
Epigenetic Differences Arise in Endothelial Cells Responding to Cobalt–Chromium


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Experimental Design
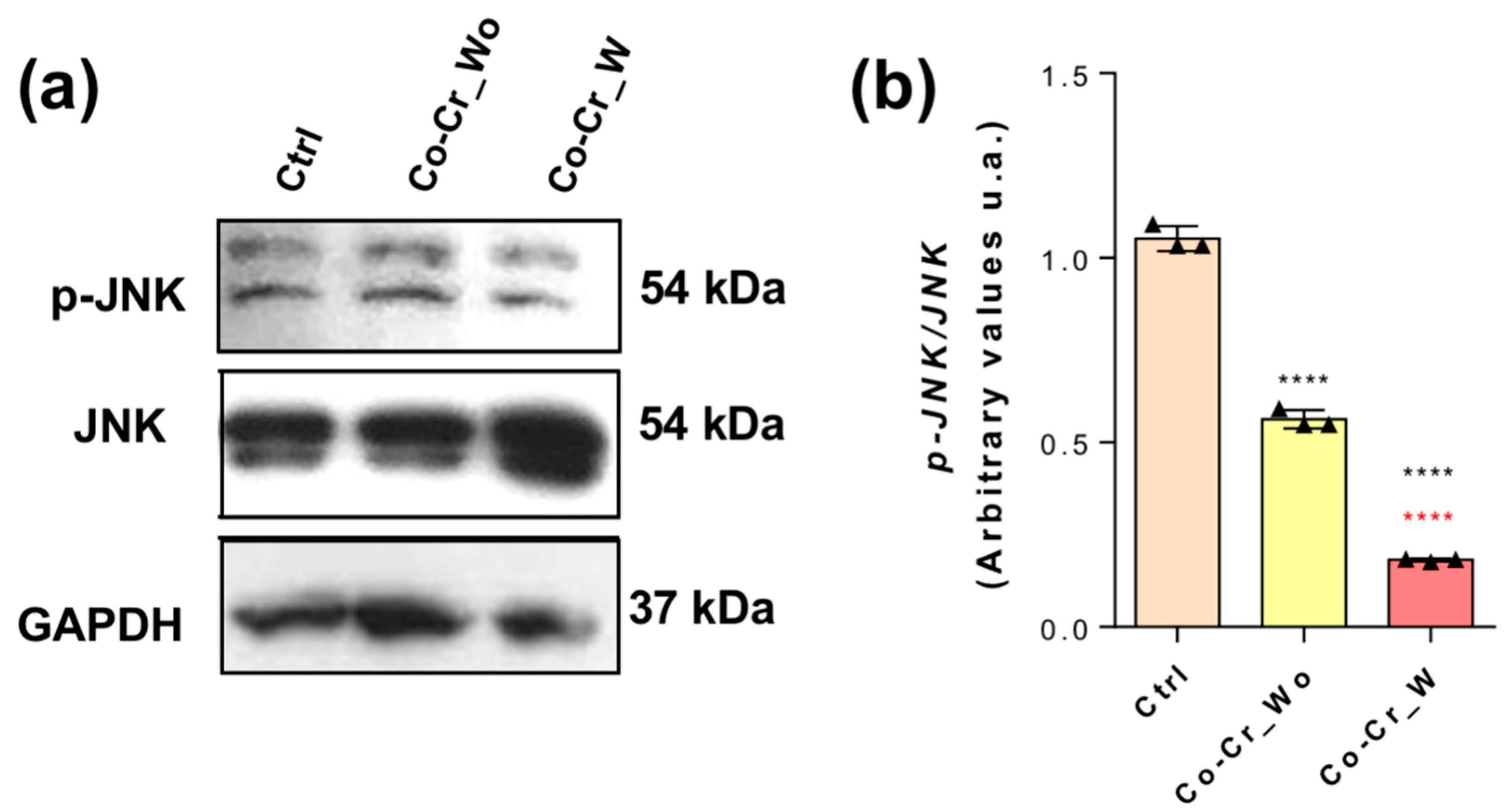
3.2. Effect of Co-Cr-Enriched Medium on JNK Phosphorylation
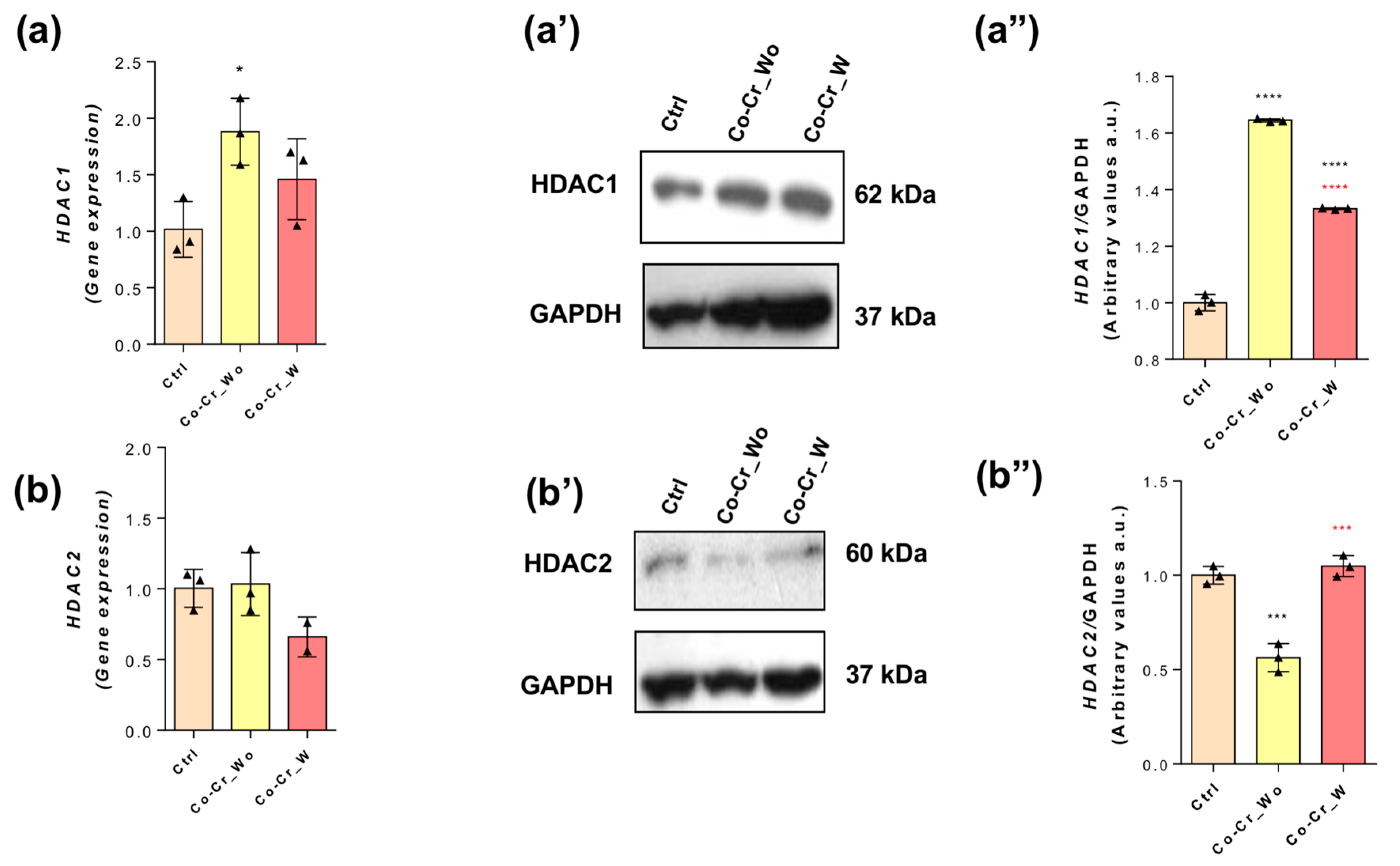
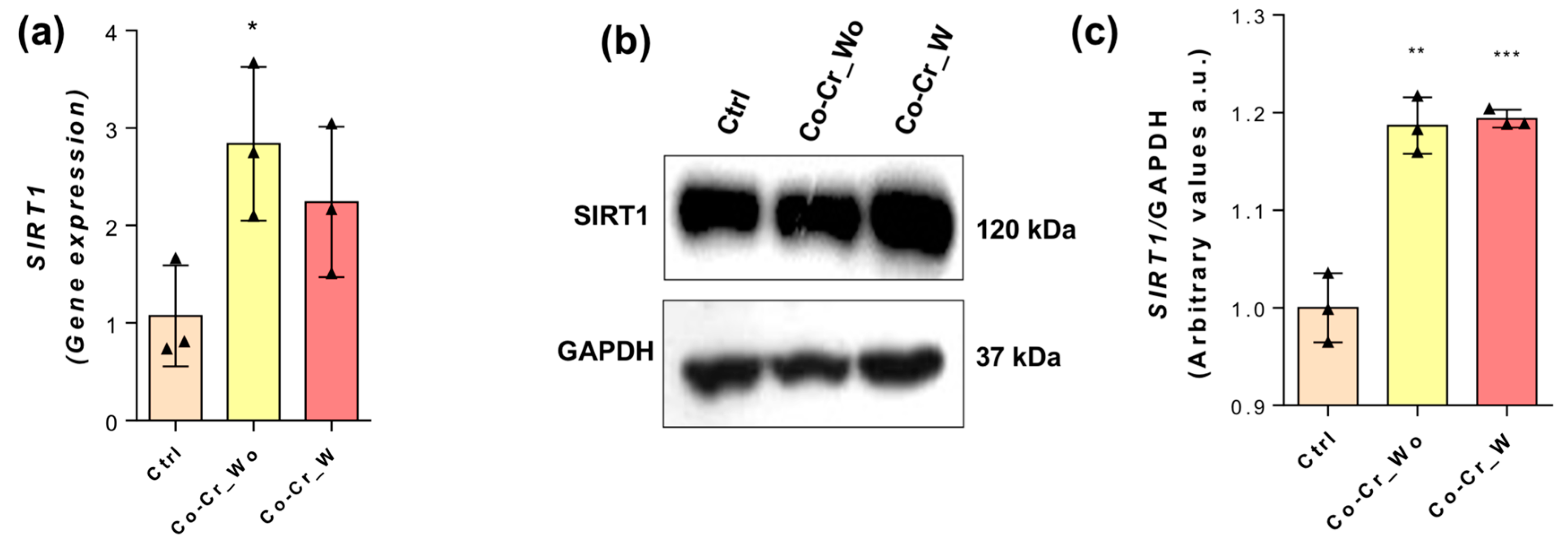
3.3. Effect of Co-Cr-Enriched Medium on the Protein Content of Histone Deacetylase Enzymes
3.4. DNA Methylation-Related DNMTs Were Investigated in Response to Co-Cr Alloys
3.5. DNA Demethylation-Related TETs Were Investigated in Response to Co-Cr Alloys
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdulghani, S.; Mitchell, G.R. Biomaterials for in Situ Tissue Regeneration: A Review. Biomolecules 2019, 9, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, E.; Boschetto, F.; Pezzotti, G. Biomaterials and Biocompatibility: An Historical Overview. J. Biomed. Mater. Res. Part A 2020, 108, 1617–1633. [Google Scholar] [CrossRef] [PubMed]
- Urban, I.A.; Monje, A. Guided Bone Regeneration in Alveolar Bone Reconstruction. Oral Maxillofac. Surg. Clin. N. Am. 2019, 31, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Bernardi, S.; Mattei, A.; Cristiano, L.; Mancini, L.; Torge, D.; Varvara, G.; Macchiarelli, G.; Marchetti, E. Morphological and Biological Evaluations of Human Periodontal Ligament Fibroblasts in Contact with Different Bovine Bone Grafts Treated with Low-Temperature Deproteinisation Protocol. Int. J. Mol. Sci. 2022, 23, 5273. [Google Scholar] [CrossRef]
- Accorsi-Mendonça, T.; Zambuzzi, W.F.; Paiva, K.B.D.S.; Lauris, J.R.P.; Cestari, T.M.; Taga, R.; Granjeiro, J.M. Expression of Metalloproteinase 2 in the Cell Response to Porous Demineralized Bovine Bone Matrix. J. Mol. Histol. 2005, 36, 311–316. [Google Scholar] [CrossRef]
- Zambuzzi, W.F.; de Oliveira, R.C.; Alanis, D.; Menezes, R.; Letra, A.; Cestari, T.M.; Taga, R.; Granjeiro, J.M. Microscopic Analisys of Porous Microgranular Bovine Anorganic Bone Implanted in Rat Subcutaneous Tissue. J. Appl. Oral Sci. 2005, 13, 382–386. [Google Scholar] [CrossRef]
