1. Introduction
The understanding of the highly complex world of immunology is a major challenge for scientists in the fields of biology, medicine, and pharmacology. For the description of biological phenomena related to the immune system, the modeling, simulation, and analysis of the immune system are considered important practices and can contribute to improved diagnostics and optimized immune treatments. For that purpose, mathematical and computational methodologies have been utilized to create models that represent biological behavior. These models can support explanations of the interaction mechanisms between pathogenic agents and the defense mechanism in intuitive and yet analyzable terms.
In the context of our research, a mathematical model is a formal model describing by means of equations relationships between quantities and how quantities change over time. Computational models such as agent-based models (ABMs) describe dependencies between activities of components of a system. Combinations of the two approaches have been used to describe, simulate, and analyze the networks and interactions in the immune system [
1,
2,
3,
4,
5]. Pappalardo
et al. provided an extensive study on vaccine administration and immune response to cancer in mice by implementing and simulating models using ABMs and cellular automata [
6,
7,
8]. Gammack
et al. [
9] provided a mathematical model based on Ordinary Differential Equations (ODEs) to investigate the early and initial immune response to mycobacterial infection (Mtb) in mice. This work has inspired Segovia-Juarez
et al. [
10] to implement the ODEs that regulate the interaction between host and pathogen using an ABM approach. Warrender
et al. [
11] use the CyCell simulator tool to simulate the interactions in early mycobacterial infection.
The different scales of the models can be an obstacle to modeling the infection process and the immune response. For biologists, it is intuitive to establish a process that involves cells, molecules, and/or organs. It is, however, not trivial to identify the line of information retrieval that connects/switches from one level to the next [
12]. A multi-scale approach has been used to connect interactions at molecular, cellular, and tissue level as well as the modeling of the dynamics from a spatial and temporal perspective. Multi-scale models, including how to connect different individual levels, have been extensively explored, e.g., [
12,
13,
14,
15]. In the study of the mycobacterial infection process and immune response, establishing an adequate method to account for multi-scale processes is still a challenge. In mathematical models, like those based on differential equations, the interactions are described in rules and equations, whereas computational models are embedded in programming code with which one cannot interact in a straightforward manner; neither can one directly understand its structure.
Thus, a graphical representation of the interactions and influences among various components that involve the bacteria and host immune cells,
i.e., molecules and proteins, that also captures the dynamics of the system would be very useful. The Petri net formalism (PN) represents a well-established technique in which a graphical representation is combined with a mathematical basis for modeling distributed concurrent systems [
16,
17]. Petri nets are successfully used to model biological behavior [
18,
19]. Heiner
et al. [
20] propose a methodology of incremental modeling using Petri nets to develop and analyze a qualitative model of the apoptotic pathway. Albergante
et al. [
21] have developed a Petri net model that simulates the formation of hepatic granuloma for
Leishimania donavani infection in mice.
In previous work [
22], we have developed a qualitative Petri net model of the mycobacterial infection process and the subsequent innate immune response. We organized our model at the level of cell dynamics, and it is characterized by steps involved in the
Mycobacterium marinum infection and granuloma formation in zebrafish. Subsequently, we used Petri nets to model the interactions between the bacteria and the host immune cell in a multi-scale approach [
23], connecting important pathways involved in the host-pathogen interactions that are acting over different scales (molecular, intracellular, and intercellular) during the innate immune response.
In this paper, we extend our two previous models by combining them in a hierarchical fashion, so as to jointly represent the mycobacterial infection process and innate immune response. The qualitative model captures the relationship between the pathogen and the host immune cells, i.e., the bacteria and the macrophages, from the perspective of cell dynamics down to interactions at intercellular, intracellular, and molecular level. The hierarchical model provides a visualization of the infection process from the moment the bacteria enter the host: the migration, proliferation, granuloma formation, and dissemination. Moreover, it models the signaling pathways that a macrophage employs to terminate the infection and the way the bacterium exploits those pathways to enhance its intracellular survival persistence. In this paper we introduce an additional visualization, which renders the infection process as modeled and simulated with the qualitative Petri nets on a 3D mesh model of the zebrafish. It is possible to correlate the information of the net, its structure, and its results to behavior observed in vivo. In this manner, we demonstrate the power of the Petri net formalism by modeling and animating the mycobacterial infection from the perspective of different but yet interconnected scales. By coupling the resulting Petri net model with the 3D mesh model visualization, we support an additional perspective for the interpretation of the biological process.
The remainder of this paper is structured as follows. In
Section 2 we focus on the material and methods, including a description of the colored qualitative Petri net, the history of modeling, and our extensions to the visualization of net information. In
Section 3 we provide a detailed discussion of our hierarchical Petri net model by defining of all of its constituent components and boundary conditions. In
Section 4 we show our results for both the hierarchical Petri net of the
Mycobacterium infection and the 3D visualization of the infection process as we have modeled it. Finally, in
Section 5 we present our conclusions and discuss the results as well as directions for our future work.
3. Implementation
The biological processes described in
Section 2.1 occur at different scales,
i.e., the molecular, the cellular, and the tissue level. Therefore, in this study, we have designed our qualitative colored Petri net model in a hierarchical structure with four different levels of representation. The model reproduces the dynamics of the steps that are involved in the infection process and innate immune response. It also models the signaling pathways activated by macrophages in response to bacteria and how the bacteria explore this to proliferate. The implemented levels operate as containers, where level 1 represents the large-scale model that contains the entire small-scale model. The information flow is triggered on the top level but eventually flows in both directions (top–down and bottom–up).
