


Marine Archaeal Extracellular Polymeric Substances from Halococcus AMS12, Their Characterization, and Biological Properties


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of Marine Archaea
2.3. Optimal Growth Parameters of Archaea
2.4. Production and Purification of EPS
2.5. Characterization of Marine Archaeal EPS
2.6. Biosurfactant and Cell Membrane Protection Assay
2.7. Enzyme Activity of Marine Archaeal EPS
2.8. Antioxidant Activity of EPS
2.9. Cell Viability (MTT) and DCFH-DA Assay
2.10. Dichloro-Dihydro-Fluorescein Diacetate DCFH-DA) Assay
3. Results
3.1. Isolation and Identification of Marine Archaea
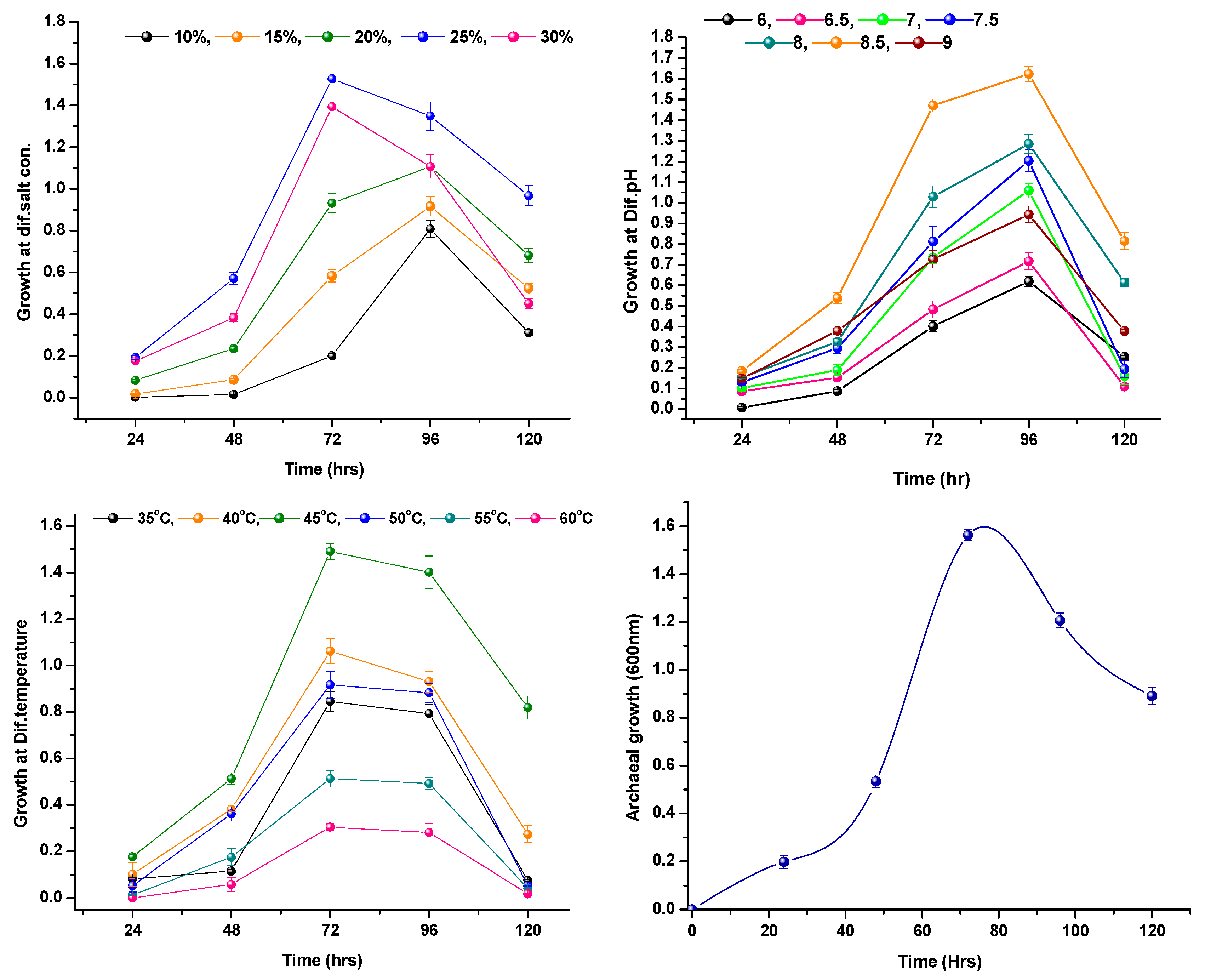
3.2. Optimal Growth Condition of Halococcus sp. AMS12
3.3. Production and Characterization of Marine Archaeal EPS
3.4. Biosurfactant Activity and Cell Membrane Protection
3.5. Enzymatic Activity of Marine Halococcus sp. AMS12
3.6. Antioxidant Activity of EPS Obtained from Halococcus sp. AMS12
3.7. Cell Viability (MTT) and DCFH-DA Assay
4. Discussion
4.1. Isolation and Identification of Marine Archaea
4.2. Optimal Growth Condition of Halococcus sp. AMS12
4.3. Production and Characterization of Marine Archaeal EPS
4.4. Biosurfactant Activity and Cell Membrane Protection
4.5. Enzymatic Activity of Marine Halococcus sp. AMS12
4.6. Antioxidant Activity of EPS Obtained from Halococcus sp. AMS12
4.7. Cell Viability (MTT) and DCFH-DA Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bullock, C. The Archaea-a biochemical perspective. Biochem. Mol. Biol. Educ. 2000, 28, 186–191. [Google Scholar]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adair, K.; Blazewicz, S.J.; Hungate, B.A.; Hart, S.C.; Dijkstra, P.; Schwartz, E. A positive relationship between the abundance of ammonia oxidizing archaea and natural abundance δ15N of ecosystems. Soil Biol. Biochem. 2013, 65, 313–315. [Google Scholar] [CrossRef]
- Kumar, S.; Karan, R.; Kapoor, S.; Singh, S.P.; Khare, S.K. Screening and isolation of halophilic bacteria producing industrially important enzymes. Braz. J. Microbiol. 2012, 43, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuneda, S.; Aikawa, H.; Hayashi, H.; Yuasa, A.; Hirata, A. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface. FEMS Microbiol. Lett. 2003, 223, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Kazak, H.; Oner, E.T.; Dekker, R.F.H. Extremophiles as sources of exopolysaccharides. In Hand book of Carbohydrate Polymers: Development, Properties and Applications; Ito, R., Matsuo, Y., Eds.; Nova Science Publishers: New York, NY, USA, 2010; pp. 605–619. [Google Scholar]
