The Evolution and Future Development of Attention Networks


Abstract
:1. Introduction
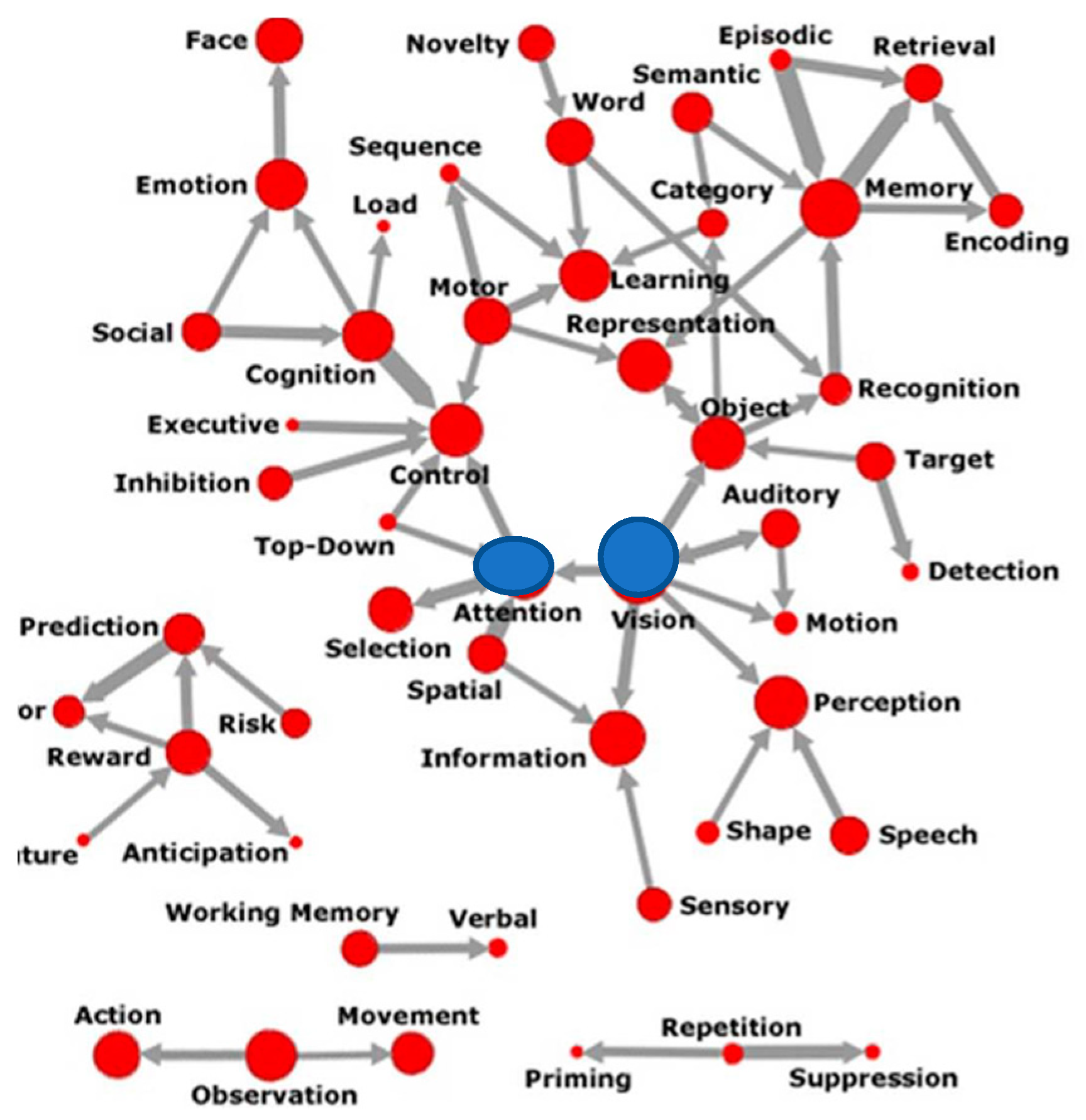
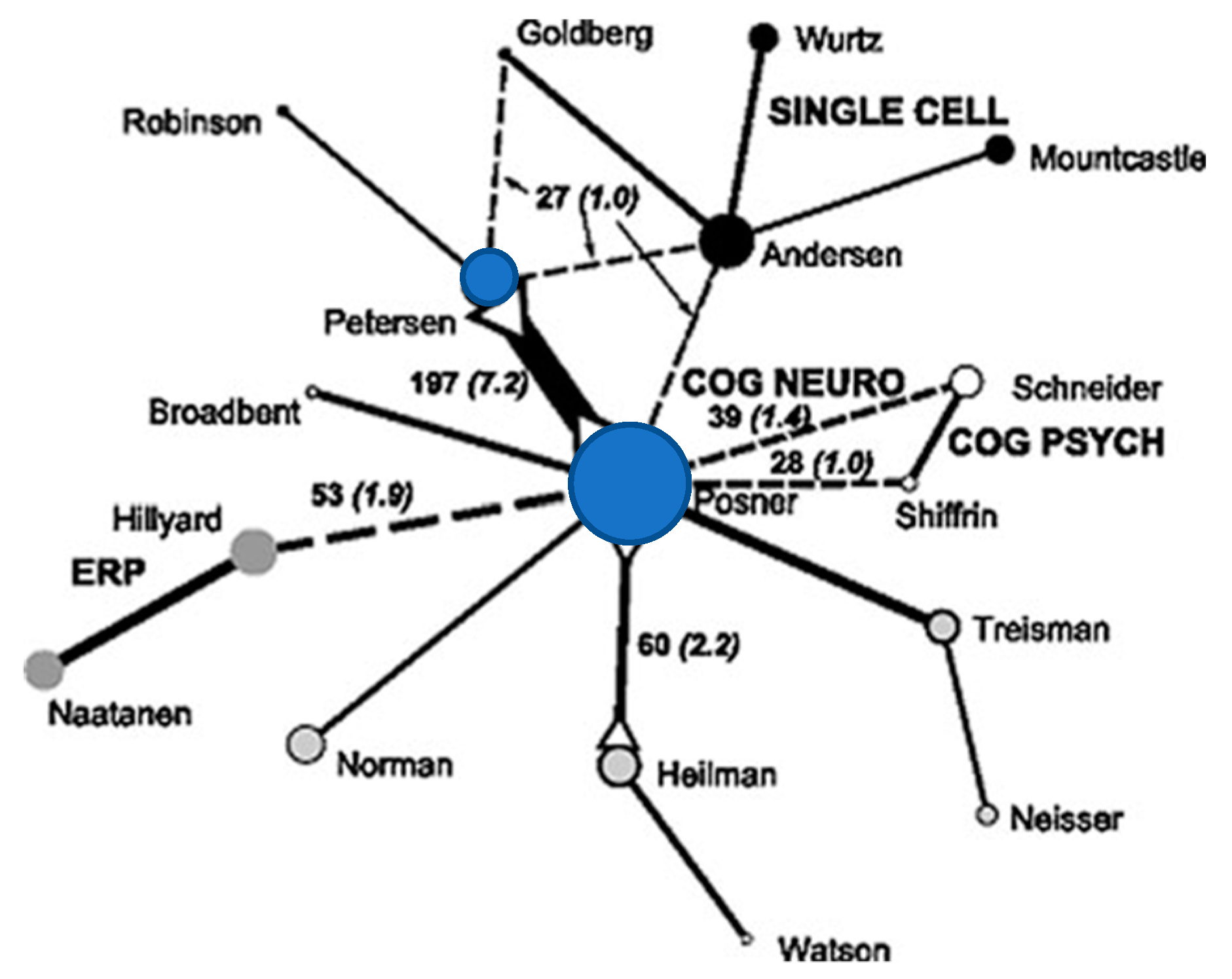
2. Centrality of Attention Networks
3. Evolution of Attention
3.1. Animal Studies
3.2. Human Development
3.3. Separability of Covert and Overt Attention
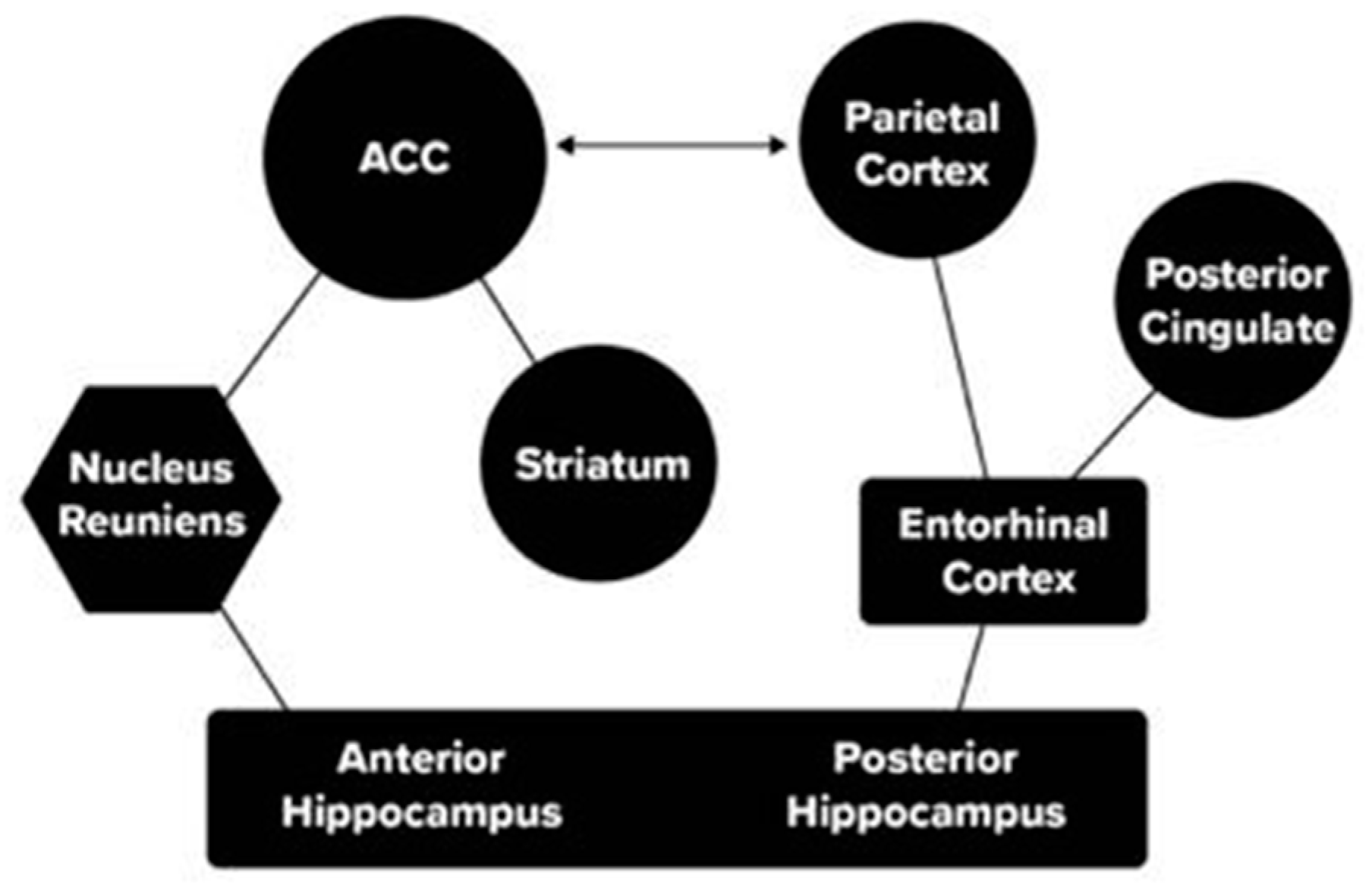
4. Interaction of Networks
Molecular Level
5. Future Studies
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alexander, Andrew S., Ryan Place, Michael J. Starrett, Elizabeth R. Chrastil, and Douglas A. Niz. 2022. Rethinking retrosplenial cortex: Perspectives and predictions. Neuron 111: 150–75. [Google Scholar] [CrossRef] [PubMed]
- Beam, Elizabeth, L. Gregory Appelbaum, Jordynn Jack, James Moody, and Scott A. Huettel. 2014. Mapping the Semantic Structure of Cognitive Neuroscience. Journal of Cognitive Neuroscience 26: 1949–65. [Google Scholar] [CrossRef] [PubMed]
- Beane, Melinda, and Richard Marrocco. 2004. Cholinergic and noradrenergic inputs to the posterior parietal cortex modulate the components of exogenous attention. In Cognitive Neuroscience of Attention. Edited by Michael I. Posner. New York: Guilford, pp. 313–25. [Google Scholar]
- Bowman, Caitlin R., and Dagmar Zeithamova. 2018. Tracking the development of specific and generalized representations during concept learning. Journal of Neuroscience 38: 2605–14. [Google Scholar] [CrossRef] [PubMed]
- Bruer, John T. 2010. Can we talk? How the cognitive neuroscience of attention emerged from neurobiology and psychology, 1980–2005. Scientometrics 83: 751–64. [Google Scholar] [CrossRef]
- Callejas, Alicia, Juan Lupianez, and Pío Tudela. 2004. The three attentional networks: On their independence and interactions. Brain and Cognition 54: 225–27. [Google Scholar] [CrossRef]
- Callejas, Alicia, Juan Lupianez, María J. Funes, and Pío Tudela. 2005. Modulations among the alerting, orienting and executive control networks. Experimental Brain Research 167: 27–37. [Google Scholar] [CrossRef]