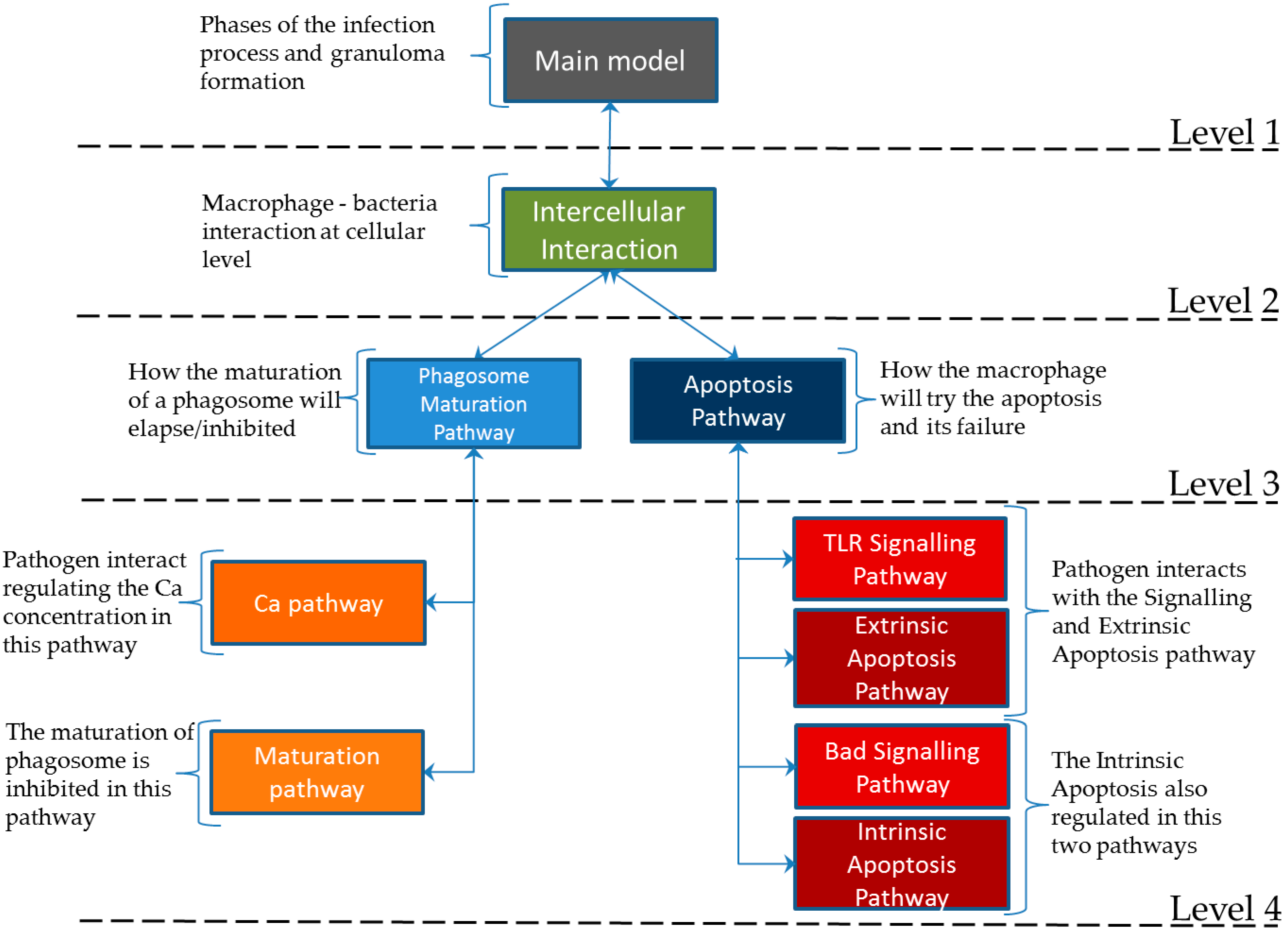
Figure 3 depicts the hierarchical structure of the net. Level 1 provides a model for the phases of the infection process: infection detection and phagocytosis, phagolysosome failure, bacterial proliferation, migration of infected macrophages to deep tissue, death of macrophages, recruitment of new immune cells and granuloma formation, intracellular spread, and granuloma dissemination. At level 2 we have modeled the bacteria and macrophage interaction on the intercellular scale. This is directly connected to level 3, in which we model the interactions at the intracellular scale; here, the important pathways related to the bacterial proliferation are modeled. At level 4 we have assembled the important interactions (from the molecular perspective) influencing the infection process.
Figure 3.
Hierarchical structure of the model. The levels are implemented as independent and interconnected subnets, following the structure of [
23].
Figure 3.
Hierarchical structure of the model. The levels are implemented as independent and interconnected subnets, following the structure of [
23].
Snoopy supplies features for the design and systematic construction of larger Petri nets. In hierarchically structured nets, coarse places and coarse transitions hide place-bordered (transition-bordered) subnets. Through these, it is possible to zoom in the model to a specific submodel and check a local behavior.
In the following sections, we present the color-set , places , transitions , and the initial marking from the main model i.e., level 1 as well as the coarse places and transitions that compose our hierarchical structured model, a qualitative colored Petri net specified as .
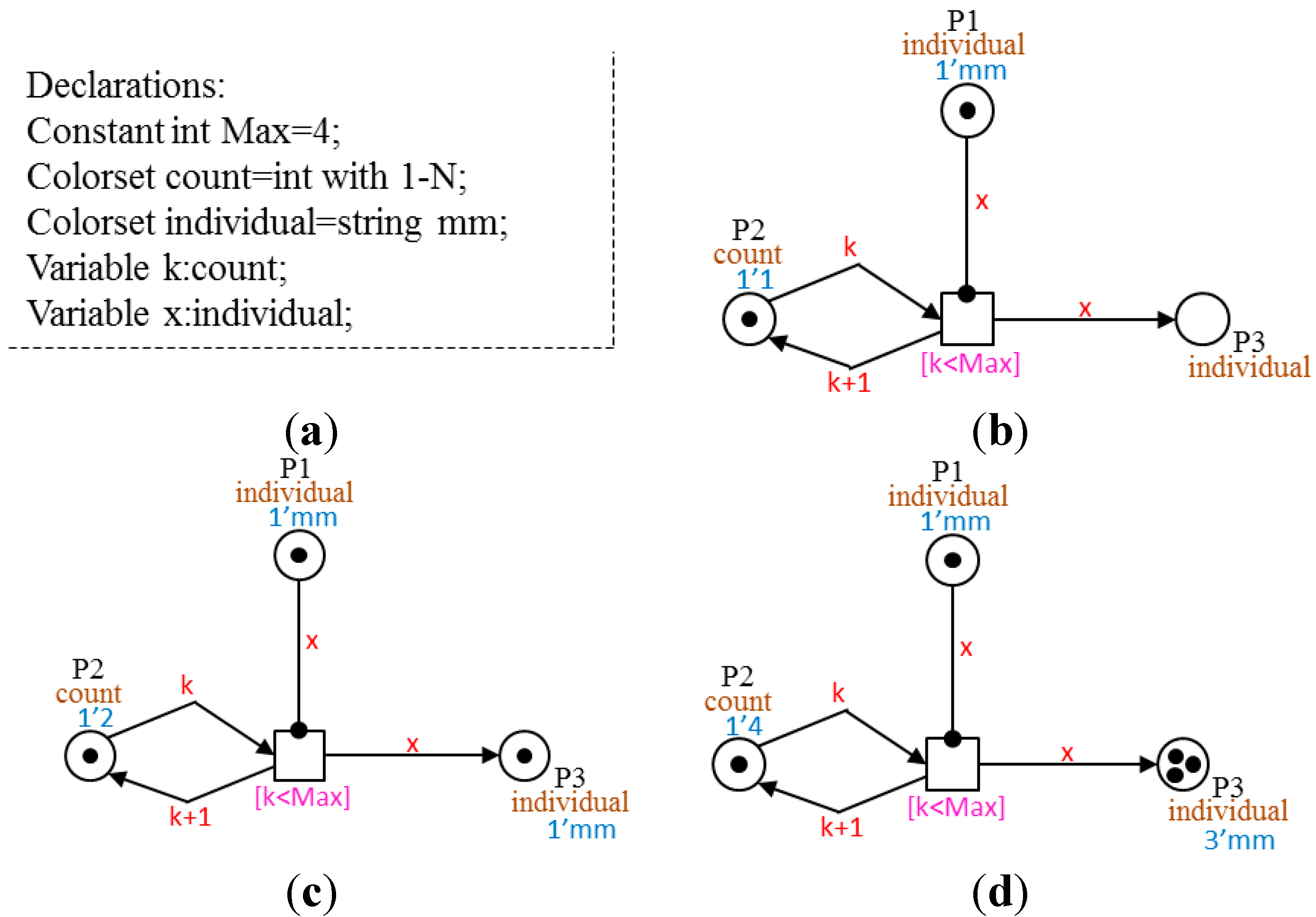
3.1. Set of Color-Sets Σ
The set of color-sets from [
22] is extended with a new color-set, Dot. This is a color-set with only one element as a color. It is used to define discrete numbers of markings that represent the presence or absence of a molecule, protein or cell in the sublevels of our model.