- Ruffing, A.; Chen, R.R. Metabolic engineering of microbes for oligosaccharide and polysaccharide synthesis. Microb. Cell Factories 2006, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Y.; Wang, J.L.; Pan, X.L. Cadmium sorption by EPSs produced by anaerobic sludge under sulfate-reducing conditions. J. Hazard. Mater. 2006, 138, 589–593. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Rusch, A.; Gaidos, E. Nitrogen cycling bacteria and archaea in the carbon sediment of a coral reef. Geobiology 2013, 11, 472–484. [Google Scholar] [CrossRef]
- Nicolaus, B.; Kambourova, M.; Oner, E.T. Exopolysaccharides from extremophiles: From fundamentals to biotechnology. Environ. Technol. 2010, 31, 1145–1158. [Google Scholar] [CrossRef]
- Poli, A.; Kazak, H.; Gurleyendag, B. High level synthesis of levan by a novel Halomonas species growing on defined media. Carbohydr. Polym. 2009, 78, 651–657. [Google Scholar] [CrossRef]
- Akolkar, A.V.; Deepa, D.; Desai, A.J. Acceleration of fish sauce fermentation by Halobacterium sp. SP1 (1). J. Appl. Microbiol. 2009, 109, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Hamid, M.; Abedi, D.; Mohmoudpour, H.R.; Akbari, V. Comparison of five methods for extraction of genomic DNA from a marine archaea Pyrococcus furiosus. Pak. J. Med. Sci. 2013, 29, 390–394. [Google Scholar]
- Delong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, C.; Patel, P.; Markande, A.R.; Kamala, K.; Sivaperumal, P. Exploration of haloarchaea for their potential application in food industry. Int. J. Environ. Sci. Technol. 2020, 17, 4455–4464. [Google Scholar] [CrossRef]
- Forster, C.F.; Clarke, A.R. The production of polymer from activated sludge by ethanolic extraction and its relation to treatment plant operation. Water Pollut. Control (Maidstone) 1983, 82, 430–433. [Google Scholar]
- Li, X.Y.; Yang, S.F. Influence of loosely bound extracellular polymeric substances (EPS) on the flocculation, sedimentation and dewaterability of activated sludge. Water Res. 2007, 41, 1022–1030. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Kamala, K.; Rajaram, R. Adsorption of Cesium ion by marine actinobacterium Nocardiopsis sp. 13H and their Extracellular Polymeric Substances (EPS) role in Bioremediation. Environ. Sci. Pollut. Res. 2018, 25, 4254–4267. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350. [Google Scholar] [CrossRef]
- Burton, K. A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem. J. 1956, 62, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.C.; Santschi, P.H. Spectrophotometric determination of total uronic acids in seawater using cation exchange separation and pre-concentration by lyophilization. Anal. Chim. Acta 2001, 427, 111–117. [Google Scholar] [CrossRef]
- Parekh, A.L.; Jung, D.H. Cholesterol determination with ferric acetate—Uranium. Anal. Chem. 1970, 42, 1423. [Google Scholar] [CrossRef]
- Kamala, K.; Sivaperumal, P.; Thilagaraj, R.; Natarajan, E. Bioremediation of Sr2+ ion radionuclide by using marine Streptomyces sp. CuOff24 Extracellular Polymeric Substances (EPS). J. Chem. Technol. Biotechnol. 2019, 95, 893–903. [Google Scholar]