- Corbetta, Maurizio, and Gordon L. Shulman. 2002. Control of goal-directed and stimulus-driven attention in the brain. Nature Neuroscience Reviews 3: 201–15. [Google Scholar] [CrossRef]
- Corbetta, Maurizio, Erbil Akbudak, Thomas E. Conturo, Abraham Z. Snyder, John M. Ollinger, Heather A. Drury, Martin R. Linenweber, Steven E. Petersen, Marcus E. Raichle, David C. Van Essen, and et al. 1998. A common network of functional areas for attention and eye movements. Neuron 21: 761–73. [Google Scholar] [CrossRef]
- Desimone, Robert, and John Duncan. 1995. Neural mechanisms of selective visual attention. Annual Review of Neuroscience 18: 193–222. [Google Scholar] [CrossRef]
- Fan, Jin, Bruce D. McCandliss, John Fossella, Jonathan I. Flombaum, and Michael I. Posner. 2005. The activation of attentional networks. Neuroimage 26: 471–79. [Google Scholar]
- Fan, Jin, Bruce D. McCandliss, Tobias Sommer, Amir Raz, and Michael I. Posner. 2002. Testing the efficiency and independence of attentional networks. Journal of Cognitive Neuroscience 3: 340–47. [Google Scholar] [CrossRef]
- Fan, Jin, Xiaosi Gu, Kevin G. Guise, Xun Liu, John Fossella, Hongbin Wang, and Michael I. Posner. 2009. Testing the behavioral interaction and integration of attentional neworks. Brain and Cognition 70: 209–20. [Google Scholar] [CrossRef] [PubMed]
- Fjell, Anders M., Kristine Beate Walhovd, Timothy T. Brown, Joshua M. Kuperman, Yoonho Chung, Donald J. Hagler, Vijay Venkatraman, J. Cooper Roddey, Matthew Erhart, Connor McCabe, and et al. 2012. Multimodalimaging of the self-regulating brain. Proceedings of the National Academy of Sciences 109: 19620–25. [Google Scholar] [CrossRef] [PubMed]
- Franco, Luis M., and Michael J. Goard. 2021. A distributed circuit for associating environmental context with motor choice in retrosplenial cortex. Science Advances 2021: eabf9815. [Google Scholar] [CrossRef] [PubMed]