Table 1 describes the five simple color-set defined in our model. We introduce three compound color-sets as product of simple, predefined color-sets.
Table 2 describes the compound color-sets of the model.
Table 1.
Simple color-sets defined for the colored Petri net model.
Table 1.
Simple color-sets defined for the colored Petri net model.
| | Color-Set | Data Type | Description |
|---|
| 1 | position | Integer | Represents a location (of a macrophage, bacteria, and/or granuloma) within the zebrafish embryo |
| 2 | individual | String | Distinguishes bacteria and macrophages (Mm, Mac), respectively |
| 3 | status | Boolean | Represents the infection status of a macrophage, i.e., healthy = true; infected = false |
| 4 | count | Integer | Represents a threshold for the simulation in the recruitment of macrophage and the dissemination of a granuloma |
| 5 | Dot | dot | Default black color, used to evaluate a condition being true or false for a specific molecule or protein, or to count a number of cells |
Table 2.
Compound color-sets defined for the colored Petri net model.
Table 2.
Compound color-sets defined for the colored Petri net model.
| | Color-Set | Product Type | Description |
|---|
| 1 | Bacteria | position, individual | Represents the M. marinum bacteria that are modeled |
| 2 | Macrophage | position, individual, status | Represents host macrophage immune cells |
| 3 | Granuloma | position, individual, count | Represents granuloma with a number of infected macrophages |
3.2. Set of Places P
Places represent a population of cells and multicellular complexes that are integrated in our model. The set of places
P that composes level 1 of our model are defined as:
These places are defined and described in
Table 3.
Table 3.
Set of places composing level 1 of the model.
Table 3.
Set of places composing level 1 of the model.
| | Place | Color-Set | Description |
|---|
| 1 | Infection | Bacteria | Represent the initial infection site. It contains the initial mycobacteria that intrude the host |
| 2 | ImmuneSystem | Macrophage | Represents the immune cells of the zebrafish. It contains the non-infected macrophage cells that respond to an infection or recruitment signaling |
| 3 | InfectionPosition | Bacteria | Maintains the information of the initial position of the infection. This is important for the 3D visualization to identify where the infection initially occurs |
| 4 | InfectedMacrophage | Macrophage | Represents the macrophage after phagocytosis of bacteria. |
| 5 | ActiveMacrophage | Macrophage | Represents the infected macrophage moving in the blood circulation. It shows the macrophage changing its position while the bacteria proliferate |
| 6 | DeadMacrophage | Macrophage | Represents the necrotic macrophages positioned in the tissue after been completely infected and signaling to new immune cells to take the infection |
| 7 | RecruitedMacrophage | count | Counts/Controls the amount of healthy macrophages that are recruited to form the granuloma |
| 8 | FormedGranuloma | Granuloma | Represents the formation of granuloma. It contains the information about the granulomas formed with their specific position and amount of macrophages. |
| 9 | MatureGranuloma | Macrophage | Represents the spread of the bacteria inside the granuloma. It contains information about the granulomas, i.e., their positions and amount of macrophages (concentration) of each granuloma |
| 10 | MacrophageDisseminated | count | Counts/Controls the amount of infected macrophages that will leave the granuloma to disseminate the infection on another position |
| 11 | IntracellularInteraction | Dot | Coarse place that holds the hierarchical subnets, i.e., the connected hierarchical layers in the model and represents the bacterial-macrophage interaction at the intercellular, intracellular, and molecular level, as defined in [23] |
3.3. Set of Transitions T
The set of transitions
T of level 1 of our model is defined as:
These describe important phases in the infection process and are regulated by thresholds that control the simulation.
Table 4 describes these transitions and their guards (if present).
3.4. Initial Marking I
An initial marking defines the number and type of colored tokens that are initially present in a specific place. For the modeling of the
Mm infection, we have defined initial makings in our previous work [
22]. There exist two types of markings,
i.e., (1) condition markings that are fixed and used to control the process; and (2) example markings that are not fixed and can be changed according to the experimentation with the net.
Table 5 and
Table 6, respectively, describe these types of markings.
All other places in the top-level model, i.e., level 1 of the hierarchical structure, are initially empty, meaning that there are no tokens in these places at the onset of the simulation.
Table 4.
Set of transitions composing level 1 of the model.
Table 4.
Set of transitions composing level 1 of the model.
| | Transition | Guard | Description |
|---|
| 1 | Phagocytosis | - | Fires once its pre-places contain bacteria and immune cells at the same position. It produces tokens that represent infected macrophages as well as tokens that hold information on the initial infection position for the 3D visualization tool. |
| 2 | PhagolysosomeFail | - | Responsible for the activation of the intracellular bacterial spread, modeled in the sublevels of the hierarchical structure, while the infected macrophage moves along the blood stream, i.e., changes its position. |
| 3 | Migration | - | Responsible for controlling the macrophage position change (movement throughout the blood stream). |
| 4 | MigrationDeepTissue | - | Fires once the macrophages reach the threshold within the bacteria migrating to deep tissue to form the granuloma. |
| 5 | Recruitment | - | Signals to the non-infected immune cells, i.e., healthy macrophages, to take over the dead macrophage in their specific tissue position, thereby forming the granuloma. |
| 6 | IntracellularSpread | - | Represents the maturation of the granuloma by releasing bactericidal material and infecting macrophages that form the granuloma. |
| 7 | Dissemination | I ≤ MaxDissemination | Controls the threshold of the amount of infected macrophages that will leave the granuloma and disseminate the infection, forming a new granuloma on a different position. |
Table 5.