- Sajjad, W.; Qadir, S.; Ahmad, M.; Rafiq, M.; Hasan, F.; Tehan, R. Ectoine: A compatible solute in radio−halophilic Stenotrophomonas sp. WMA−LM 19 strain to prevent ultraviolet−induced protein damage. J. Appl. Microbiol. 2018, 125, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Kamala, K.; Sivaperumal, P.; Gobalakrishnan, R.; Swarnakumar, N.S.; Rajaram, R. Isolation and characterization of biologically active alkaloids from marine actinobacteria Nocardiopsis sp. NCS1. Biocatal. Agric. Biotechnol. 2015, 4, 63–69. [Google Scholar] [CrossRef]
- Sivaperumal, P.; Kamala, K.; Rajaram, R.; Mishra, S.S. Melanin from marine Streptomyces (MVCS13) with potential effect against ornamental fish pathogens of Carassius auratus (Linnaeus, 1758). Biocatal. Agric. Biotechnol. 2014, 3, 134–141. [Google Scholar] [CrossRef]
- Bhavna, V.S.C.; Chopra, B.S.; Raut, S.; Sagar, A.; Asish, M.D.B.; Kharti, N. Antioxidant and wound healing property of gelsolin in 3T3-L1 Cells. Oxidative Med. Cell. Longev. 2020, 2020, 4045365. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 12, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Munson, M.A.; Nedwell, D.B.; Embley, T.M. Phylogentic diversity of archaea in sediment samples from coastal salt marsh. Appl. Environ. Microbiol. 1997, 63, 4729–4733. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Paola, L. Marine polysaccharide in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar]
- Cao, H.; Auguet, J.C.; Gu, J.D. Global ecological pattern of ammonia oxidizing archaea. PLoS ONE 2013, 8, e52853. [Google Scholar] [CrossRef] [Green Version]
- Uma, G.; Mariavincent, M.B.; Vincent, S.G.P.; Selvaraj, J.N.; Thavasimuthu, C. Nature and bioprospecting of haloalkaliphilics: A review. World J. Microbiol. Biotechnol. 2020, 36, 66. [Google Scholar] [CrossRef] [PubMed]
- Asha, K.R.T.; Vinitha, D.A.J.; Kiran, S.G.; Manjusha, W.A.; Sukumaran, N.; Selvin, J. Isolation and cultivation of Halophilic archaea rom solar salterns located in peninsular coast of India. Int. J. Microbiol. 2005, 1, 1–4. [Google Scholar]
- Bertoldo, C.; Antranikian, G. Biotechnology of archaea: Biotechnology Marine biotechnology. In Encyclopedia of Life Support System; Doelle, H.W., MarinBerovic, S.R., Eds.; Technical University Hamburg: Hamburg, Germany, 2000; Volume IX, pp. 186–219. [Google Scholar]
- Harada, M.R.; Yoza, B.A.; Masutani, S.M.; Li, Q.X. Diversity of archaea communities within contaminated sane samples from Johnston atoll. Bioremed. J. 2013, 17, 182–189. [Google Scholar] [CrossRef]
- Ellis, D.G.; Bizzoco, R.W.; Kelley, S.T. Halophilicarchaea determined from geothermal vent aerosols. Environ. Microbiol. 2008, 10, 1582–1590. [Google Scholar] [CrossRef]
- Kendall, M.M.; Boone, D.R. Cultivation of methanogens from shallow marine sediments at Hydrate ridge, Oregon. Archaea 2006, 2, 31–38. [Google Scholar] [CrossRef]