- Gabay, Shai, Tali Leibovich, Avi Ben-Simon, Avishai Henik, and Ronen Segev. 2013. Inhibition of return in the archer fish. Nature Communications 4: 1657. [Google Scholar] [CrossRef] [PubMed]
- Gregoriou, Georgia G., Stephen J. Gotts, and Robert Desimone. 2012. Cell-Type-Specific Synchronization of Neural Activity in FEF with V4 during Attention. Neuron 73: 581–94. [Google Scholar] [CrossRef] [PubMed]
- Hassett, Thomas C., and Robert R. Hampton. 2022. Control of Attention in Rhesus Monkeys Measured Using a Flanker Task. Attention Perception and Psychophysics 84: 2155–66. [Google Scholar] [CrossRef] [PubMed]
- Li, Shuting, C. May, A. J. Hannan, K. A. Johnson, and E. L. Burrows. 2021. Assessing attention orienting in mice: A novel touchscreen adaptation of the Posner-style cueing task. Neuropsychopharmacology 46: 432–41. [Google Scholar] [CrossRef] [PubMed]
- Posner, Michael I. 1978. Chronometric Explorations of Mind. Hillsdale: Lawrence Erlbaum Associates. [Google Scholar]
- Posner, Michael I. 1988. Structures and functions of selective attention. In Master Lectures in Clinical Neuropsychology and Brain Function: Research, Measurement, and Practice. Edited by T. Boll and B. Bryant. Worcester: American Psychological Association, pp. 171–202. [Google Scholar] [CrossRef]
- Posner, Michael I., Aldis P. Weible, Pascale Voelker, Mary K. Rothbart, and Cristopher M. Niell. 2022. Decision Making as a Learned Skill in Mice and Humans. Frontiers in Neuroscience 16: 834701. [Google Scholar]
- Posner, Michael I., and Mary K. Rothbart. 1980. The development of attentional mechanisms. In Nebraska Symposium on Motivation. Edited by J. H. Flowers. Lincoln: Nebraska University Press, pp. 1–49. [Google Scholar]
- Posner, Michael I., and Steven E. Petersen. 1990. The attention system of the human brain. Annual Review of Neuroscience 13: 25–42. [Google Scholar] [CrossRef]