- Zambuzzi, W.F.; Ferreira, C.V.; Granjeiro, J.M.; Aoyama, H. Biological Behavior of Pre-Osteoblasts on Natural Hydroxyapatite: A Study of Signaling Molecules from Attachment to Differentiation. J. Biomed. Mater. Res. A 2011, 97A, 193–200. [Google Scholar] [CrossRef]
- Bakhshandeh, B.; Zarrintaj, P.; Oftadeh, M.O.; Keramati, F.; Fouladiha, H.; Sohrabi-Jahromi, S.; Ziraksaz, Z. Tissue Engineering; Strategies, Tissues, and Biomaterials. Biotechnol. Genet. Eng. Rev. 2017, 33, 144–172. [Google Scholar] [CrossRef]
- Zor, F.; Selek, F.N.; Orlando, G.; Williams, D.F. Biocompatibility in Regenerative Nanomedicine. Nanomedicine 2019, 14, 2763–2775. [Google Scholar] [CrossRef]
- Brånemark, P.I.; Hansson, B.O.; Adell, R.; Breine, U.; Lindström, J.; Hallén, O.; Ohman, A. Osseointegrated Implants in the Treatment of the Edentulous Jaw. Experience from a 10-Year Period. Scand. J. Plast. Reconstr. Surg. Suppl. 1977, 16, 1–132. [Google Scholar]
- Salgado, A.J.; Coutinho, O.P.; Reis, R.L. Bone Tissue Engineering: State of the Art and Future Trends. Macromol. Biosci. 2004, 4, 743–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guehennec, L.; Soueidan, A.; Layrolle, P.; Amouriq, Y.; Le Guéhennec, L.; Soueidan, A.; Layrolle, P.; Amouriq, Y. Surface Treatments of Titanium Dental Implants for Rapid Osseointegration. Dent. Mater. Publ. Acad. Dent. Mat. 2007, 23, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.R.; da Costa Fernandes, C.J.; da Silva, R.A.; Constantino, V.R.L.; Koh, I.H.J.; Zambuzzi, W.F. Mg–Al and Zn–Al Layered Double Hydroxides Promote Dynamic Expression of Marker Genes in Osteogenic Differentiation by Modulating Mitogen-Activated Protein Kinases. Adv. Healthc. Mater. 2018, 7, 1700693. [Google Scholar] [CrossRef] [PubMed]
- Pinto, T.S.; Martins, B.R.; Ferreira, M.R.; Bezerra, F.; Zambuzzi, W.F. Nanohydroxyapatite-Blasted Bioactive Surface Drives Shear-Stressed Endothelial Cell Growth and Angiogenesis. BioMed Res. Int. 2022, 2022, 1433221. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Milani, R.; Rangel, E.C.; Peppelenbosch, M.; Zambuzzi, W. OsteoBLAST: Computational Routine of Global Molecular Analysis Applied to Biomaterials Development. Front. Bioeng. Biotechnol. 2020, 8, 565901. [Google Scholar] [CrossRef]