Condition markings initially defined in the colored Petri net model.
Table 5.
Condition markings initially defined in the colored Petri net model.
| | Place | Marking | Description |
|---|
| 1 | RecruitedMacrophage | 1`(1) | Initializes the counting of the number of macrophages recruited to aggregate into the dead macrophage. It has a threshold defined by a constant MaxAggregation i.e., 5. |
| 2 | MacrophageDisseminated | 1`(1) | Initializes the counter of the amount of infected macrophages that leave the granuloma and spread the infection to different positions. It has a threshold defined by a constant MaxDissemination i.e., 3. |
Table 6.
Example markings initially defined in the colored Petri net model.
Table 6.
Example markings initially defined in the colored Petri net model.
| | Place | Marking | Description |
|---|
| 1 | Infection | 1`(1,mm) ++1`(2,mm) ++1`(3,mm) | Defines the initial concentration of mycobacteria that will intrude the host. We have defined three different positions to represent different initial infection sites. |
| 2 | ImmuneSystem | 1`(1,mac,true) ++1`(2,mac,true) ++…++1`(12,mac,true) | Defines the initial concentration of non-infected macrophages in the host. The positions and amount are empirical information used just to represent their presence in the host. |
3.5. Sub-Models in the Hierarchical Structure
Following the tree of the hierarchical structure, the submodels, implemented through the coarse place IntracellularInteraction, model a complex process involving various host-bacterial factors in a cross-talk interaction distributed in the sublevels. In order to get a consistent view of the entire interaction process, we express the most important reactions by simplifying the pathways at different levels of abstraction. The simplification of the pathways corresponds to our previously defined modeling decisions [
23], where each biochemical compound or receptor is defined as a place. The relations between biochemical substances are represented by transitions with corresponding arcs for the reactions. Inhibitions and degradations are represented by inhibitor arcs. Signaling and catalytic atomic events are represented by read-arcs. We specifically use the color-set Dot to represent the presence of a protein, component, and/or cell that is involved in the interaction process between the macrophage and bacteria once it is phagocytosed. To hierarchically connect the different pathways we use coarse transitions and coarse places structuring all the sublevels, as shown in
Figure 3.
4. Results
The simulation environment for the model represents the innate immune response to
