Limits to the Biofortification of Leafy Brassicas with Zinc

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Growth Conditions
2.3. Plant Analysis
2.4. Statistics
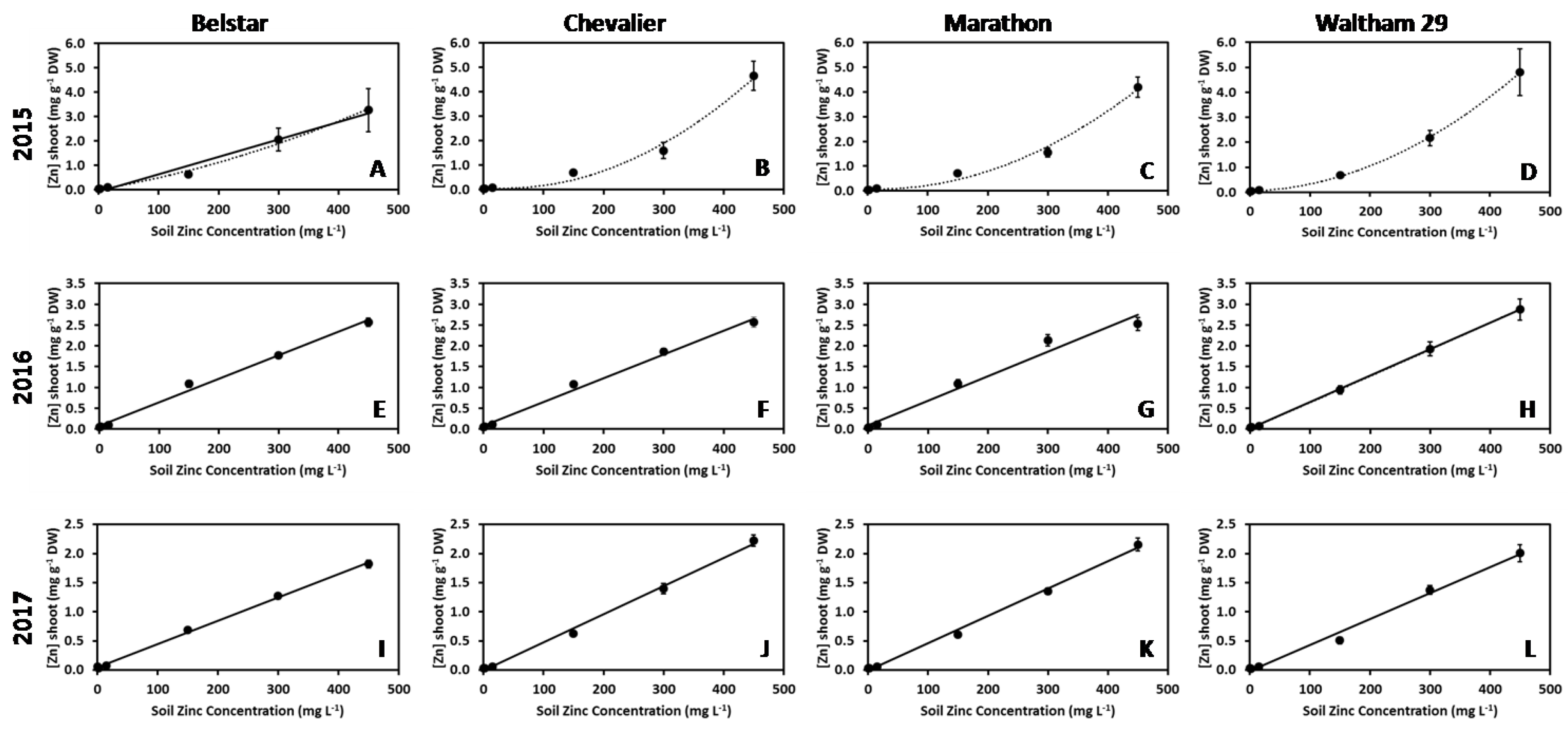
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.R.; Singh, G.M.; Brown, K.H. Estimating the global prevalence of inadequate zinc intake from national food balance sheets: Effects of methodological assumptions. PLoS ONE 2012, 7, e50565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumssa, D.B.; Joy, E.J.M.; Ander, E.L.; Watts, M.J.; Young, S.D.; Walker, S.; Broadley, M.R. Dietary calcium and zinc deficiency risks are decreasing but remain prevalent. Sci. Rep. 2015, 5, 10974. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Biofortification of edible crops. In eLS; John Wiley & Sons: Chichester, UK, 2016. [Google Scholar] [CrossRef]
- Cakmak, I.; McLaughlin, M.J.; White, P. Zinc for better crop production and human health. Plant Soil 2017, 411, 1–4. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Khoshgoftarmanesh, A.H.; Schulin, R.; Chaney, R.L.; Daneshbakhsh, B.; Afyuni, M. Micronutrient-efficient genotypes for crop yield and nutritional quality in sustainable agriculture. A review. Agron. Sustain. Dev. 2010, 30, 83–107. [Google Scholar] [CrossRef]
- Joy, E.J.M.; Ander, E.L.; Young, S.D.; Black, C.R.; Watts, M.J.; Chilimba, A.D.C.; Chilimba, B.; Siyame, E.W.P.; Kalimbira, A.A.; Hurst, R.; et al. Dietary mineral supplies in Africa. Physiol. Plant. 2014, 151, 208–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velu, G.; Ortiz-Monasterio, I.; Cakmak, I.; Hao, Y.; Singh, R.P. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 2014, 59, 365–372. [Google Scholar] [CrossRef]
- Gartler, J.; Robinson, B.; Burton, K.; Clucas, L. Carbonaceous soil amendments to biofortify crop plants with zinc. Sci. Total Environ. 2013, 465, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Šlosár, M.; Mezeyová, I.; Hegedüsová, A.; Andrejiová, A.; Kováčik, P.; Lošák, T.; Kopta, T.; Keutgen, A.J. Effect of zinc fertilisation on yield and selected qualitative parameters of broccoli. Plant Soil Environ. 2017, 63, 282–287. [Google Scholar] [CrossRef]
- Pfeiffer, W.H.; McClafferty, B. HarvestPlus: Breeding crops for better nutrition. Crop Sci. 2007, 47, S88–S105. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed]
- Sayre, R.; Beeching, J.R.; Cahoon, E.B.; Egesi, C.; Fauquet, C.; Fellman, J.; Fregene, M.; Gruissem, W.; Mallowa, S.; Manary, M.; et al. The BioCassava Plus Program: Biofortification of cassava for Sub-Saharan Africa. Ann. Rev. Plant Biol. 2011, 62, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Phattarakul, N.; Rerkasem, B.; Li, L.J.; Wu, L.H.; Zou, C.Q.; Ram, H.; Sohu, V.S.; Kang, B.S.; Surek, H.; Kalayci, M.; et al. Biofortification of rice grain with zinc through zinc fertilization in different countries. Plant Soil 2012, 361, 131–141. [Google Scholar] [CrossRef]