- Zambuzzi, W.F.; Bonfante, E.A.; Jimbo, R.; Hayashi, M.; Andersson, M.; Alves, G.; Takamori, E.R.; Beltrao, P.J.; Coelho, P.G.; Granjeiro, J.M. Nanometer Scale Titanium Surface Texturing Are Detected by Signaling Pathways Involving Transient FAK and Src Activations. PloS ONE 2014, 9, e95662. [Google Scholar] [CrossRef]
- Da Silva, R.A.; Fernandes, C.J.D.C.; Feltran, G.D.S.; Gomes, A.M.; de Camargo Andrade, A.F.; Andia, D.C.; Peppelenbosch, M.P.; Zambuzzi, W.F. Laminar Shear Stress-Provoked Cytoskeletal Changes Are Mediated by Epigenetic Reprogramming of TIMP1 in Human Primary Smooth Muscle Cells. J. Cell. Physiol. 2019, 234, 6382–6396. [Google Scholar] [CrossRef]
- Machado, M.I.P.; Gomes, A.M.; Rodrigues, M.F.; Silva Pinto, T.; da Costa Fernandes, C.J.; Bezerra, F.J.; Zambuzzi, W.F. Cobalt-Chromium-Enriched Medium Ameliorates Shear-Stressed Endothelial Cell Performance. J. Trace Elem. Med. Biol. 2019, 54, 163–171. [Google Scholar] [CrossRef]
- Rossi, M.C.; Bezerra, F.J.B.; Silva, R.A.; Crulhas, B.P.; Fernandes, C.J.C.; Nascimento, A.S.; Pedrosa, V.A.; Padilha, P.; Zambuzzi, W.F. Titanium-Released from Dental Implant Enhances Pre-Osteoblast Adhesion by ROS Modulating Crucial Intracellular Pathways. J. Biomed. Mater. Res. Part A 2017, 105, 2968–2976. [Google Scholar] [CrossRef]
- Martins, B.R.; Pinto, T.S.; da Costa Fernandes, C.J.; Bezerra, F.; Zambuzzi, W.F. PI3K/AKT Signaling Drives Titanium-Induced Angiogenic Stimulus. J. Mater. Sci. Mater. Med. 2021, 32, 18. [Google Scholar] [CrossRef]
- Bezerra, F.; Ferreira, M.R.; Fontes, G.N.; da Costa Fernandes, C.J.; Andia, D.C.; Cruz, N.C.; da Silva, R.A.; Zambuzzi, W.F. Nano Hydroxyapatite-Blasted Titanium Surface Affects Pre-Osteoblast Morphology by Modulating Critical Intracellular Pathways. Biotechnol. Bioeng. 2017, 114, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.J.C.; Bezerra, F.; Ferreira, M.R.; Andrade, A.F.C.; Pinto, T.S.; Zambuzzi, W.F. Nano Hydroxyapatite-Blasted Titanium Surface Creates a Biointerface Able to Govern Src-Dependent Osteoblast Metabolism as Prerequisite to ECM Remodeling. Colloids Surf. B Biointerfaces 2018, 163, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, C.J.C.; Bezerra, F.; do Carmo, M.D.D.; Feltran, G.S.; Rossi, M.C.; da Silva, R.A.; de M. Padilha, P.; Zambuzzi, W.F. CoCr-Enriched Medium Modulates Integrin-Based Downstream Signaling and Requires a Set of Inflammatory Genes Reprograming in Vitro. J. Biomed. Mater. Res. Part A 2018, 106, 839–849. [Google Scholar] [CrossRef]