M. marinum infection in the zebrafish embryo. The elements of the qualitative colored Petri net described in the previous sections represent key factors involved in the processes of infection, innate immune response, and granuloma formation. Moreover, the interactions are represented by the firing rules that describe the behavior of the model:
- (i)
Signaling of the intruding bacteria, detected by non-infected macrophages followed by phagocytosis;
- (ii)
Migration of the infected macrophage to the deep tissue within bacterial replication motivated by the phagolysosome failure in the macrophage, causing the cell death;
- (iii)
Recruitment of non-infected macrophage in response to signals of the dead macrophage, clustering to form the granuloma;
- (iv)
Granuloma maturation and bacterial spread between aggregated macrophage;
- (v)
Infection dissemination through infected macrophages that escape from the matured granuloma, forming new granulomas at different positions.
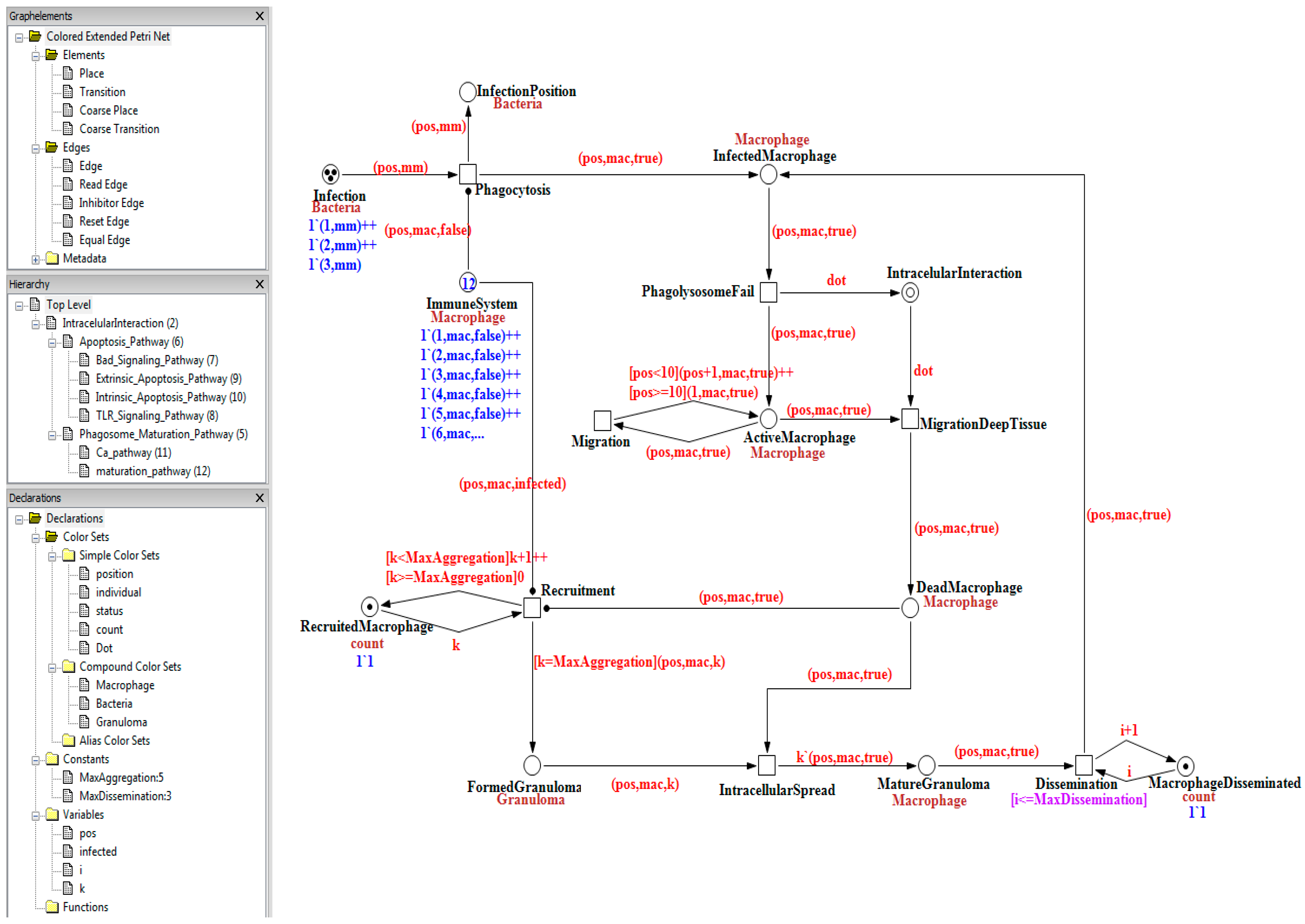
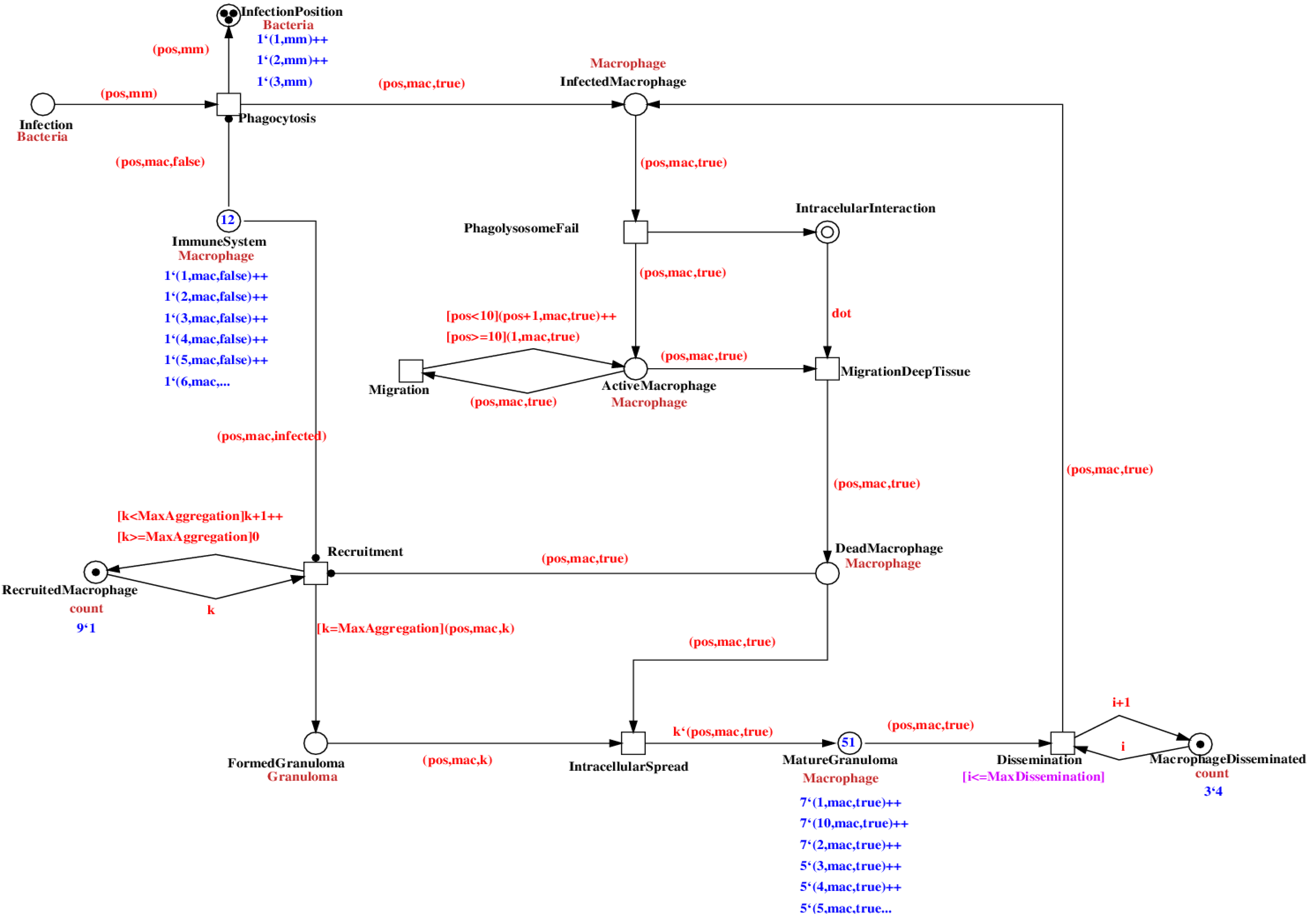
The top-layer level of our model is depicted in
Figure 4. Places are represented by circles and coarse places by double circles. Transitions are represented by squares and coarse transitions by a double square. The number of tokens is expressed inside the places and their dataset below the color-set (cf
. Figure 4, dark blue). Arrows represent the arcs labeled with their expression on top of it (cf
. Figure 4, light red). The arcs with a black dot as an arrowhead are read arcs. In the notation of our PN software environment, these are represented as two arcs in opposite directions between place and transition with an identical arc expression. However, the tokens are not consumed, just tested for their presence.
Figure 4.
Visualization of the qualitative colored Petri net model implemented in the Snoopy framework [
44]. In the left panel, information about the Petri net model is displayed: the hierarchical structure and definitions used in the model, the color-set, the constants, and variables. The main panel shows level 1 (cf
. Top Level) of the hierarchical Petri net model and its properties.
Figure 4.
Visualization of the qualitative colored Petri net model implemented in the Snoopy framework [
44]. In the left panel, information about the Petri net model is displayed: the hierarchical structure and definitions used in the model, the color-set, the constants, and variables. The main panel shows level 1 (cf
. Top Level) of the hierarchical Petri net model and its properties.