- Posner, Michael I., Mary K. Rothbart, Brad E. Sheese, and Pascale Voelker. 2014. Developing Attention: Behavioral and Brain Mechanisms. Advances in Neuroscience 2014: 405094. [Google Scholar] [CrossRef]
- Rizzolatti, Giacomo, Lucia Riggio, Isabella Dascola, and Carlo Umilta. 1987. Reorienting attention across the horizontal and vertical meridians: Evidence in favor of a premotor theory of attention. Neuropsychologia 25, Pt 1: 31–40. [Google Scholar] [CrossRef] [PubMed]
- Rueda, M. R., Jin Fan, Jessica Halparin, Dana B. Gruber, Lisha P. Lercari, Bruce D. McCandliss, and Michael I. Posner. 2004a. Development of attention during childhood. Neuropsychologia 42: 1029–40. [Google Scholar] [CrossRef] [PubMed]
- Rueda, M. R., Michael I. Posner, Mary K. Rothbart, and Clintin P. Davis-Stober. 2004b. Development of the time course for processing conflict: An event-related potentials study with 4 year olds and adults. BMC Neuroscience 5: 391471–2202. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, Nicholas A., and Tirin Moore. 2012. Lumping and Splitting the Neural Circuitry of Visual Attention. Neuron 73: 411–12. [Google Scholar] [CrossRef]
- Thompson, Kirk G., Keri L. Biscoe, and Takashi R. Sato. 2005. Neuronal basis of covert spatial attention in the frontal eye field. Journal of Neuroscience 25: 9479–87. [Google Scholar] [CrossRef]
- Wang, Lupeng, and Richard J. Krauzlis. 2018. Visual selective attention in mice. Current Biology 28: 676–85.e4. [Google Scholar] [CrossRef]
- Washburn, David A. 1994. Stroop-like Effects for Monkeys and Humans: ProcessingSpeed or Strength of Association? Psychological Science 5: 375–79. [Google Scholar] [CrossRef]
- Weible, Aldis P. 2013. Remembering to attend: The anterior cingulate cortex and remote memory. Behavioural Brain Research 245: 63–75. [Google Scholar] [CrossRef]
- Weible, Aldis P., Michael I. Posner, and Cristopher M. Neill. 2019. Differential involvement of three brain regions during mouse skill learning. eNeuro 6: ENEURO.0143-19.2019. [Google Scholar] [CrossRef]