- Da Costa Fernandes, C.J.; Ferreira, M.R.; Bezerra, F.J.B.; Zambuzzi, W.F. Zirconia Stimulates ECM-Remodeling as a Prerequisite to Pre-Osteoblast Adhesion/Proliferation by Possible Interference with Cellular Anchorage. J. Mater. Sci. Mater. Med. 2018, 29, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagatsuma, A.; Arakawa, M.; Matsumoto, H.; Matsuda, R.; Hoshino, T.; Mabuchi, K. Cobalt Chloride, a Chemical Hypoxia-Mimicking Agent, Suppresses Myoblast Differentiation by Downregulating Myogenin Expression. Mol. Cell. Biochem. 2020, 470, 199–214. [Google Scholar] [CrossRef]
- Howie, D.W.; Rogers, S.D.; McGee, M.A.; Haynes, D.R. Biologic Effects of Cobalt Chrome in Cell and Animal Models. Clin. Orthop. Relat. Res. 1996, 329, S217–S232. [Google Scholar] [CrossRef]
- Caicedo, M.S.; Pennekamp, P.H.; McAllister, K.; Jacobs, J.J.; Hallab, N.J. Soluble Ions More than Particulate Cobalt-Alloy Implant Debris Induce Monocyte Costimulatory Molecule Expression and Release of Proinflammatory Cytokines Critical to Metal-Induced Lymphocyte Reactivity. J. Biomed. Mater. Res. Part A 2010, 93, 1312–1321. [Google Scholar] [CrossRef]
- Celesti, C.; Gervasi, T.; Cicero, N.; Giofrè, S.V.; Espro, C.; Piperopoulos, E.; Gabriele, B.; Mancuso, R.; Lo Vecchio, G.; Iannazzo, D. Titanium Surface Modification for Implantable Medical Devices with Anti-Bacterial Adhesion Properties. Materials 2022, 15, 3283. [Google Scholar] [CrossRef]
- Al-Noaman, A.; Rawlinson, S.C.F.; Hill, R.G. MgF2- Containing Glasses as a Coating for Titanium Dental Implant. I- Glass Powder. J. Mech. Behav. Biomed. Mater. 2022, 125, 104948. [Google Scholar] [CrossRef]
- Yuan, Y.; Luo, R.; Ren, J.; Zhang, L.; Jiang, Y.; He, Z. Design of a New Ti-Mo-Cu Alloy with Excellent Mechanical and Antibacterial Properties as Implant Materials. Mater. Lett. 2022, 306, 130875. [Google Scholar] [CrossRef]
- Celesti, C.; Iannazzo, D.; Espro, C.; Visco, A.; Legnani, L.; Veltri, L.; Visalli, G.; Di Pietro, A.; Bottino, P.; Chiacchio, M.A. Chitosan/POSS Hybrid Hydrogels for Bone Tissue Engineering. Materials 2022, 15, 8208. [Google Scholar] [CrossRef]