- Kendall, M.M.; Wardlaw, G.D.; Tang, C.F.; Bonin, A.S.; Liu, Y.; Valentine, D.L. Diversity of archaea in marine sediments from Skan bay, Alaska, including cultivated methanogens and description of Methanogenium boonei sp. nov. Appl. Environ. Microbiol. 2007, 73, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Roling, W.F.M.; Couto de Brito, I.R.; Swannell, R.P.J.; Head, I.M. Response of archaeal communities in beach sediments to spilled oil and bioremediation. Appl. Environ. Microbiol. 2004, 70, 2614–2620. [Google Scholar] [CrossRef] [Green Version]
- Kakhki, A.M.; Amoozegar, M.A.; Khaledi, E.M. Diversity of haloarchaeal strains isolated from salt alke. Int. J. Environ. Sci. Technol. 2011, 8, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Patil, J.; Bajekal, S. Diversity of hydrolytic enzymes in haloalkaliphilic archaea isolated from Lonarlake. Int. J. Sci. Res. 2013, 2, 414–415. [Google Scholar]
- Namwong, S.; Tanaupawat, S. Characterization of lipase producing Haloarchula strain from thai fish sauce. Chiang Mai J. Sci. 2017, 44, 39–49. [Google Scholar]
- Bergey, E.; Stinson, M. Heparin-inhibitable basement membrane-binding protein of Streptococcus pyogenes. Infect. Immun. 1998, 56, 1715–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wight, T.N.; Kinsella, M.G.; Qwarnstrom, E. The role of proteglycans in cell adhesion, migration and proliferation. Curr. Opin. Cell Biol. 1992, 4, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, J.C.; Burns, B.P. Extremophilic adaptation and biotechnological applications in diverse environments. AIMS Microbiol. 2015, 2, 251–261. [Google Scholar] [CrossRef]
- Popescu, G.; Dumitru, L. Biosorption of some heavy metals from media with high salt concentrations by halophilic Archaea. Biotechnol. Biotechnol. Equip. 2009, 23, 791–795. [Google Scholar] [CrossRef]
- Poli, A.; Di Donato, P.; Nicolaus, G.R. Synthesis, production, and biotechnological applications of exopolysaccharides and polyhydroxyalkanoates by Archaea. Archaea 2011, 2011, 693253. [Google Scholar] [CrossRef] [Green Version]
- Gana, K.S.; Gana, M.L.; Khemii, S.; Naimi, F.F.; Bouanane, N.A.; Penninckx, M.; Hacene, H. Isolation and characterization of archaea able to produce biosurfactant. J. Ind. Microbiol. Biotechnol. 2009, 36, 727–738. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Timmis, K.N.; Wray, V.; Fredrickso, H.L. Characterization of new lipopeptide surfactant produced by thermotolerant and halotolerant Subsurface Bacillus licheniformis BAS 50. Appl. Environ. Microbiol. 1995, 61, 1706–1713. [Google Scholar] [CrossRef]
- Trebbau de Acevedo, G.; McInerney, M.J. Emulsifying activity in thermophilic and extremely thermophlic microorganisms. J. Ind. Microbiol. Biotechnol. 1996, 16, 1–7. [Google Scholar]