- Zou, C.Q.; Zhang, Y.Q.; Rashid, A.; Ram, H.; Savasli, E.; Arisoy, R.Z.; Ortiz-Monasterio, I.; Simunji, S.; Wang, Z.H.; Sohu, V.; et al. Biofortification of wheat with zinc through zinc fertilization in seven countries. Plant Soil 2012, 361, 119–130. [Google Scholar] [CrossRef]
- Saltzman, A.; Birol, E.; Bouis, H.E.; Boy, E.; De Moura, F.F.; Islam, Y.; Pfeiffer, W.H. Biofortification: Progress toward a more nourishing future. Glob. Food Secur. 2013, 2, 9–17. [Google Scholar] [CrossRef]
- Pucher, A.; Høgh-Jensen, H.; Gondah, J.; Hash, C.T.; Haussmann, B.I.G. Micronutrient density and stability in West African pearl millet—Potential for biofortification. Crop Sci. 2014, 54, 1709–1720. [Google Scholar] [CrossRef]
- Mallikarjuna, M.G.; Thirunavukkarasu, N.; Hossain, F.; Bhat, J.S.; Jha, S.K.; Rathore, A.; Agrawal, P.K.; Pattanayak, A.; Reddy, S.S.; Gularia, S.K.; et al. Stability performance of inductively coupled plasma mass spectrometry-phenotyped kernel minerals concentration and grain yield in maize in different agro-climatic zones. PLoS ONE 2015, 10, e0139067. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-P.; Zhang, Y.-Q.; Tong, Y.-P.; Xue, Y.-F.; Liu, D.Y.; Zhang, W.; Deng, Y.; Meng, Q.-F.; Yue, S.-C.; Yan, P.; et al. Harvesting more grain zinc of wheat for human health. Sci. Rep. 2017, 7, 7016. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Thompson, J.A.; Wright, G.; Rasmussen, S.K. Biofortifying Scottish potatoes with zinc. Plant Soil 2017, 411, 151–165. [Google Scholar] [CrossRef]
- Thomson, C.A.; Newton, T.R.; Graver, E.J.; Jackson, K.A.; Reid, P.M.; Hartz, V.L.; Cussler, E.C.; Hakim, I.A. Cruciferous vegetable intake questionnaire improves cruciferous vegetable intake estimates. J. Am. Diet. Assoc. 2007, 107, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 25 October 2017).
- Moreno, D.A.; Carvajal, M.; López-Berenguer, C.; Garcia-Viguera, C. Chemical and biological characterization of nutraceutical compounds of broccoli. J. Pharm. Biomed. Anal. 2006, 41, 1508–1522. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrijver, R.; Hansen, M.; Gerhauser, C.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219. [Google Scholar] [CrossRef] [PubMed]
- Harrison, H.C.; Bergman, E.L. Calcium, magnesium, and potassium interrelationships affecting cabbage production. J. Am. Soc. Hortic. Sci. 1981, 106, 500–503. [Google Scholar]