- Da Silva, R.A.; Fuhler, G.M.; Janmaat, V.T.; da C. Fernandes, C.J.; da Silva Feltran, G.; Oliveira, F.A.; Matos, A.A.; Oliveira, R.C.; Ferreira, M.R.; Zambuzzi, W.F.; et al. HOXA Cluster Gene Expression during Osteoblast Differentiation Involves Epigenetic Control. Bone 2019, 125, 74–86. [Google Scholar] [CrossRef]
- Célio, C.J.; da Silva, R.A.; Fretes Wood, P.; Teixeira, S.A.; Bezerra, F.; Zambuzzi, W.F. The Molecular Pathway Triggered by Zirconia in Endothelial Cells Involves Epigenetic Control. Tissue Cell 2021, 73, 101627. [Google Scholar] [CrossRef]
- Fitz-James, M.H.; Cavalli, G. Molecular Mechanisms of Transgenerational Epigenetic Inheritance. Nat. Rev. Genet. 2022, 23, 325–341. [Google Scholar] [CrossRef]
- Delcuve, G.P.; Rastegar, M.; Davie, J.R. Epigenetic Control. J. Cell. Physiol. 2009, 219, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Lappalainen, T.; Greally, J.M. Associating Cellular Epigenetic Models with Human Phenotypes. Nat. Rev. Genet. 2017, 18, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Prachayasittikul, V.; Prathipati, P.; Pratiwi, R.; Phanus-Umporn, C.; Malik, A.A.; Schaduangrat, N.; Seenprachawong, K.; Wongchitrat, P.; Supokawej, A.; Prachayasittikul, V.; et al. Exploring the Epigenetic Drug Discovery Landscape. Expert Opin. Drug Discov. 2017, 12, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Tsumagari, K.; Baribault, C.; Terragni, J.; Chandra, S.; Renshaw, C.; Sun, Z.; Song, L.; Crawford, G.E.; Pradhan, S.; Lacey, M.; et al. DNA Methylation and Differentiation: HOX Genes in Muscle Cells. Epigenetics Chromatin 2013, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- De Souza Queiroz, K.C.; Zambuzzi, W.F.; Santos de Souza, A.C.; da Silva, R.A.; Machado, D.; Justo, G.Z.; Carvalho, H.F.; Peppelenbosch, M.P.; Ferreira, C.V. A Possible Anti-Proliferative and Anti-Metastatic Effect of Irradiated Riboflavin in Solid Tumours. Cancer Lett. 2007, 258, 126–134. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Ferreira, R.S.; Assis, R.I.F.; Feltran, G.D.S.; do Rosário Palma, I.C.; Françoso, B.G.; Zambuzzi, W.F.; Andia, D.C.; da Silva, R.A. Genome-Wide DNA (Hydroxy) Methylation Reveals the Individual Epigenetic Landscape Importance on Osteogenic Phenotype Acquisition in Periodontal Ligament Cells. J. Periodontol. 2022, 93, 435–448. [Google Scholar] [CrossRef]
