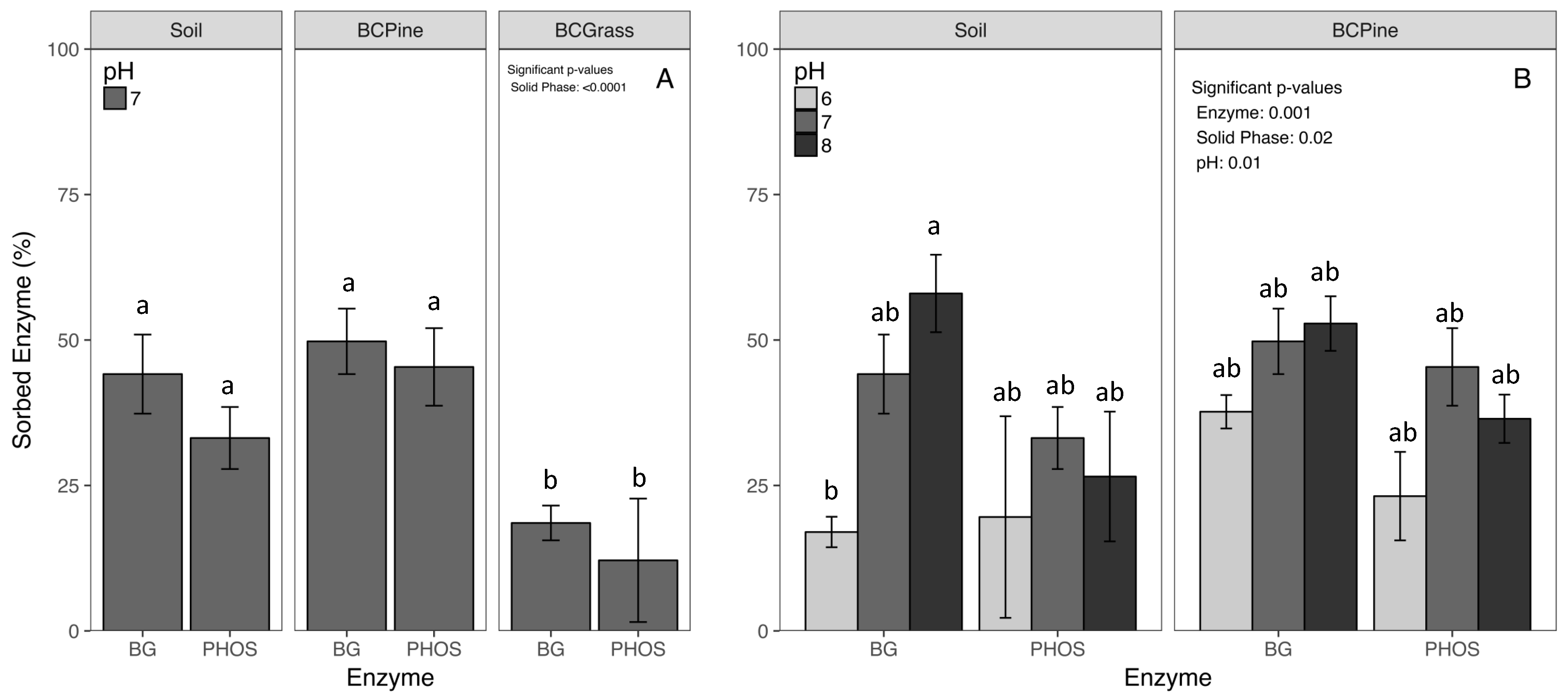
3.1. Enzyme Sorption
3.2. Enzyme Activity
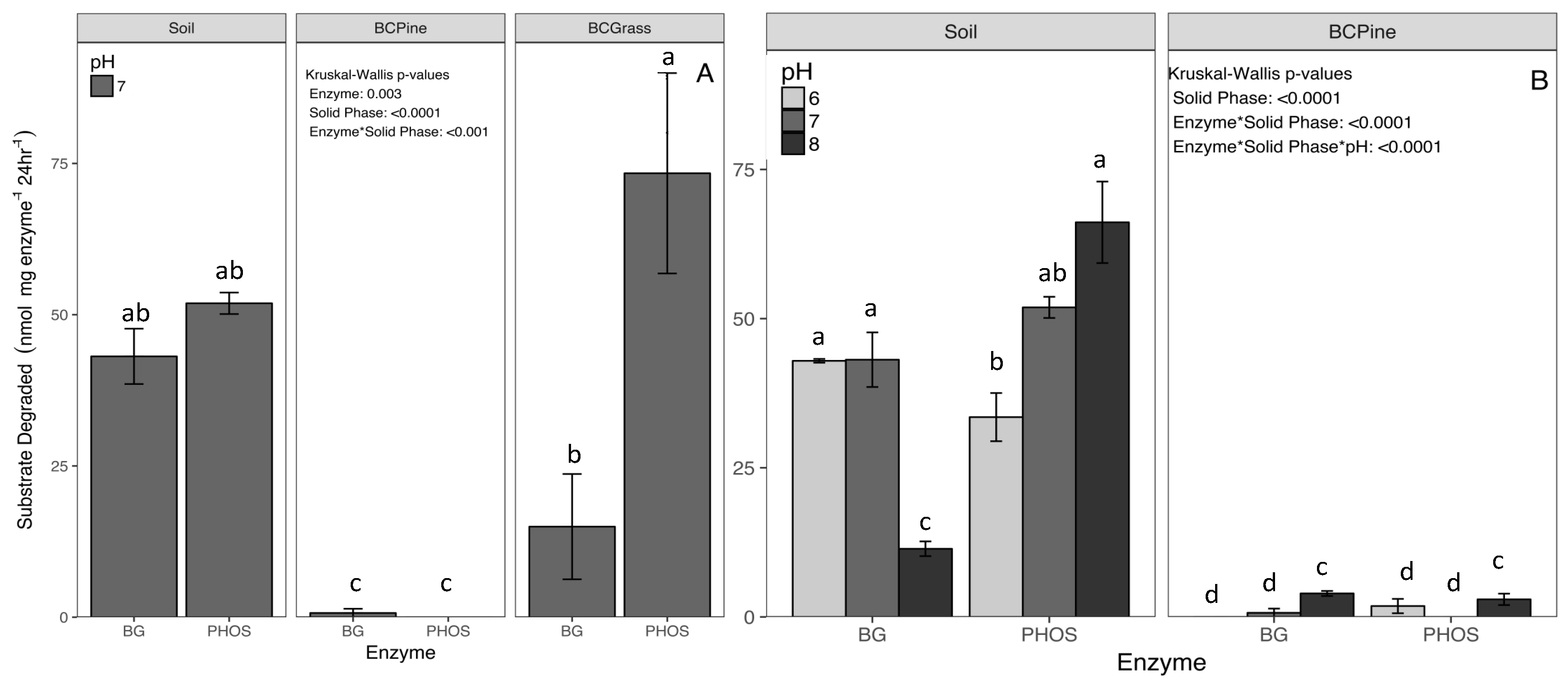
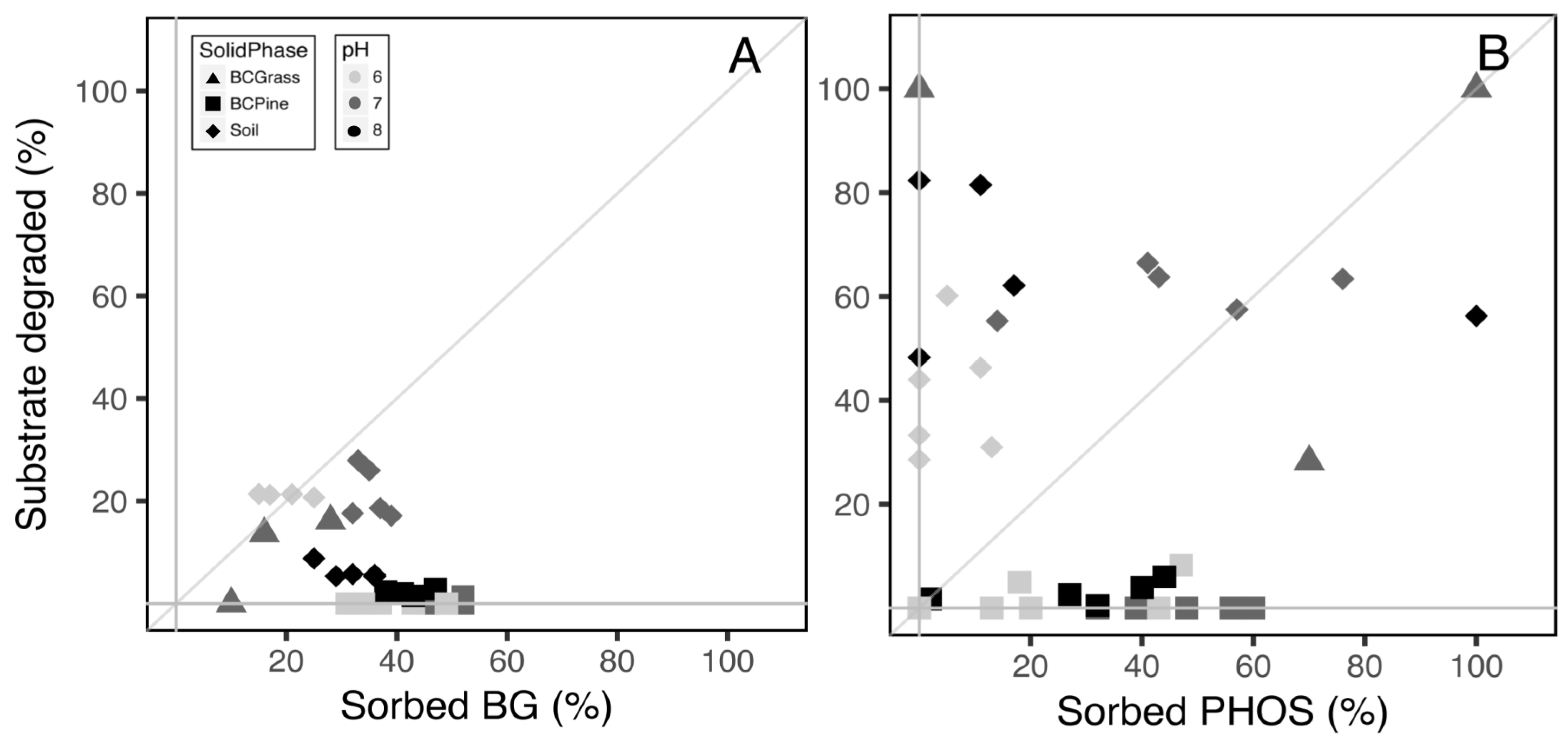
3.3. Assay of Substrate Stability and Sorption
4. Discussion
4.1. Enzyme Sorption
4.2. Enzyme Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Lehmann, J. Bio-energy in the black. Front. Ecol. Environ. 2007, 5, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, S.; Verheijen, F.G.A.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Enders, A.; Hanley, K.; Whitman, T.; Joseph, S.; Lehmann, J. Characterization of biochars to evaluate recalcitrance and agronomic performance. Bioresour. Technol. 2012, 114, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: An Introduction. Sci. Technol. 2009, 1, 1–12. [Google Scholar]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil Enzyme Activities as Biological Indicators of Soil Health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.W.; Fricks, B.E.; Rocca, J.D.; Steinweg, J.M.; McMahon, S.K.; Wallenstein, M.D. High-throughput fluorometric measurement of potential soil extracellular enzyme activities. J. Vis. Exp. 2013, 81, e50961. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D. Cheaters, diffusion and nutrients constrain decomposition by microbial enzymes in spatially structured environments. Ecol. Lett. 2005, 8, 626–635. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia (Jena) 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Makoi, J.H.J.R.; Ndakidemi, P.A. Selected soil enzymes: Examples of their potential roles in the ecosystem. Afr. J. Biotechnol. 2008, 7, 181–191. [Google Scholar]
