Carbon Nanoparticles Functionalized with Carboxylic Acid Improved the Germination and Seedling Vigor in Upland Boreal Forest Species

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Mechanical Scarification + Stratification + CNPs + Gibberellic Acid (GA)
2.2. Chemical Scarification + Stratification + CNPs + GA
2.3. Tetrazolium Test
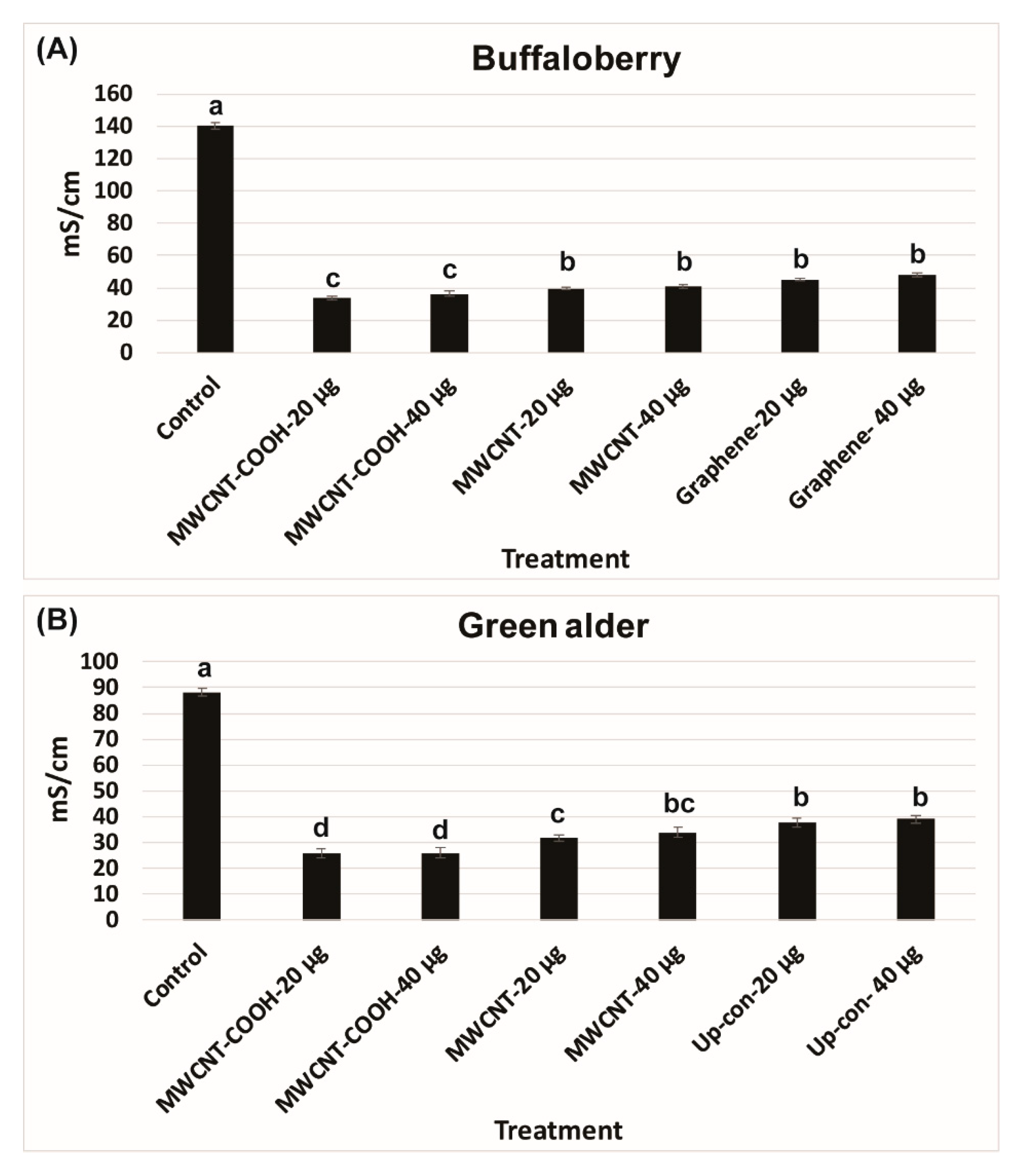
2.4. Electrical Conductivity Test
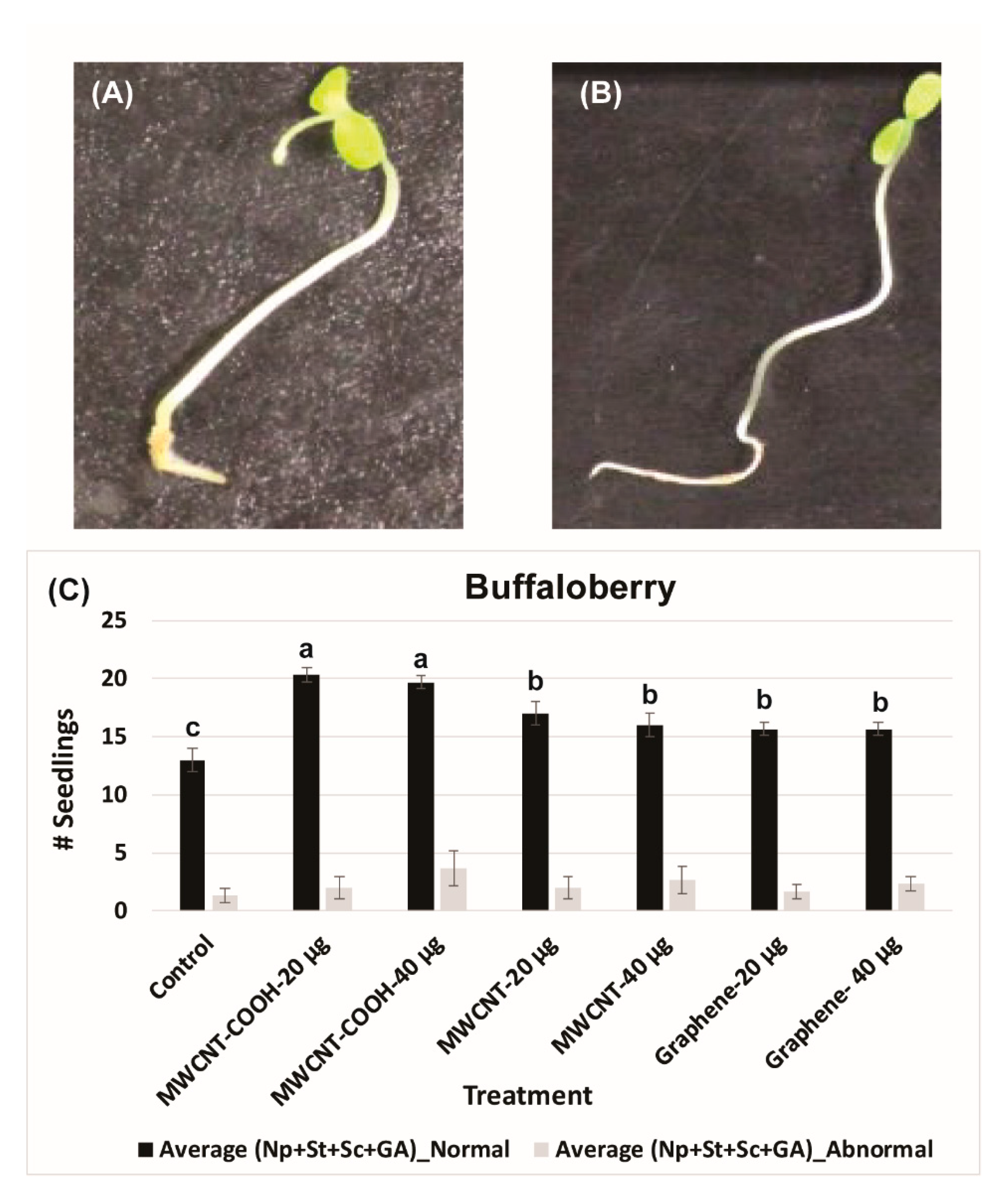
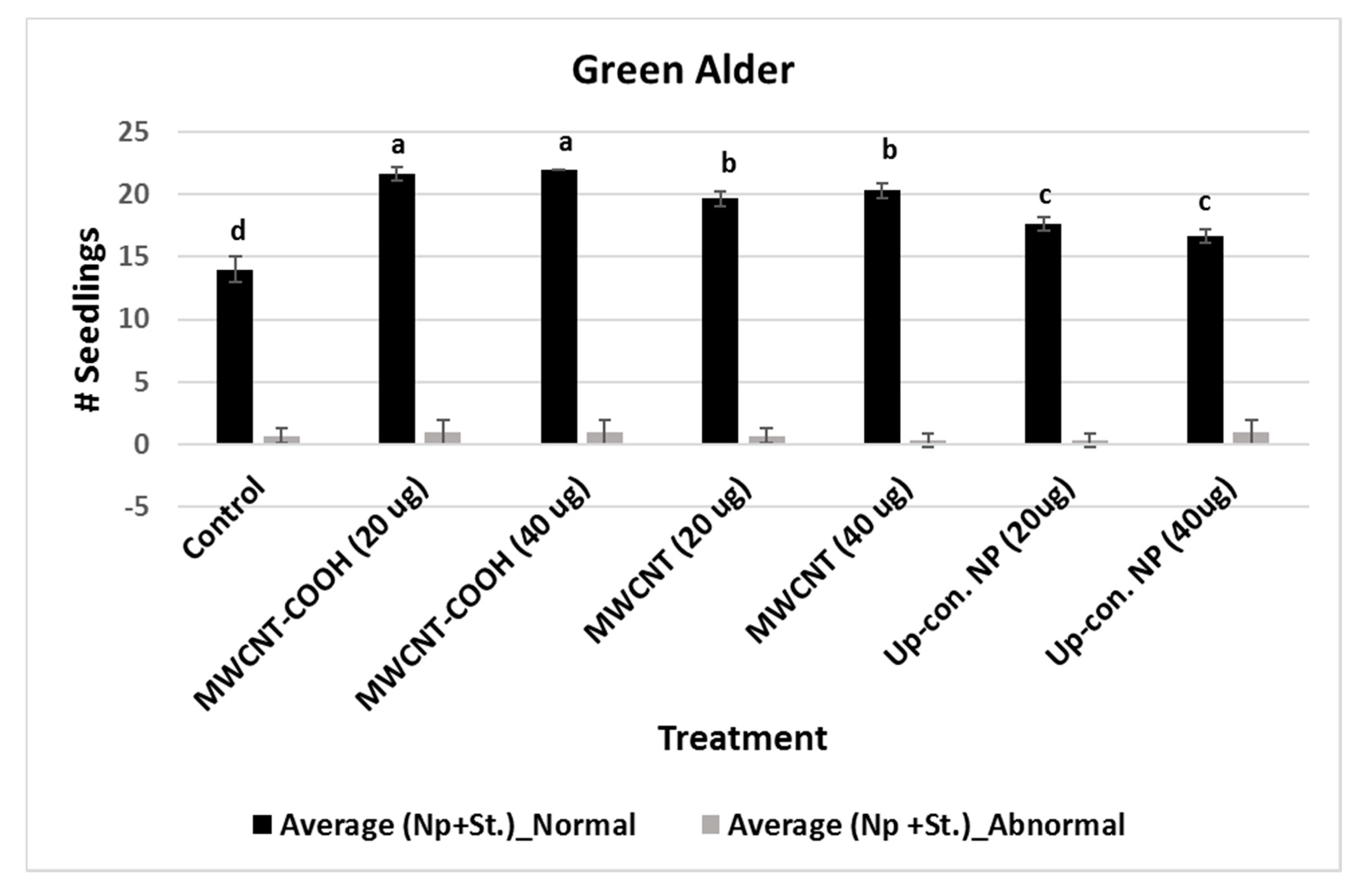