- Da Silva, R.A.; Ferreira, M.R.; Gomes, A.M.; Zambuzzi, W.F. LncRNA HOTAIR Is a Novel Endothelial Mechanosensitive Gene. J. Cell. Physiol. 2020, 235, 4631–4642. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long Non-Coding RNA HOTAIR Reprograms Chromatin State to Promote Cancer Metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Razzouk, S.; Sarkis, R. Smoking and Diabetes. Epigenetics Involvement in Osseointegration. New York State Dent. J. 2013, 79, 27–30. [Google Scholar]
- Uysal, F.; Ozturk, S.; Akkoyunlu, G. DNMT1, DNMT3A and DNMT3B Proteins Are Differently Expressed in Mouse Oocytes and Early Embryos. J. Mol. Histol. 2017, 48, 417–426. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.A.; da S. Feltran, G.; da C. Fernandes, C.J.; Zambuzzi, W.F. Osteogenic gene markers are epigenetically reprogrammed during contractile-to-calcifying vascular smooth muscle cell phenotype transition. Cell Signal 2020, 66, 109458. [Google Scholar] [CrossRef]
- Loo, S.K.; Ab Hamid, S.S.; Musa, M.; Wong, K.K. DNMT1 Is Associated with Cell Cycle and DNA Replication Gene Sets in Diffuse Large B-Cell Lymphoma. Pathol. Res. Pract. 2018, 214, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Svedruzic, Z.M.; Svedružić, Ž.M. Dnmt1 Structure and Function. Prog. Mol. Biol. Transl. Sci. 2011, 101, 221–254. [Google Scholar] [CrossRef]
- Szyf, M. The Role of DNA Methyltransferase 1 in Growth Control. Front. Biosci. A J. Virtual Libr. 2001, 6, D599–D609. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F. The DNA Methyltransferase Family: A Versatile Toolkit for Epigenetic Regulation. Nat. Rev. Genet. 2018, 19, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-X.; He, X.-Q.; Li, F.-X.; Wu, Y.-S.; Gao, Y.; Li, M. Risk-Association of DNA Methyltransferases Polymorphisms with Gastric Cancer in the Southern Chinese Population. Int. J. Mol. Sci. 2012, 13, 8364–8378. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The Use of Cobalt Chloride as a Chemical Hypoxia Model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Keusch, J.J.; Wang, L.; Saito, M.; Hess, D.; Wang, X.; Melancon, B.J.; Helquist, P.; Gut, H.; Matthias, P. Structural Insights into HDAC6 Tubulin Deacetylation and Its Selective Inhibition. Nat. Chem. Biol. 2016, 12, 748–754. [Google Scholar] [CrossRef]