- Kopsell, D.E.; Kopsell, D.A.; Lefsrud, M.G.; Curran-Celentano, J. Variability in elemental accumulations among leafy Brassica oleracea cultivars and selections. J. Plant Nutr. 2004, 27, 1813–1826. [Google Scholar] [CrossRef]
- Broadley, M.R.; Lochlainn, S.; Hammond, J.P.; Bowen, H.C.; Cakmak, I.; Eker, S.; Erdem, H.; King, G.J.; White, P.J. Shoot zinc (Zn) concentration varies widely within Brassica oleracea L. and is affected by soil Zn and phosphorus (P) levels. J. Hortic. Sci. Biotechnol. 2010, 85, 375–380. [Google Scholar] [CrossRef]
- Singh, B.K.; Sharma, S.R.; Singh, B. Variation in mineral concentrations among cultivars and germplasms of cabbage. J. Plant Nutr. 2010, 33, 95–104. [Google Scholar] [CrossRef]
- Kałużewicz, A.; Bosiacki, M.; Frąszczak, B. Mineral composition and the content of phenolic compounds of ten broccoli cultivars. J. Elementol. 2016, 21, 53–65. [Google Scholar] [CrossRef]
- Šindelářová, K.; Száková, J.; Tremlová, J.; Mestek, O.; Praus, L.; Kaňa, A.; Najmanová, J.; Tlustoš, P. The response of broccoli (Brassica oleracea convar. italica) varieties on foliar application of selenium: Uptake, translocation, and speciation. Food Addit. Contam. A 2015, 32, 2027–2038. [Google Scholar] [CrossRef]
- Xiao, Z.L.; Codling, E.E.; Luo, Y.G.; Nou, X.W.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Comp. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef]
- Lewu, F.B.; Lewu, M.N.; Mavengahama, S. Influence of genotype and harvesting age on the mineral dynamics of Brassica oleracea var. acephala cultivated in South Africa. J. Food Agric. Environ. 2012, 10, 563–566. [Google Scholar]
- Singh, B.K.; Sharma, S.R.; Singh, B. Genetic combining ability for concentration of mineral elements in cabbage head (Brassica oleracea var. capitata L.). Euphytica 2012, 184, 265–273. [Google Scholar] [CrossRef]
- MacNicol, R.D.; Beckett, P.H.T. Critical tissue concentrations of potentially toxic elements. Plant Soil 1985, 85, 107–109. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R.; Thompson, J.A.; McNicol, J.W.; Crawley, M.J.; Poulton, P.R.; Johnston, A.E. Testing the distinctness of shoot ionomes of angiosperm families using the Rothamsted Park Grass Continuous Hay Experiment. New Phytol. 2012, 196, 101–109. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R.; Gregory, P.J. Managing the nutrition of plants and people. Appl. Environ. Soil Sci. 2012, 2012, 104826. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Xian, X. Effect of chemical forms of cadmium, zinc, and lead in polluted soils on their uptake by cabbage plants. Plant Soil 1989, 113, 257–264. [Google Scholar] [CrossRef]