- Poremba, K.; Gunkel, W.; Lang, S.; Wagner, F. Toxicity testing of synthetic and biogenic surfactants on marine microorganisms. Environ. Toxicol. Water Qual. 1991, 6, 157–163. [Google Scholar] [CrossRef]
- Surekha, K.S.; Banat, I.M.; Dhakephalkar, P.K.; Banpurkar, A.G.; Chopade, B.A. Biosurfactants, Bioemulsifiers and exopolysaccharides from marine microorganisms. Biotechnol. Adv. 2010, 28, 436–450. [Google Scholar]
- Djeridi, I.; Militon, C.; Grossi, V.; Cuny, P. Evidence of surfactant production by the haloarchaeaon Haloferax sp. MSNC14 hydrocarbon containing media. Extremophiles 2013, 17, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, A.; Smyth, T.J.P.; Marchant, R.; Banat, I.M. Production and roles of biosurfactants and bioemulsifiers in accessing hydrophobic substrates. In Handbook of Hydrocarbon and Lipid Microbiology; Kenneth, N.T., Ed.; Springer: London, UK; Berlin/Heidelberg, Germany, 2010; Chapter 47; Volume 2 part 7, pp. 1501–1512. [Google Scholar]
- Watanabe, K.; Tabuchi, M.; Morinaga, Y.; Yoshinaga, F. Structural features and properties of bacterial cellulose produced in agitated culture. Cellulose 1998, 5, 187–200. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol. Lett. 2017, 364, fnx120. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine Bacillus circulans. J. Appl. Microbiol. 2008, 4, 675–684. [Google Scholar] [CrossRef]
- Shivani, S.; Saptashwa, D.; Narayanan, K.B.; Rajnish, K.N. Bacterial exopolysaccharides from biofilms: Role in antimicrobial resistance and treatments. J. Genet. Eng. Biotechnol. 2021, 19, 140. [Google Scholar]
- Zorgani, A.; Patron, K.; Desvaux, M. New insight in the structural features of haloadaptation in α-amylases from halophilicarchaea following homology modeling: Folded and stable confirmation maintained through low hydrophobicity and high negative charged surface. J. Comput.-Aided Mol. Des. 2014, 28, 721–734. [Google Scholar] [CrossRef]
- Kamala, K.; Sivaperumal, P.; Bilal, A.P.; Mohammad, K.S. Identification of haloarchaea during fermentation of Sardinella longiceps for being the starter culture to accelerate fish sauce production. Int. J. Food Sci. Technol. 2021, 56, 5717–5725. [Google Scholar] [CrossRef]
- Singh, A.; Singh, A.K. Isolation, characterization and exploring biotechnological potential of halophilicarchaea from salterns of western India. 3 Biotech 2018, 8, 45. [Google Scholar] [CrossRef]
- Beblovranesevic, K.; Galinski, E.; Rachel, R.; Huber, H.; Rettberg, P. Influence of osmotic stress on desiccation and irradiation tolerance of hyper thermophilic microorganisms. Arch. Microbiol. 2017, 199, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Trincone, A. Molecular fishing: Marine oligosaccharides. Front. Mar. Sci. 2014, 1, 26. [Google Scholar] [CrossRef]