- Li, T.; Zhang, C.; Hassan, S.; Liu, X.; Song, F.; Chen, K.; Zhang, W.; Yang, J. Histone Deacetylase 6 in Cancer. J. Hematol. Oncol. 2018, 11, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaluza, D.; Kroll, J.; Gesierich, S.; Yao, T.-P.; Boon, R.A.; Hergenreider, E.; Tjwa, M.; Rossig, L.; Seto, E.; Augustin, H.G.; et al. Class IIb HDAC6 Regulates Endothelial Cell Migration and Angiogenesis by Deacetylation of Cortactin. EMBO J. 2011, 30, 4142–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kästle, M.; Woschee, E.; Grune, T. Histone Deacetylase 6 (HDAC6) Plays a Crucial Role in P38MAPK-Dependent Induction of Heme Oxygenase-1 (HO-1) in Response to Proteasome Inhibition. Free. Radic. Biol. Med. 2012, 53, 2092–2101. [Google Scholar] [CrossRef]
- Zambuzzi, W.F.; Bruni-Cardoso, A.; Granjeiro, J.M.; Peppelenbosch, M.P.; De Carvalho, H.F.; Aoyama, H.; Ferreira, C.V. On the Road to Understanding of the Osteoblast Adhesion: Cytoskeleton Organization Is Rearranged by Distinct Signaling Pathways. J. Cell. Biochem. 2009, 108, 134–144. Available online: https://pubmed.ncbi.nlm.nih.gov/19562668/ (accessed on 8 February 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (ID) | Primer | 5′–3′ Sequence | Reactions Condition |
|---|---|---|---|
| HDAC1 (3065) | Forward | CTGGCCATCATCTCCTTGAT | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | ACCAGAGACGTGGAAACTGG | ||
| HDAC2 (3066) | Forward | TTCTCAGTGCACCCAGTCAG | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | CCAGTATCCTTGGGGGAAAT | ||
| HDAC3 (8841) | Forward | ACGTGGGCAACTTCCACTAC | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | GACTCTTGGTGAAGCCTTGC | ||
| HDAC6 (10013) | Forward | AAGTAGGCAGAACCCCCAGT | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | GTGCTTCAGCCTCAAGGTTC | ||
| SIRT1 (23411) | Forward | GCAGATTAGTAGGCGGCTTG | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | TCTGGCATGTCCCACTATCA | ||