2.5. Evaluation of Normal and Abnormal Seedlings
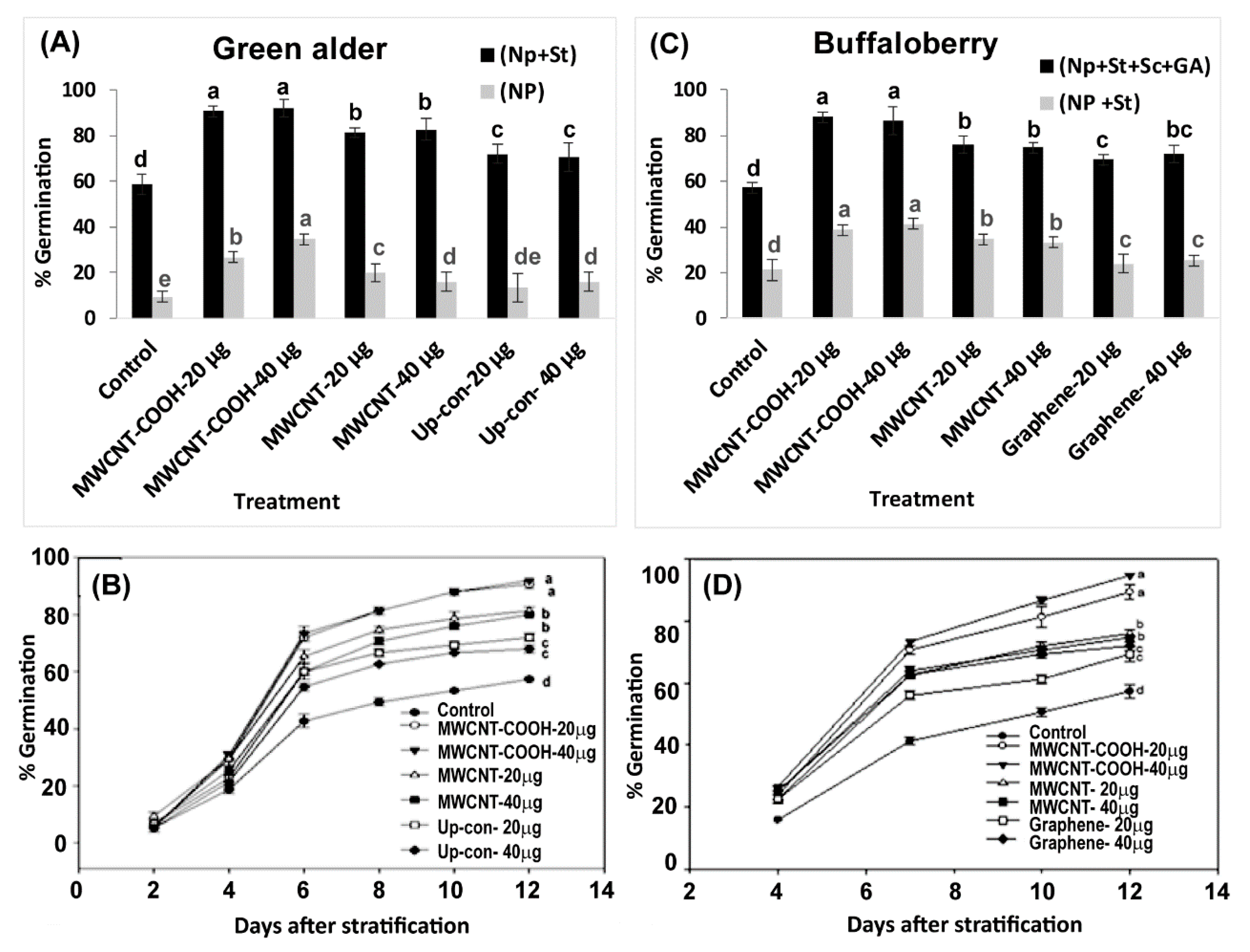
2.6. Germination Percentages
2.7. Seedling Vigor Index (SVI)
2.8. Lipid Extraction and Analyses
2.8.1. Chemicals
2.8.2. Lipid Extraction
2.9. Plant Membrane Lipids Analysis Using UHPLC-C30RP-HESI-HRMS/MS
2.10. Data Processing
3. Results
3.1. Seed and Seedling Quality Analysis
3.2. Effects of Nanopriming on Resolving Seed Dormancy
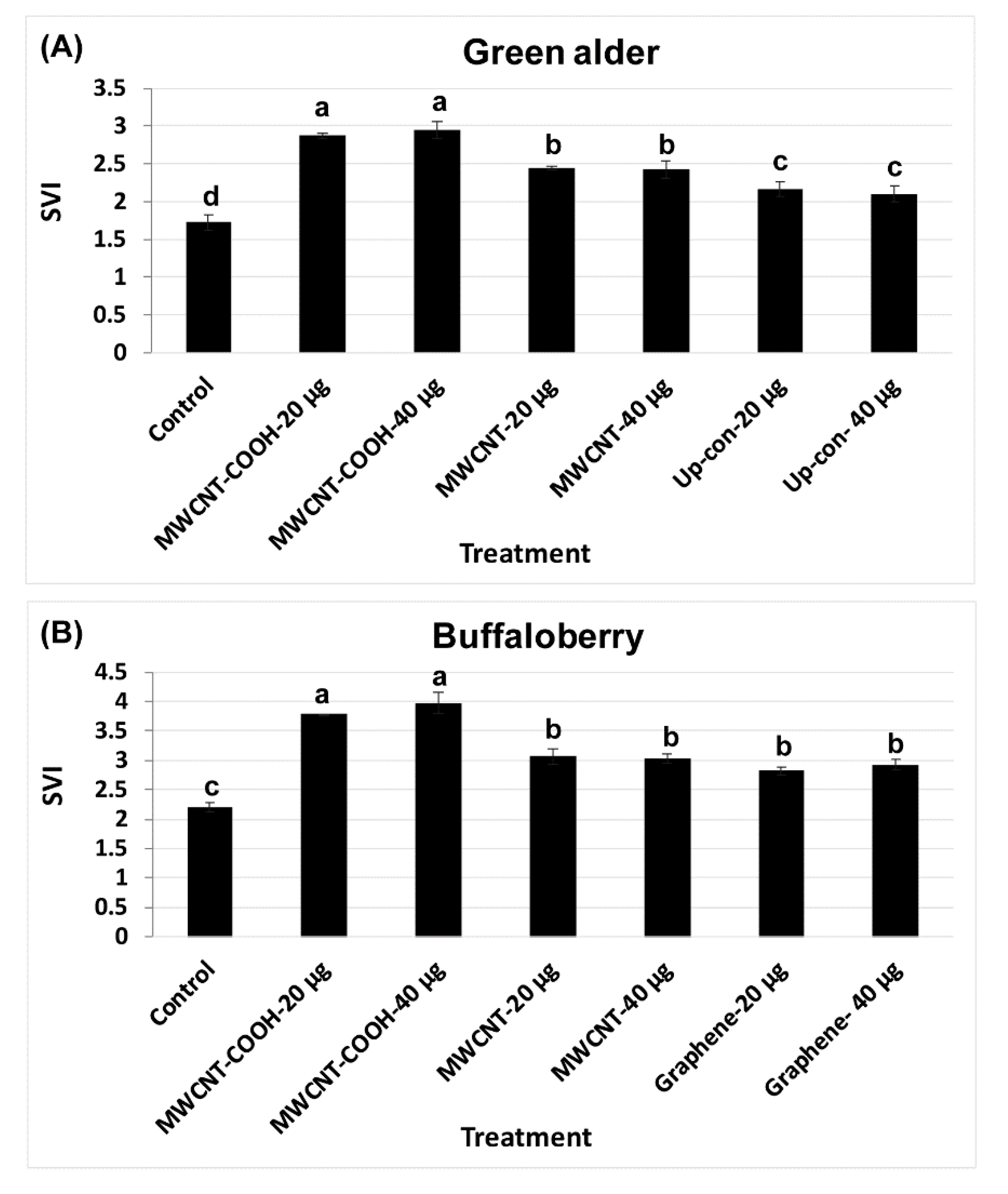
3.3. Seedling Vigor Index