- Handreck, K.A. Total and extractable copper and zinc as assessors of phytotoxicity in soilless potting media. Commun. Soil Sci. Plant Anal. 1994, 25, 2313–2340. [Google Scholar] [CrossRef]
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Barrameda-Medina, Y.; Montesinos-Pereira, D.; Romero, L.; Ruiz, J.M.; Blasco, B. Comparative study of the toxic effect of Zn in Lactuca sativa and Brassica oleracea plants: I. Growth, distribution, and accumulation of Zn, and metabolism of carboxylates. Environ. Exp. Bot. 2014, 107, 98–104. [Google Scholar] [CrossRef]
- Paradisone, V.; Barrameda-Medina, Y.; Montesinos-Pereira, D.; Romero, L.; Esposito, S.; Ruiz, J.M. Roles of some nitrogenous compounds protectors in the resistance to zinc toxicity in Lactuca sativa cv. Phillipus and Brassica oleracea cv. Bronco. Acta Physiol. Plant. 2015, 37, 137. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Baczek-Kwinta, R.; Bartoszek, A.; Piekarska, A.; Huk, A.; Manikowska, A.; Antonkiewicz, J.; Namiesnik, J.; Konieczka, P. The dose-dependent influence of zinc and cadmium contamination of soil on their uptake and glucosinolate content in white cabbage (Brassica oleracea var. capitata f. alba). Environ. Toxicol. Chem. 2012, 31, 2482–2489. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.G.; Grewal, H.S.; Graham, R.D. Dry matter production and uptake of zinc and phosphorus in two oilseed rape genotypes under differential rates of zinc and phosphorus supply. J. Plant Nutr. 1998, 21, 25–38. [Google Scholar] [CrossRef]
- Bernhard, R.; Verkleij, J.A.C.; Nelissen, H.J.M.; Vink, J.P.M. Plant-specific responses to zinc contamination in a semi-field lysimeter and on hydroponics. Environ. Pollut. 2005, 138, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Allica, J.; Becerril, J.M.; Garbisu, C. Assessment of the phytoextraction potential of high biomass crop plants. Environ. Pollut. 2008, 152, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, S.H.; Wang, P.F.; Hou, J.; Zhang, W.J.; Li, W.; Lin, Z.P. The effect of excess Zn on mineral nutrition and antioxidative response in rapeseed seedlings. Chemosphere 2009, 75, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Peško, M.; Molnárová, M.; Fargašová, A. Effect of lead and zinc treatments on Brassica napus L. (cv. Verona) plants: Accumulation and physio-biochemical changes. Fresenius Environ. Bull. 2015, 24, 3213–3219. [Google Scholar]
- Ebbs, S.D.; Kochian, L.V. Toxicity of zinc and copper to Brassica species: Implications for phytoremediation. J. Environ. Qual. 1997, 26, 776–781. [Google Scholar] [CrossRef]
- Hamlin, R.L.; Schatz, C.; Barker, A.V. Zinc accumulation in Indian mustard as influenced by nitrogen and phosphorus nutrition. J. Plant Nutr. 2003, 26, 177–190. [Google Scholar] [CrossRef]