In the model, the data value of the colored Petri net formalism is used to represent the quantities that change during the simulation. Although our focus is on a qualitative model, the quantitative aspects promote a better interpretation of the dynamical aspects of the simulation. Level 1, depicted in
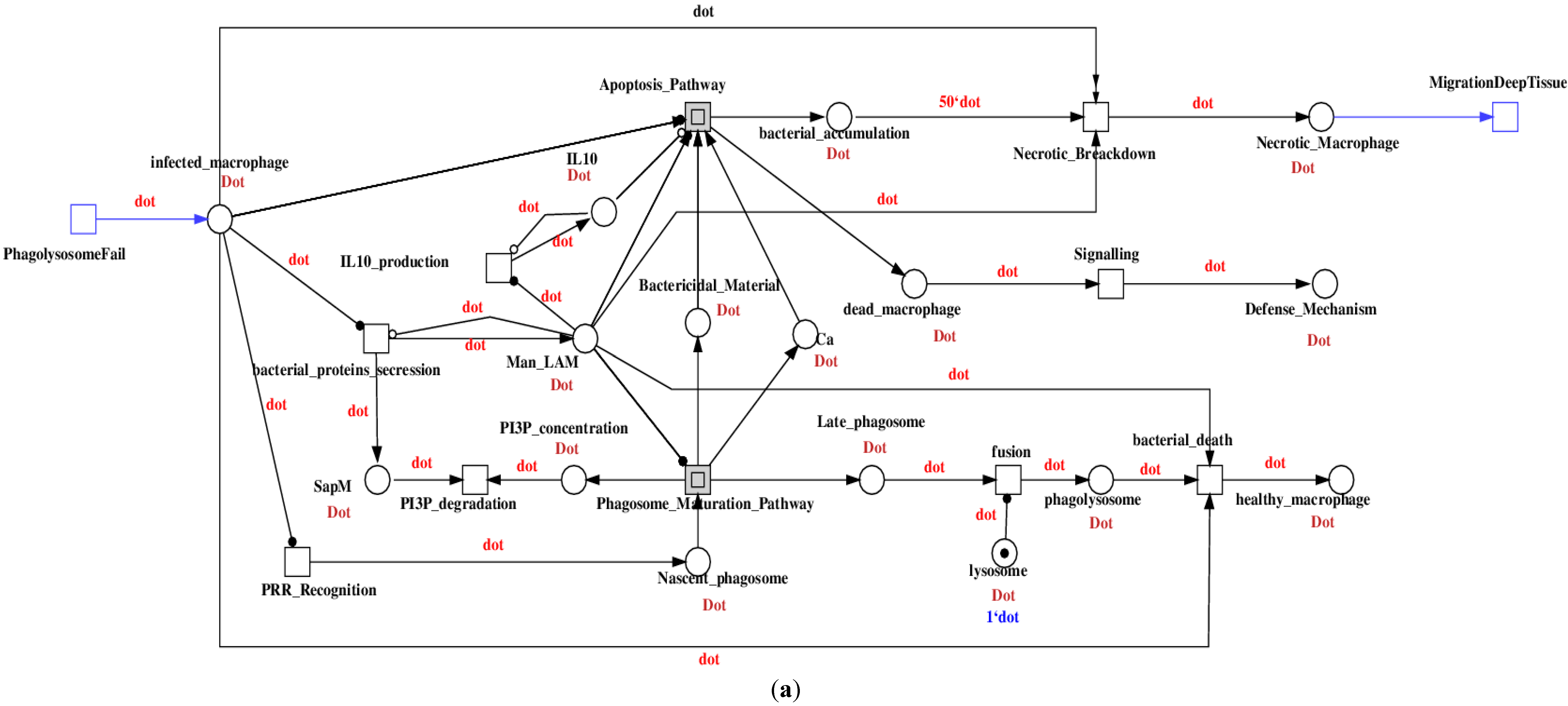
Figure 4, models the different stages of the infection process. It represents the phases of injection, phagocytosis, migration, granuloma formation, and dissemination. Expanding the hierarchical structure, the connection of the sub-models and their components are depicted in
Figure 5. Level 2 is modeled in the coarse place IntracelularInteraction of level 1 (cf
. Figure 4). It represents the intercellular interaction between mycobacterium and a macrophage and is triggered by the phagolysosome fail (cf
. Figure 5a). Level 3 models the intracellular interaction, represented by the coarse transitions: Phagosome_Maturation_Pathway (cf
. Figure 5b) and Apoptosis_Pathway (cf
. Figure 5c). These course places bridge the signaling started at the cell wall (cf. level 1) that triggers the interactions between molecules. Level 4 models the molecular interaction of the proteins released by the bacteria and the proteins from the macrophage. They are represented by the coarse places Extrinsic_Apoptosis_Pathway, Intrinsic_Apoptosis_Pathway, Ca_Pathway, Maturation_Pathway, and also the coarse transitions Bad_Signaling_Pathway and TLR_Signaling_Pathway.
We have defined some boundaries to limit the model for a better qualitative analysis of the behavior of the system. The intracellular bacterial proliferation is defined by the arc expression 2`dot, which represents the offspring of two new bacteria every time the transition
{bacterial_proliferation} fires (cf.
Figure 5b). The capacity of the infected macrophage is limited to a concentration of 50 bacteria. After the place
{bacterial_acumulation} reaches this boundary,
i.e., arc expression 50`dot, the transition
{Necrotic_Breakdown} can fire. This action changes the infected macrophage into a necrotic macrophage that will leave the bloodstream and migrate into the tissue (cf.
Figure 5a).
Another threshold used is related to the position of macrophages and granulomas. Nezhinsky
et al. [
45] have developed an image processing platform for the analysis of high-throughput screens in zebrafish. In this platform, infected zebrafish embryos are automatically recognized as a shape. Relative to the shape, the bacterial infection is analyzed as an infection spread; to this end, the shape is divided into 12 regions of infection. This research has inspired us to determine 12 relative positions based on this division to model the presence of the macrophages, their movement during the infection process, and also the spread of the granuloma. The concentration of aggregated macrophages is limited by setting a constant called
MaxAggregation. Moreover, the infection dissemination is limited further by setting a constant
MaxDissemination (cf
. Figure 4). We use these thresholds to control the amount of cells that form the granuloma, as well as the number of dissident infected macrophages that are released from the granuloma to spread the infection to other positions.
The outcome of our model reproduces the early stages of the mycobacterial infection process and innate immune response. We use the animation mode available in the Snoopy framework to verify the dynamic behavior of our model. This property enables the animation of token flow through the net, in order to observe the causality of the model and its behavior on the whole hierarchical structure. To show the importance of the immune cells, molecules, and processes in the dynamics of the infection, we performed a simulation based on our specific experimental scenario, defined in
Section 2.1.
Figure 5.
Submodels in the hierarchical structure: (
a) Level 2: Intercellular interaction between bacteria and macrophage. (
b) Apoptosis_Pathway and (
c) Phagosome_Maturation_Pathway represent the intracellular interaction positioned at level 3. Level 4 represents the molecular interactions defined in [
23]; they are modeled in the coarse places and coarse transitions (data not shown).
Figure 5.
Submodels in the hierarchical structure: (
a) Level 2: Intercellular interaction between bacteria and macrophage. (
b) Apoptosis_Pathway and (
c) Phagosome_Maturation_Pathway represent the intracellular interaction positioned at level 3. Level 4 represents the molecular interactions defined in [
23]; they are modeled in the coarse places and coarse transitions (data not shown).