- Alster, C.; Koyama, A.; Johnson, N.G.; Wallenstein, M.D.; von Fischer, J.C. Temperature sensitivity of soil microbial communities: An application of macromolecular rate theory to microbial respiration. J. Geophys. Res. Biogeosci. 2016, 121, 1420–1433. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz-Ferreiro, J.; Gascó, G.; Gutiérrez, B.; Méndez, A. Soil biochemical activities and the geometric mean of enzyme activities after application of sewage sludge and sewage sludge biochar to soil. Biol. Fertil. Soil. 2012, 48, 511–517. [Google Scholar] [CrossRef]
- Bailey, V.L.; Fansler, S.J.; Smith, J.L.; Bolton, H. Reconciling apparent variability in effects of biochar amendment on soil enzyme activities by assay optimization. Soil Biol. Biochem. 2011, 43, 296–301. [Google Scholar] [CrossRef]
- Elzobair, K.A.; Stromberger, M.E.; Ippolito, J.A. Stabilizing effect of biochar on soil extracellular enzymes after a denaturing stress. Chemosphere 2016, 142, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, E.J.; Hansen, N.; Wallenstein, M.; Cotrufo, M.F. Biochar and manure amendments impact soil nutrients and microbial enzymatic activities in a semi-arid irrigated maize cropping system. Agric. Ecosyst. Environ. 2016, 233, 404–414. [Google Scholar] [CrossRef]
- Burns, R.G. Enzyme activity in soil: Location and a possible role in microbial ecology. Soil Biol. Biochem. 1982, 14, 423–427. [Google Scholar] [CrossRef]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment: Activity, Ecology, and Applications; Marcel Dekker: New York, NY, USA, 2002. [Google Scholar]
- Quiquampoix, H.; Servagent-noinville, S. Enzyme adsorption on soil mineral surfaces and consequences for catalytic activity. In Enzymes in the Environment: Activity, Ecology, and Applications; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002. [Google Scholar]