| DNMT1 (1786) | Forward | AGGACCCAGACAGAGAAGCA | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | GTACGGGAATGCTGAGTGGT | ||
| DNMT3A (13435) | Forward | AGGAAGCCCATCCGGGTGCTA | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | AGCGGTCCACTTGGATGCCC | ||
| DNMT3B (1789) | Forward | TCGACTTGGTGGTTATTGTCTG | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | TCGAGCTACAAGACTGCTTGG | ||
| TET1 (80312) | Forward | GCCCCTCTTCATTACCAAGTC | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | CGCCAGTTGCTTATCAAAATC | ||
| TET2 (54790) | Forward | GGTGCCTCTGGAGTGACTGT | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | GGAAAATGCAAGCCCTATGA | ||
| TET3 (200424) | Forward | GGTCAGGCTGGTTTACAACG | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | GGCATAGACCCACACACATCT | ||
| GAPDH (2597) | Forward | AAGGTGAAGGTCGGAGTCAA | 95 °C-15 s; 58 °C-30 s; 72 °C-60 s |
| Reverse | AATGAAGGGGTCATTGATGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da C. Fernandes, C.J.; da Silva, R.A.F.; de Almeida, G.S.; Ferreira, M.R.; de Morais, P.B.; Bezerra, F.; Zambuzzi, W.F. Epigenetic Differences Arise in Endothelial Cells Responding to Cobalt–Chromium. J. Funct. Biomater. 2023, 14, 127. https://doi.org/10.3390/jfb14030127
da C. Fernandes CJ, da Silva RAF, de Almeida GS, Ferreira MR, de Morais PB, Bezerra F, Zambuzzi WF. Epigenetic Differences Arise in Endothelial Cells Responding to Cobalt–Chromium. Journal of Functional Biomaterials. 2023; 14(3):127. https://doi.org/10.3390/jfb14030127
Chicago/Turabian Styleda C. Fernandes, Célio Junior, Rodrigo A. Foganholi da Silva, Gerson Santos de Almeida, Marcel Rodrigues Ferreira, Paula Bertin de Morais, Fábio Bezerra, and Willian F. Zambuzzi. 2023. "Epigenetic Differences Arise in Endothelial Cells Responding to Cobalt–Chromium" Journal of Functional Biomaterials 14, no. 3: 127. https://doi.org/10.3390/jfb14030127
APA Styleda C. Fernandes, C. J., da Silva, R. A. F., de Almeida, G. S., Ferreira, M. R., de Morais, P. B., Bezerra, F., & Zambuzzi, W. F. (2023). Epigenetic Differences Arise in Endothelial Cells Responding to Cobalt–Chromium. Journal of Functional Biomaterials, 14(3), 127. https://doi.org/10.3390/jfb14030127