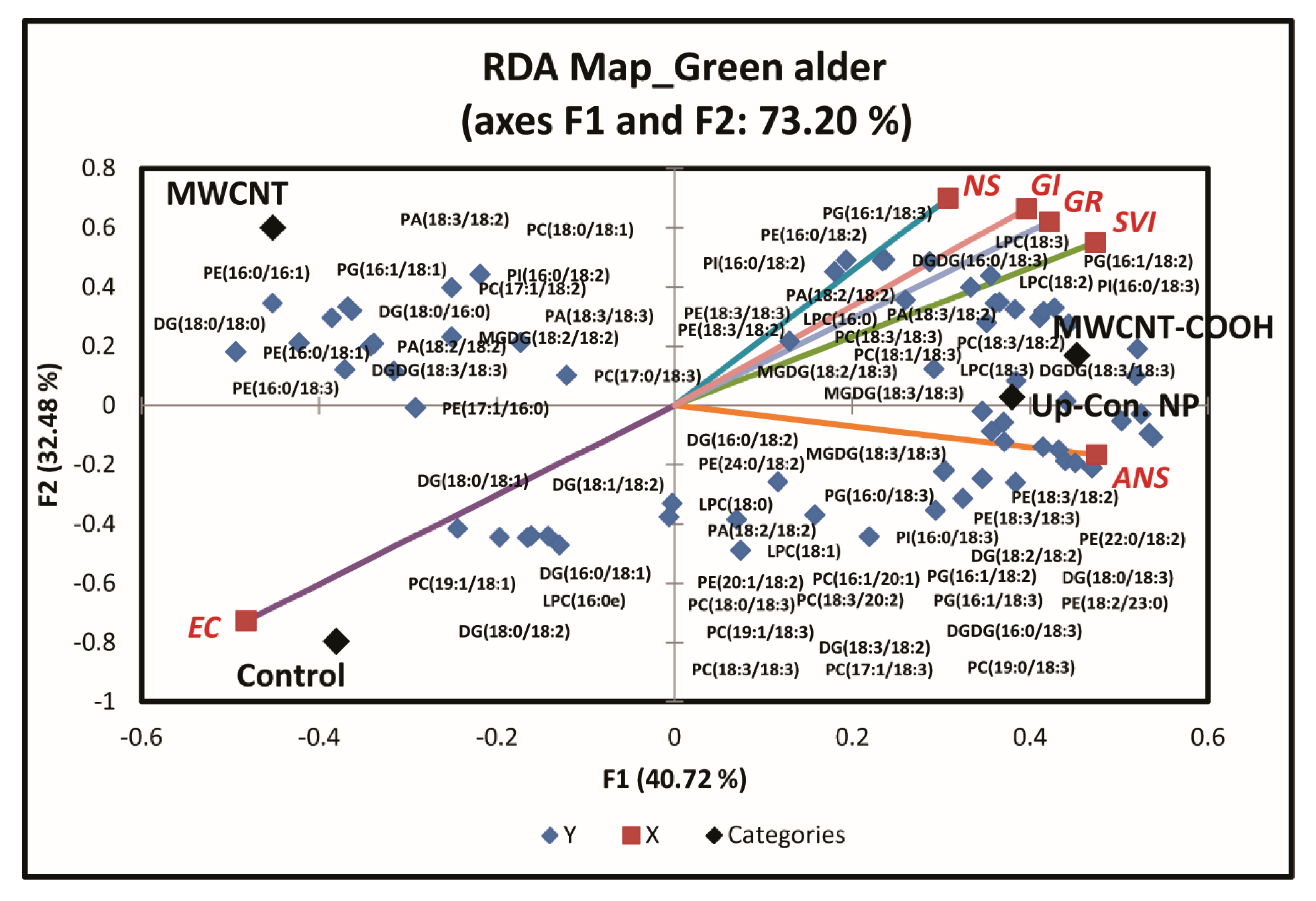
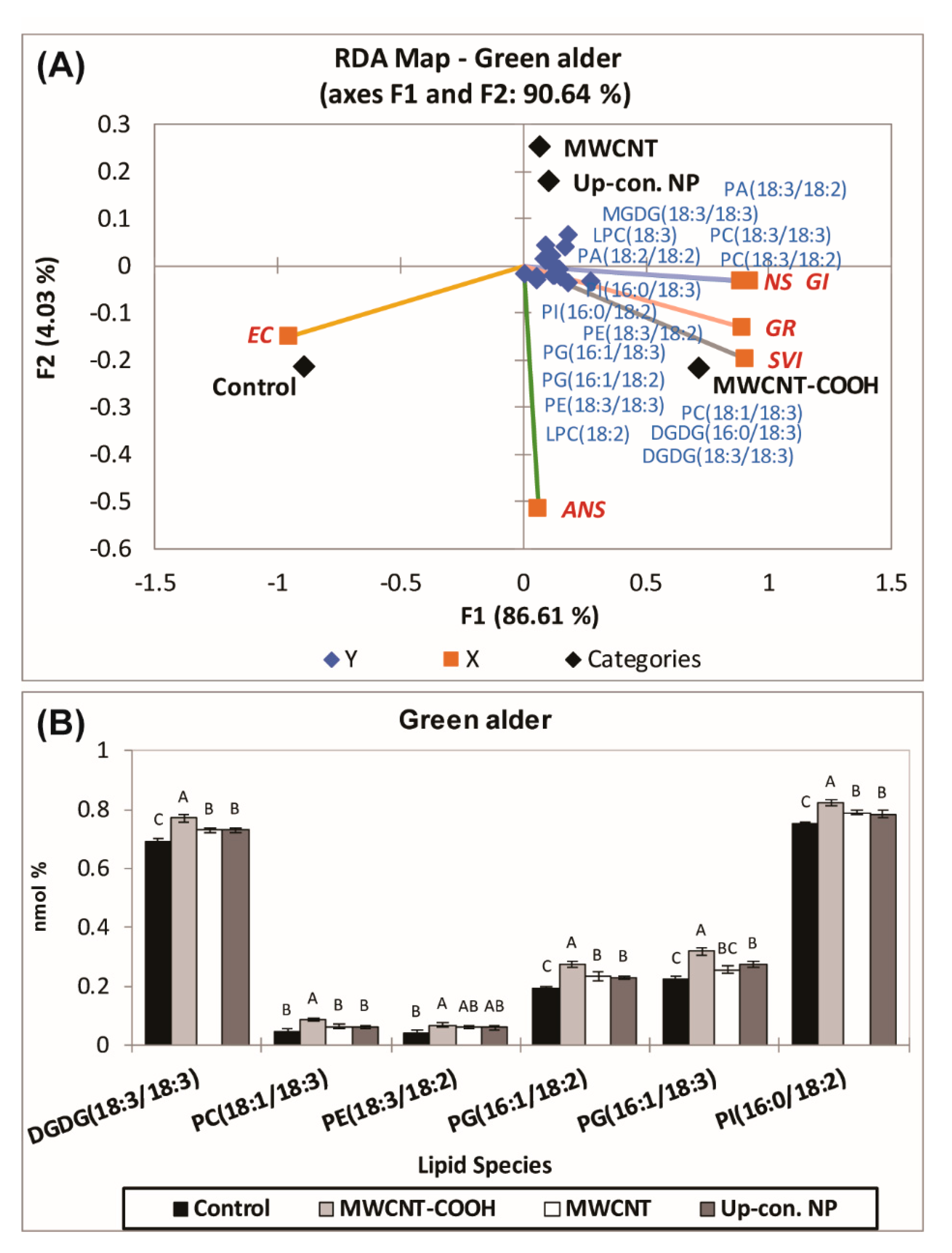
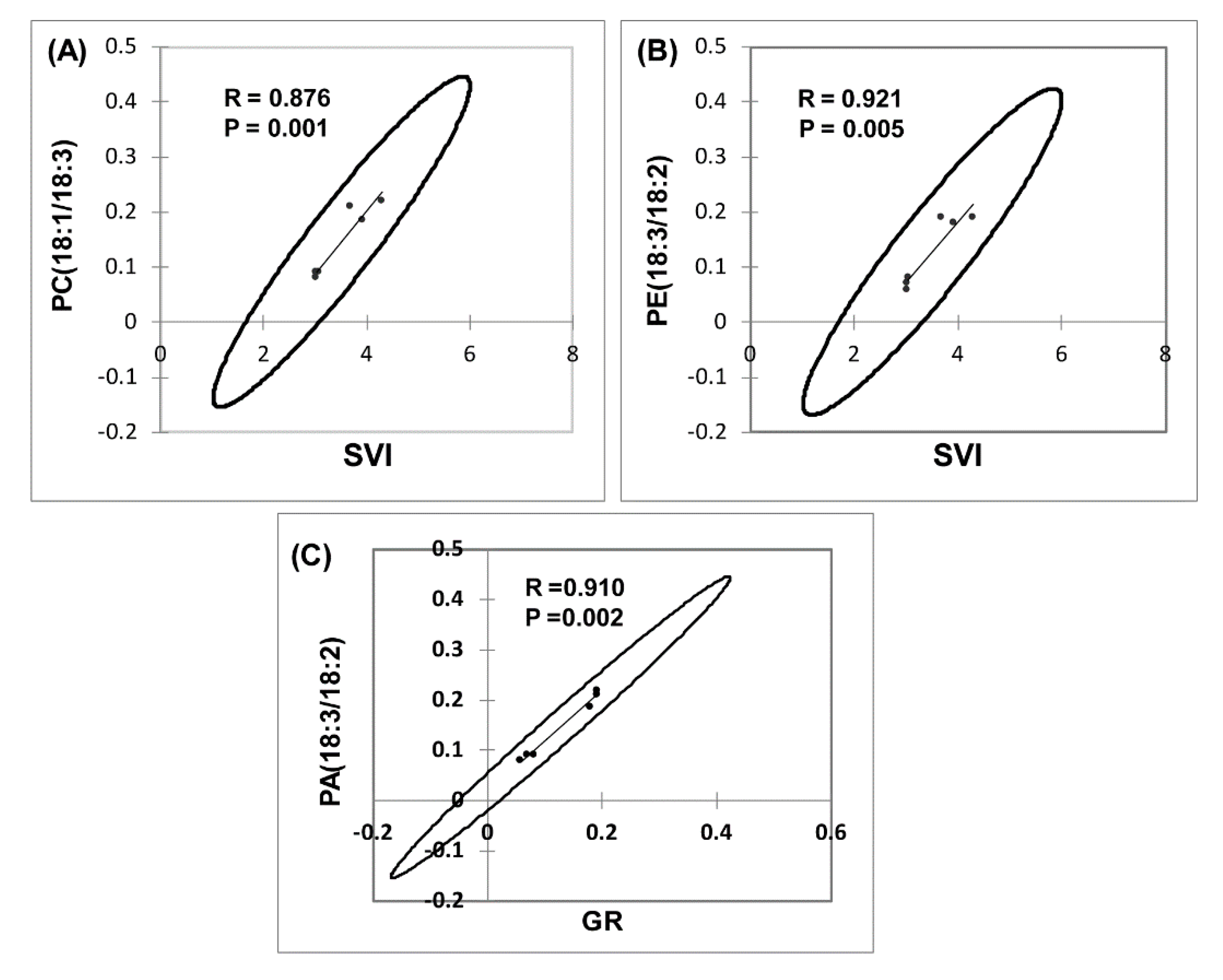
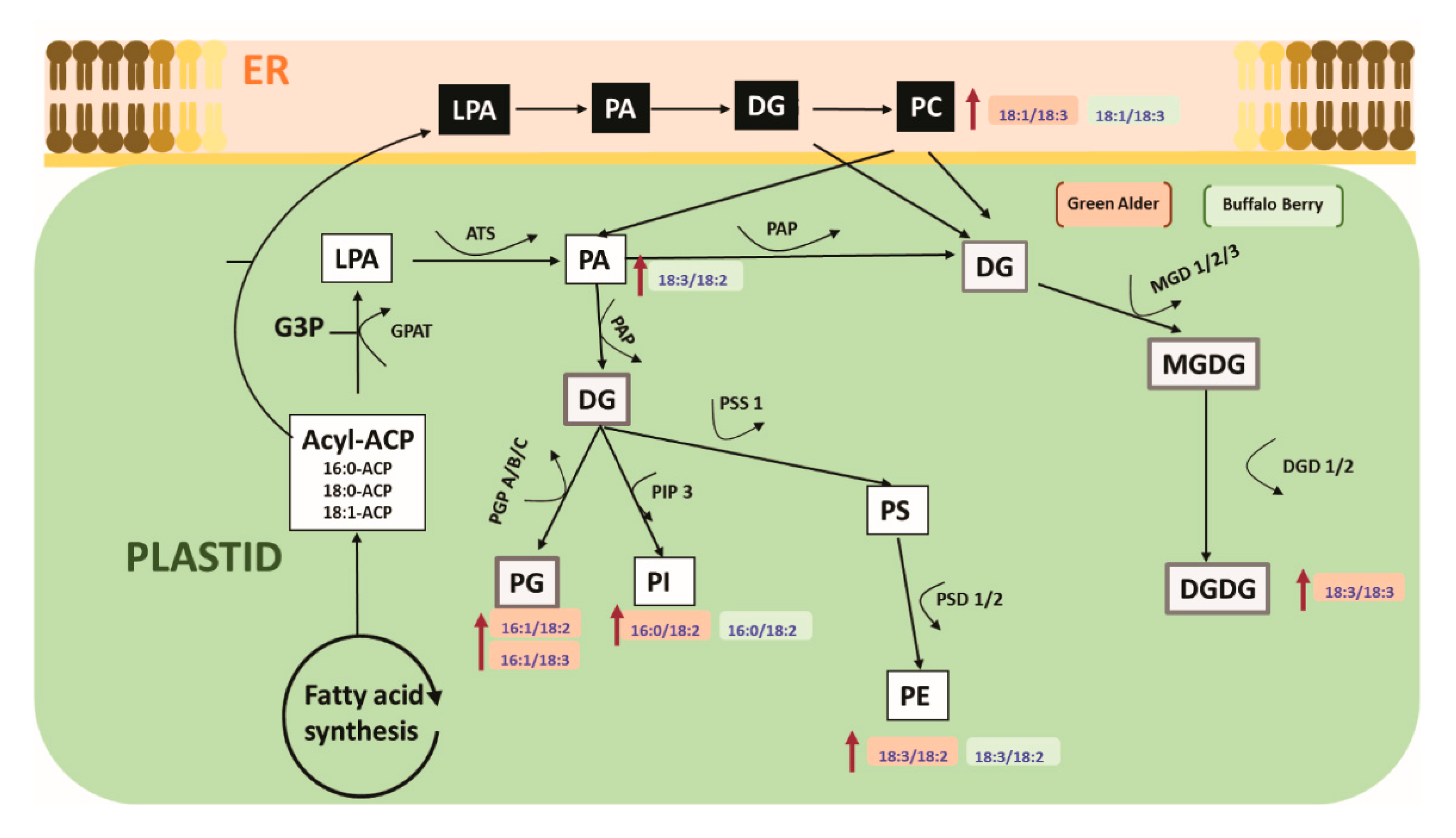
3.4. Roles of Lipid Metabolism in Overcoming Seed Dormancy in Upland Boreal Forest Species
4. Discussion
4.1. Effect of Carbon Nanoparticles on Seedling Quality
4.2. Effects of Carbon Nanoparticles in Resolving Seed Dormancy in Upland Boreal Forest Species
4.3. Effect of Carbon Nanoparticles in Enhancing Seedling Vigor