- Sandhya, C.; Nampoothiri, K.M.; Pandey, A. Microbial Enzymes and Biotransformations; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Sarkar, J.M.; Leonowicz, A.; Bollag, J.M. Immobilization of enzymes on clays and soils. Soil Biol. Biochem. 1989, 21, 223–230. [Google Scholar] [CrossRef]
- Datta, R.; Anand, S.; Moulick, A.; Baraniya, D.; Pathan, S.I.; Rejsek, K.; Vranova, V.; Sharma, M.; Sharma, D.; Kelkar, A.; et al. How enzymes are adsorbed on soil solid phase and factors limiting its activity: A review. Int. Agrophys. 2017, 31, 287–302. [Google Scholar] [CrossRef]
- Quiquampoix, H.; Burns, R.G. Interactions between proteins and soil mineral surfaces: Environmental and health consequences. Elements 2007, 3, 401–406. [Google Scholar] [CrossRef]
- Wu, F.; Jia, Z.; Wang, S.; Chang, S.X.; Startsev, A. Contrasting effects of wheat straw and its biochar on greenhouse gas emissions and enzyme activities in a Chernozemic soil. Biol. Fertil. Soil. 2013, 49, 555–565. [Google Scholar] [CrossRef]
- Lammirato, C.; Miltner, A.; Kaestner, M. Effects of wood char and activated carbon on the hydrolysis of cellobiose by β-glucosidase from Aspergillus niger. Soil Biol. Biochem. 2011, 43, 1936–1942. [Google Scholar] [CrossRef]
- Swaine, M.; Obrike, R.; Clark, J.M.; Shaw, L.J. Biochar alteration of the sorption of substrates and products in soil enzyme assays. Appl. Environ. Soil Sci. 2013. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Liu, D.; Mentler, A.; Zehetner, F.; Zechmeister-Boltenstern, S. Biochar application reduces protein sorption in soil. Org. Geochem. 2015, 87, 21–24. [Google Scholar] [CrossRef]