We start the simulation by defining the amount of bacteria and their initial position at the place of infection, adding the initial markings: {1`(1,mm)++ 1`(2,mm)++ 1`(3,mm)}. The ImmuneSystem place, which contains macrophages, sends to the initial infected position a non-infected macrophage to phagocytose the bacteria. The macrophage becomes infected through a failure of the phagolysosomal process. Next, in the sublevels of the net the proliferation of bacteria will occur while the infected macrophage is migrating along the blood circulation. In the net, the proliferation process is triggered by sending a token 1`dot, to the coarse place IntracelularInteraction. At level 2, the infected macrophage attempts to kill the bacteria by activating the phagosome maturation or apoptosis process. The signaling process starts by sending a token from the place Nascent_Phagosome to the coarse transition Phagosome_Maturation_Pathway, and from the place Bactericiadal_Material to the coarse transition Apoptosis_Pathway. At level 3, these pathways trigger molecular interaction, by sending tokens to the coarse places and coarse transitions that model the protein–protein interactions at level 4. The bacteria interact with both pathways by releasing the ManLAM as presented by the place Man_LAM. It escapes being killed and instead proliferates inside the macrophage (cf. place Bacterial_acumulation), causing a macrophage necrotic breakdown. At this moment the necrotic macrophage leaves the blood circulation (at level 1) and migrates to deep tissue. There it will recruit non-infected macrophages to form a granuloma (cf. place Formed_Granuloma). The bacteria will spread inside the granuloma, and will disseminate the infection by releasing an infected macrophage into the blood circulation. These processes repeat for each infected macrophage that leaves the granuloma.
The amount of infected macrophages leaving the granuloma is limited through the constant
MaxDissemination. Once this threshold is reached, no further transition is enabled to occur and the execution will terminate in its final state. Since our model is focused on the qualitative aspects of the bacterial infection process and innate immune response, we bound our simulation by this constant to represent the initial dissemination of the infection. The final state then provides the quantity of granuloma formed during this process and their respective positions, accumulated at the place MatureGranuloma. In
Figure 6 the final state of the net is depicted; an animation sequence of this PN can be found in [
46]. By saving the final state result of the CPN it becomes possible to derive the complete state space from the Petri net file. This file is subsequently used as the input for the 3D visualization of the process. The Snoopy file can be obtained upon request.
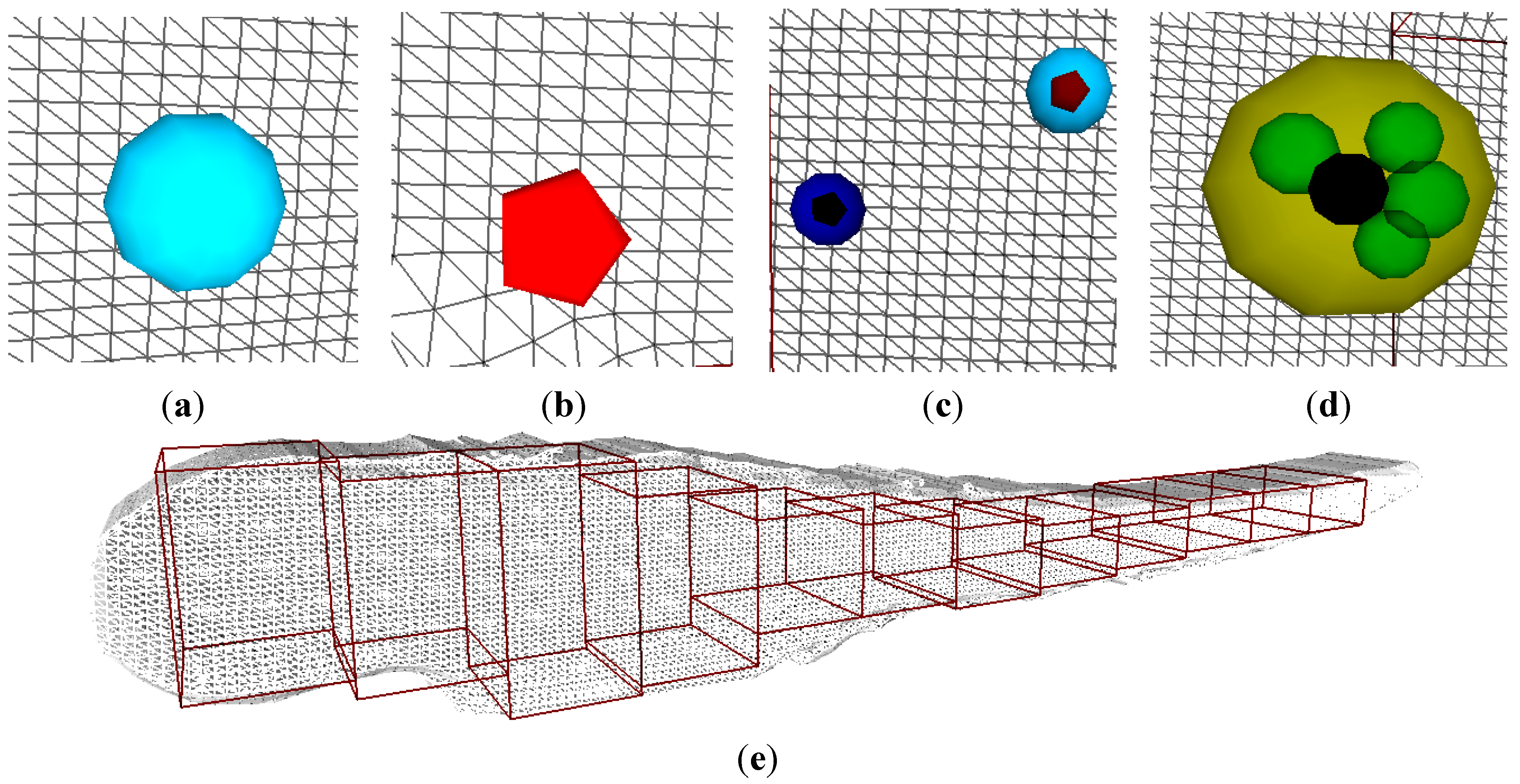
The 3D visualization tool was built to provide an alternative animation of the Petri net. We defined graphical elements to represent bacteria, macrophages, and granulomas, whereas a 3D mesh object represents the zebrafish embryo. The 3D visualization of the infection process plays the exact same simulation as the animation of the colored Petri net model. We define 12 regions in the mesh to represent the positions where infection can occur. The positions are relative to spatial enumerations in the zebrafish model and normalized to its total length. The 3D animation is played over the entire volume of the 3D mesh. The 3D animation visualization software arranges the positioning of the objects in such a way that there is no overlap between the objects, nor do they mutually collide. In
Figure 7, the graphical elements and the object environment of the 3D visualization are depicted.
Figure 6.
Animation mode at its final state, where there are no more enabled transitions to occur. We save this state and use the file as input to our 3D visualization tool.
Figure 6.
Animation mode at its final state, where there are no more enabled transitions to occur. We save this state and use the file as input to our 3D visualization tool.
Figure 7.
3D visualization tool and its elements: (a) Macrophage; (b) Bacteria; (c) Infected macrophages; (d) Granuloma; (e) Fish embryo with the 12 regions where infection can occur.