- Hamlin, R.L.; Barker, A.V. Influence of ammonium and nitrate nutrition on plant growth and zinc accumulation by Indian mustard. J. Plant Nutr. 2006, 29, 1523–1541. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Genotypes | Maximum Shoot Biomass | Critical Zn Concentration | ||
|---|---|---|---|---|---|
| Substrate Zn Regression (g DW) | Shoot Zn Regression (g DW) | Substrate (mg L−1) | Shoot (mg g−1 DW) | ||
| Cabbage | |||||
| 2014 | Bison | 1.07 | 1.06 | 96 | 0.789 |
| 2014 | Cape Horn | 2.08 | 2.06 | 100 | 0.802 |
| 2014 | Tundra | 1.52 | 1.52 | 260 | 1.201 |
| 2014 | Red Drumhead | 1.48 | 1.45 | 5 | 0.074 |
| Broccoli | |||||
| 2015 | Belstar | 11.33 | 11.33 | 107 | 0.434 |
| 2015 | Chevalier | 10.72 | 10.72 | 105 | 0.499 |
| 2015 | Marathon | 11.87 | 11.87 | 121 | 0.514 |
| 2015 | Waltham 29 | 9.40 | 9.64 | 121 | 0.117 |
| 2016 | Belstar | 3.12 | 3.12 | 139 | 1.018 |
| 2016 | Chevalier | 3.68 | 3.68 | 120 | 0.901 |
| 2016 | Marathon | 3.54 | 3.55 | 43 | 0.277 |
| 2016 | Waltham 29 | 2.91 | 2.91 | 68 | 0.406 |
| 2017 | Belstar | 5.51 | 5.51 | 408 | 1.666 |
| 2017 | Chevalier | 7.03 | 7.03 | 308 | 1..424 |
| 2017 | Marathon | 6.68 | 6.70 | 332 | 1.527 |
| 2017 | Waltham 29 | 6.18 | 6.24 | 271 | 1.195 |
| Year | Gradient (mg Zn kg−1 DW/mg Zn L−1), Intercept (mg Zn kg−1 DW) | |||
|---|---|---|---|---|
| Bison | Cape Horn | Tundra | Red Drumhead | |
| 2014 | 5.802, 73.64 | 5.209, 85.92 (A) | 4.620, 4.706 | 5.641, 83.76 (A) |
| Belstar | Chevalier | Marathon | Waltham 29 | |
| 2015 | 7.126, 70.16 | 5.170, 5.644 (E) | 5.015, 12.83 (E) | 6.853, 36.36 (E) |
| 2016 | 5.654, 74.84 | 5.714, 80.16 | 5.894, 88.42 | 6.350, 12.32 |
| 2017 | 3.997, 49.46 | 4.837, 8.768 | 4.680, 7.523 | 4.452, 11.53 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, P.J.; Pongrac, P.; Sneddon, C.C.; Thompson, J.A.; Wright, G. Limits to the Biofortification of Leafy Brassicas with Zinc. Agriculture 2018, 8, 32. https://doi.org/10.3390/agriculture8030032
White PJ, Pongrac P, Sneddon CC, Thompson JA, Wright G. Limits to the Biofortification of Leafy Brassicas with Zinc. Agriculture. 2018; 8(3):32. https://doi.org/10.3390/agriculture8030032
Chicago/Turabian StyleWhite, Philip J., Paula Pongrac, Claire C. Sneddon, Jacqueline A. Thompson, and Gladys Wright. 2018. "Limits to the Biofortification of Leafy Brassicas with Zinc" Agriculture 8, no. 3: 32. https://doi.org/10.3390/agriculture8030032
APA StyleWhite, P. J., Pongrac, P., Sneddon, C. C., Thompson, J. A., & Wright, G. (2018). Limits to the Biofortification of Leafy Brassicas with Zinc. Agriculture, 8(3), 32. https://doi.org/10.3390/agriculture8030032