- Lammirato, C.; Miltner, A.; Wick, L.Y.; Kästner, M. Hydrolysis of cellobiose by -glucosidase in the presence of soil minerals—Interactions at solid-liquid interfaces and effects on enzyme activity levels. Soil Biol. Biochem. 2010, 42, 2203–2210. [Google Scholar] [CrossRef]
- Watanabe, T.; Sato, T.; Yoshioka, S.; Koshijima, T.; Kuwahara, M. Purification and properties of Aspergillus niger beta-glucosidase. Eur. J. Biochem. 1992, 209, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Durmus, A.; Eicken, C.; Sift, B.H.; Kratel, A.; Kappl, R.; Hüttermann, J.; Krebs, B. The active site of purple acid phosphatase from sweet potatoes (Ipomoea batatas): Metal content and spectroscopic characterization. Eur. J. Biochem. 1999, 260, 709–716. [Google Scholar] [CrossRef] [PubMed]
- National Cooperative Soil Survey. Fort Collins Series. Available online: https://soilseries.sc.egov.usda.gov (accessed on 1 October 2018).
- Abulobaida, M. The Effect of Irrigaiton and Cropping Systems on Soil Carbon and Nitrogen Stocks and Organic Matter Aggregation in Semi-Arid Lands. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2014. [Google Scholar]
- Vogelmann, E.S.; Prevedello, J.; Awe, G.O.; Reinert, D.J. Soil hydrophobicity: Comparative study of usual determination methods. Cienc. Rural 2015, 45, 260–266. [Google Scholar] [CrossRef]
- Asadi, A.; Huat, B.B.K.; Hanafi, M.M.; Mohamed, T.A.; Shariatmadari, N. Role of organic matter on electroosmotic properties and ionic modification of organic soils. Geosci. J. 2009, 13, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Ogle, D.H. FSA: Fisheries Stock Analysis, R Package Version 0.8.20. Available online: https://cran.r-project.org (accessed on 1 October 2018).