Figure 7.
3D visualization tool and its elements: (a) Macrophage; (b) Bacteria; (c) Infected macrophages; (d) Granuloma; (e) Fish embryo with the 12 regions where infection can occur.
In the 3D animation, the initial bacteria are introduced according to the information of the Petri net model. It reads the colored tokens in the place InfectionPosition, normalizing the position values and placing the bacteria objects in the predefined regions accordingly. In the Petri net model, the non-infected macrophages are in all of the 12 possible regions as they are transported through the blood vessels. The granuloma formation and dissemination occur on the basis of the information from the colored tokens as present in the place MatureGranuloma. The constants
MaxAggregation and
MaxDissemination bound the infection process. The animation is time-independent and sequentially follows the infection steps as defined in the model. The 3D animation renders the dynamics of infection process and granuloma formation according to the final state space from the Petri net. For inspection and perusal, an animation sequence can be found in [
46]. In
Figure 8a comparison is provided of the 3D visualization with the Petri net data with a real zebrafish embryo that has been infected with
Mm. The microscope image of a seven-day-old zebrafish depicted in
Figure 8a is the result of an analysis for the spread of the
Mm bacteria after six days of infection (
dpi). This result was obtained with specific software and the granulomas extracted from the image are overlaid as magenta blobs [
27,
45]. The final result of the 3D animation visualization is depicted in
Figure 8b. As one can appreciate here, the simulation reproduces the distribution of the granulomas resulting from the infection process along the fish, in a similar pattern as occurs in the real zebrafish. Therefore, it is possible to correlate our final result with
in vivo experiments although we are not using quantitative data. The threshold for manipulating the dynamics of the CPN model is the only quantitative information that has been used, and this threshold is not directly based on the analysis of empirical data. We, therefore, cannot directly compare the amount of granuloma or the distribution of the infection as obtained from our CPN model to
in vivo situations. However, we can extrapolate from the results of our simulation that the structure of our Petri net model qualitatively represents the behavior of the
in vivo infection process.
Figure 8.
(a) Microscope image of a seven-day-old zebrafish (7 dpf) infected with wild-type Mm bacteria one day old, referred to as six days post-infection (6 dpi). The granulomas are displayed as magenta blobs. (b) The end state of the 3D animation with all the granulomas formed according to the final state space of the colored Petri net model.
Figure 8.
(a) Microscope image of a seven-day-old zebrafish (7 dpf) infected with wild-type Mm bacteria one day old, referred to as six days post-infection (6 dpi). The granulomas are displayed as magenta blobs. (b) The end state of the 3D animation with all the granulomas formed according to the final state space of the colored Petri net model.
5. Conclusions and Discussions
The model we presented in this work is concerned with an infection scenario: the innate immune response to M. marinum in zebrafish. It represents the dynamics of bacterial proliferation, granuloma formation, and dissemination. The model captures the relevant functional processes and their interconnections, including signaling and activation or inhibition of the immune responses at different levels of abstraction. In sublevels, we have connected the most important pathways in order to model the response of the macrophage on the exposure to pathogenic mycobacteria. Information about the proteins released by the bacteria, their interference with the immune response, and the pathways involved in this process are also taken into account.
We focused on modeling the qualitative aspects of the infection process through connecting two complementary models: one for the reaction process and biochemical components implemented as a qualitative Petri net (without colors) [
23]. The other model [
22] represents the dynamics of the cells in the infection process, which is implemented as a colored Petri net. The resulting hierarchical Petri net model covers the relevant phases of the infection process, as well as the interaction pathways related to the infection persistence.
In order to combine the two models into one net model, the QPN model [
23] was first converted into a QPN
C model. The color-set Dot was introduced to represent the presence of signaling molecules, proteins, and concentrations of cells involved in the interaction between macrophage and bacteria that are phagocytosed. Through this color-set, the two models were combined in one modular structure. Using the Snoopy platform in animation mode, the combined hierarchical model could be executed.
The animation mode of Snoopy allows one to verify the dynamics of an animated Petri net by analyzing the markings encountered during a simulation. This showed that our Petri net model was able to replicate the steps in the infection process. This is due to the fact that the structure of the Petri net is based on the actual description of the infection process as extracted from the literature. Hence, the qualitative aspects in the sense of cause-and-effect are in accordance with the actual process. The quantities used in the model have no direct significance but to support the qualitative modeling and serve only as threshold. Neither quantities nor time are essential to the working of the model.
From a biological perspective, the additional 3D visualization environment has proven to be an interesting complementary approach to illustrate the infection process. The possibility of visualizing the infection, from the introduction of bacteria to granuloma formation as it would happen in vivo, provides a better understanding of the infection process.
A comparison of the simulation results with an
in vivo experiment (an infected zebrafish at 6 dpi) confirms once more the strength of the qualitative aspects of our model (cf.
Figure 8).
Because the 3D visualization tool reads markings of the Petri net model, changes in the initial marking of the net are directly reflected in the 3D visualization. Thus it can lead to more insight into the infection process, e.g., how dissemination and concentration of granulomas depend on the initial position of the bacteria, the amount of aggregation cells that form a granuloma, as well as on the number of infected macrophages that can leave the granuloma. By changing the number and/or positions of tokens, “what-if” scenarios can be executed to represent different biological conditions. Disruptions of pathways, the presence or absence of proteins, and also different positions of cells can be simulated as part of the experimentation in the animation mode.
For the further development of “what-if” scenarios, however, a quantitative model is necessary that, through support of empirical data, would enable us to do quantitative analysis as well as perform simulations and predictions. An advantage of our Petri net model is its modularity, facilitating the conversion from a qualitative model to a quantitative model without changing the structure. Since the 3D visualization can perform an animation based on the final state space of the net, it would then provide an even more realistic perspective of the infection process. Currently, we are collecting and analyzing data from zebrafish infection studies to use these as a basis for such a quantitative model. As a next step, we will integrate these data in our simulation framework and will then be able to perform different scenarios as part of the simulation process. This will contribute to identifying important parameters that can help to unravel mechanisms related to the mycobacterial infection process and innate immune response.
In summary, in this paper, we have presented the coupling of two distinct models of different aspects of the mycobacterial infection process and innate immune response. To understand infection and the immune response, it is necessary to analyze the process from its epidemiology down to genetic levels. We have modeled the dynamics of the infection and also the intracellular, intercellular, and molecular interactions, interconnecting the models in a hierarchical structure. The resulting multi-scale Petri net model allows for the observation of events at a given scale and how they interact with higher or lower levels. In addition, we provided an animation of the model using the Snoopy framework and related the Petri net output to a 3D environment. The hierarchical structuring of information in Petri net has the potential to become an important tool for research into infectious diseases, tuberculosis in particular.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