- Wu, Tingting, Melissa-Ann A. Mackie, Chao Chen, and Jin Fan. 2022. Representational coding of overt and covert orienting of visuospatial attention in the frontoparietal network. NeuroImage 261: 119499. [Google Scholar] [CrossRef]
- Xu, Wei, and Thomas C. Sudhof. 2013. A neural circuit for memory specificity and generalization. Science 339: 1290–95. [Google Scholar] [CrossRef]
- Zhu, Mengjin, and Shuhong Zhao. 2007. Candidate gene identification approach: Progress and challenges. International Journal of Biological Studies 3: 420–27. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Valid | Invalid | Orienting (All in ms) | |
|---|---|---|---|
| Humans * | 380 | 410 | 30 |
| Monkeys * | 420 | 445 | 25 |
| Rats *+ | 400 | 430 | 30 |
| Mice + | 420 | 450 | 30 |
| Age | Congruent | Incongruent | Difference |
|---|---|---|---|
| 4 * | 12.4 | 21.9 | 9.5 |
| 6 + | 8 | 23.6 | 15.6 |
| 7 + | 5.4 | 5.9 | 0.5 |
| 8 + | 4.9 | 4.7 | −0.2 |
| 9 + | 1.9 | 3.5 | 1.6 |
| Network | Modulator | Genes |
|---|---|---|
| Alerting | Norepinephrine | ADRA2A, NET |
| Orienting | Acetylcholine | CHRNA4, APOE |
| Executive | Dopamine | DRD4, DAT1, COMT, MAOA, DBH |
| Serotonin | TPH2, 5HTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Posner, M.I. The Evolution and Future Development of Attention Networks. J. Intell. 2023, 11, 98. https://doi.org/10.3390/jintelligence11060098
Posner MI. The Evolution and Future Development of Attention Networks. Journal of Intelligence. 2023; 11(6):98. https://doi.org/10.3390/jintelligence11060098
Chicago/Turabian StylePosner, Michael I. 2023. "The Evolution and Future Development of Attention Networks" Journal of Intelligence 11, no. 6: 98. https://doi.org/10.3390/jintelligence11060098
APA StylePosner, M. I. (2023). The Evolution and Future Development of Attention Networks. Journal of Intelligence, 11(6), 98. https://doi.org/10.3390/jintelligence11060098