- Baron, M.H.; Revault, M.; Servagent-Noinville, S.; Abadie, J.; Quiquampoix, H. Chymotrypsin Adsorption on Montmorillonite: Enzymatic Activity and Kinetic FTIR Structural Analysis. J. Colloid Interface Sci. 1999, 214, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Oleszczuk, P.; Jośko, I.; Futa, B.; Pasieczna-Patkowska, S.; Pałys, E.; Kraska, P. Effect of pesticides on microorganisms, enzymatic activity and plant in biochar-amended soil. Geoderma 2014, 214, 10–18. [Google Scholar] [CrossRef]
- Du, Z.; Wang, Y.; Huang, J.; Lu, N.; Liu, X.; Lou, Y.; Zhang, Q. Consecutive biochar application alters soil enzyme activities in the winter wheat—Growing season. Soil Sci. 2014, 179, 75–83. [Google Scholar] [CrossRef]
- Wang, X.; Song, D.; Liang, G.; Zhang, Q.; Ai, C.; Zhou, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Shindo, H.; Watanabe, D.; Onaga, T.; Urakawa, M.; Nakahara, O.; Huang, Q. Adsorption, activity, and kinetics of acid phosphatase as influenced by selected oxides and clay minerals. Soil Sci. Plant. Nutr. 2002, 48, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Sopeña, F.; Bending, G.D. Impacts of biochar on bioavailability of the fungicide azoxystrobin: A comparison of the effect on biodegradation rate and toxicity to the fungal community. Chemosphere 2013, 91, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Leprince, F.; Quiquampoix, H. Extracellular enzyme activity in soil: Effect of pH and ionic strength on the interaction with montmorillonite of two acid phosphatases secreted by the ectomycorrhizal fungus Hebeloma cylindrosporum. Eur. J. Soil Sci. 1996, 47, 511–522. [Google Scholar] [CrossRef]
- Zimmerman, A.R.; Gao, B.; Ahn, M.Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | Abbrev. | Source | Expected Activity (U/mg) c | Optimal pH | Isoelectric Point pH | Atomic Weight (kDa) |
|---|---|---|---|---|---|---|
| β-Glucosidase | BG | Aspergillus niger | ≥0.75 | 4.0 | 4 a | 240 a |
| Acid Phosphatase | PHOS | Sweet potato | 0.5–3.0 | 4.8 | 5.2 b | 110–112 b |
| Solid Phase | Description | pHH20 | BET Surface Area (m2 g−1) | BET Average Pore Width (nm) | Isoelectric Point (pH) | Hydrophobicity (seconds) |
|---|---|---|---|---|---|---|
| Soil | Irrigated sandy clay loam | 8.7 | 27.5 | 6.16 | <1 | Low (7.7) |
| Pinewood Biochar | Max pyrolysis temperature: 400–700 °C a | 9.2 | 232.7 | 4.51 | 1–2 | Medium (22) |
| Grass Biochar | Max pyrolysis temperature: 300 °C | 9.9 | 6.3 | 19.12 | 1 | High (>300) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foster, E.J.; Fogle, E.J.; Cotrufo, M.F. Sorption to Biochar Impacts β-Glucosidase and Phosphatase Enzyme Activities. Agriculture 2018, 8, 158. https://doi.org/10.3390/agriculture8100158
Foster EJ, Fogle EJ, Cotrufo MF. Sorption to Biochar Impacts β-Glucosidase and Phosphatase Enzyme Activities. Agriculture. 2018; 8(10):158. https://doi.org/10.3390/agriculture8100158
Chicago/Turabian StyleFoster, Erika J., Emily J. Fogle, and M. Francesca Cotrufo. 2018. "Sorption to Biochar Impacts β-Glucosidase and Phosphatase Enzyme Activities" Agriculture 8, no. 10: 158. https://doi.org/10.3390/agriculture8100158
APA StyleFoster, E. J., Fogle, E. J., & Cotrufo, M. F. (2018). Sorption to Biochar Impacts β-Glucosidase and Phosphatase Enzyme Activities. Agriculture, 8(10), 158. https://doi.org/10.3390/agriculture8100158