- Badarinath, A.V.; Mallikarjuna, K.R.; Chetty, C.M.S.; Ramkanth, S.; Rajan, T.V.S.; Gnanaprakash, K. A review on In-vitro antioxidant methods: Comparisions, correlations and considerations. Int. J. PharmTech Res. 2010, 2, 1276–1285. [Google Scholar]
- Meir, S.; Kanner, J.; Akiri, B.; Hadas, S.P. Determination and involvement of aqueous reducing compounds in oxidative defense system of various senescing leaves. J. Agric. Food Chem. 1995, 43, 1813–1819. [Google Scholar] [CrossRef]
- Liu, F.; Ng, T.B. Antioxidative and free radical scavenging activities of selected medicinal herbs. Life Sci. 2000, 66, 725–735. [Google Scholar] [CrossRef]
- Qi, H.M.; Zhang, Q.B.; Zhao, T.T.; Chen, R.; Zhang, H.; Niu, X.; Li, Z. Antioxidant activity ofdifferent sulfate content derivatives of polysaccharide extracted from Ulvapertusa (Chlorophyta) in vitro. Int. J. Biol. Macromol. 2005, 37, 195–199. [Google Scholar] [CrossRef]
- Virginia, H.; Sarah, L.E.; Rachel, J.S.; Nathaniel, T.; Josseph, S.; Adam, E.; Cecilia, G. Mitochondrial nitric oxide synthase: Role in pathophysiology. IUBMB Life 2003, 55, 599–603. [Google Scholar]
- Williams, D.E.; Davies, J.; Patrick, B.O.; Bottriell, H.; Tarling, T.; Roberge, M.; Andersen, R.J. Caldoniamides A-G, Tryptophan- derived alkaloids produced in culture by Streptomyces uncialis. Org. Lett. 2008, 10, 3501–3504. [Google Scholar] [CrossRef]
- Kamala, K.; Karuppiah, V.; Sivakumar, K. Comparative evolution of in vitro antioxidant potent of the marine actinobacteria from Gulf of mannar biosphere reserve. Int. J. Pharm. Biol. Sci. 2013, 4, 207–216. [Google Scholar]
- Sai Ut, S.; Benjakul, S.; Sumpavapon, P. Screening gelatinolytic enzyme producing bacteria for production of hydrolysate with antioxidant activity. Int. Proc. Chem. Biol. Environ. Eng. 2013, 53, 55–59. [Google Scholar]
- Mao, G.D.; Thomas, P.D.; Lopaschuk, G.D.; Poznansky, M.J. Superoxide Dismutase (SOD)-Catalase Conjugates—Role of Hydrogen Peroxide and the Fenton Reaction in SOD Toxicity. J. Biol. Chem. 1993, 268, 416–420. [Google Scholar] [CrossRef]
- Schafer, G.; Stefan, A.; Ralf, M. Archaeal complex II: Classical and nonclassical succinate quinone reductase within unusual features. Biochim. Biophys. Acta 2002, 1553, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Wakagi, T.; Hiroshi, N.; Masayuki, M.; Shinya, F. Archaeal Mo-containing glyceraldehyde oxidoreductse isozymes exhibit diverse substrate specificities through unique subunit assemblies. PLoS ONE 2016, 11, e0147333. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, L.Z.; Qin, Y.L.; Liang, Z.L.; Li, X.T.; Yin, H.Q.; Liu, L.J.; Liu, S.J.; Jiang, C.Y. Comparative genomic analysis reveals the metabolism and evolution of the thermophilic archaeal genus Metallosphaera. Front. Microbiol. 2020, 11, 1192. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wang, H.; Zhang, Z.; Li, K.; Zhang, X.; Mora-Lopez, M.; Jiang, C.; Liu, C.; Wang, L.; Zhu, Y.; et al. Genome sequencing of Sulfolobus so A20 from costa Rica and comparative analyses of the putative pathways of carbon, nitrogen and sulfur metabolism in various Sulfolobus strains. Front. Microbiol. 2016, 7, 1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheaton, G.H.; Mukherjee, A.; Kelly, R.M. Transcriptomes of the extremely thermoacidophilic archaeon Metallosphaera sedula exposed to metal shock reveal generic and specific metal responses. Appl. Environ. Microbiol. 2016, 82, 4613–4627. [Google Scholar] [CrossRef] [PubMed]
- Mesle, M.; Beam, J.P.; Jay, Z.J.; Bodle, B.; Bogenschutz, E.L.; Inskeep, W.P. Hydrogen peroxide cycling in high-temperature acidic geothermal springs and potential implications for oxidative stress response. Front. Mar. Sci. 2017, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Safarpour, A.; Ebrahimi, M.; Fazeli, S.A.S.; Amoozegar, M.A. Supernatant metabolites from halophilicarchaea to reduce tumorgenesis in prostate cancer In-vitro and In-vivo. Iran. J. Pharm. Res. 2019, 18, 241–253. [Google Scholar]
- Hou, J.; Cui, H.L. In vitro antioxidant, antihemolytic and anticancer activity of the carotenoids from halophilicarchaea. Curr. Microbiol. 2018, 75, 266–271. [Google Scholar] [CrossRef]
- Leone, A.; Zefferino, R.; Longo, C.; Leo, L.; Zacheo, G. Supercritical CO2-extracted tomato oleoresins enhance gap junction intercellular communications and recover from mercury chloride inhibition in keratinocytes. J. Agric. Food Chem. 2010, 58, 4769–4778. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kannan, K.; Pitchiah, S.; Joseph, J.G.; Ganapathy, D.; Sundarrajan, S.; Ramakrishna, S. Marine Archaeal Extracellular Polymeric Substances from Halococcus AMS12, Their Characterization, and Biological Properties. J. Mar. Sci. Eng. 2022, 10, 1788. https://doi.org/10.3390/jmse10111788
Kannan K, Pitchiah S, Joseph JG, Ganapathy D, Sundarrajan S, Ramakrishna S. Marine Archaeal Extracellular Polymeric Substances from Halococcus AMS12, Their Characterization, and Biological Properties. Journal of Marine Science and Engineering. 2022; 10(11):1788. https://doi.org/10.3390/jmse10111788
Chicago/Turabian StyleKannan, Kamala, Sivaperumal Pitchiah, Jeevankumar Guduri Joseph, Dhanraj Ganapathy, Subramanian Sundarrajan, and Seeram Ramakrishna. 2022. "Marine Archaeal Extracellular Polymeric Substances from Halococcus AMS12, Their Characterization, and Biological Properties" Journal of Marine Science and Engineering 10, no. 11: 1788. https://doi.org/10.3390/jmse10111788
APA StyleKannan, K., Pitchiah, S., Joseph, J. G., Ganapathy, D., Sundarrajan, S., & Ramakrishna, S. (2022). Marine Archaeal Extracellular Polymeric Substances from Halococcus AMS12, Their Characterization, and Biological Properties. Journal of Marine Science and Engineering, 10(11), 1788. https://doi.org/10.3390/jmse10111788