4.4. Role of Lipid Metabolism in Resolving Seed Dormancy in Upland Boreal Forest Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Optimizing Plant Population: Crop Emergence and Establishment; FAO: Rome, Italy, 2011. [Google Scholar]
- Lechowska, K.; Kubala, S.; Wojtyla, Ł.; Nowaczyk, G.; Quinet, M.; Lutts, S.; Garnczarska, M. New insight on water status in germinating brassica napus seeds in relation to priming-improved germination. Int. J. Mol. Sci. 2019, 20, 540. [Google Scholar] [CrossRef] [Green Version]
- Weller, S.L.; Florentine, S.K.; Chauhan, B.S. Influence of selected environmental factors on seed germination and seedling emergence of Dinebra panicea var. brachiata (Steud.). Crop Prot. 2019, 117, 121–127. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2007, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Nabeta, K.; Matsuura, H. Chemical inhibitors of viviparous germination in the fruit of watermelon. Plant Cell Physiol. 2010, 51, 1594–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.H.; Franklin, S.E.; Guo, X.L.; Stenhouse, G.B. Narrow-linear and small-area forest disturbance detection and mapping from high spatial resolution imagery. J. Appl. Remote Sens. 2009, 3, 033570. [Google Scholar] [CrossRef]
- De Bortoli, L.A.; Pinno, B.D.; MacKenzie, M.D.; Li, E.H.Y. Plant community composition and tree seedling establishment in response to seeding and weeding treatments on different reclamation cover soils. Can. J. For. Res. 2019, 49, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Margenau, E.L.; Wood, P.B.; Weakland, C.A.; Brown, D.J. Trade-offs relating to grassland and forest mine reclamation approaches in the central Appalachian region and implications for the songbird community. Avian Conserv. Ecol. 2019, 14. [Google Scholar] [CrossRef]
- Rosner, L.S.; Harrington, J.T. Russet buffaloberry seeds. Native 2003, Fall, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Beddes, T.; Kratsch, H.A. Seed germination of roundleaf buffaloberry (Shepherdia rotundifolia) and silver buffaloberry (Shepherdia argentea) in three substrates. J. Environ. Hortic. 2009, 27, 129–133. [Google Scholar] [CrossRef]
- Vogel, J.G.; Gower, S.T. Carbon and nitrogen dynamics of boreal jack pine stands with and without a green alder understory. Ecosystems 1998, 1, 386–400. [Google Scholar] [CrossRef]
- Kaur, J.; Schoonmaker, A.; Sobze, J.-M. Length of cold stratification period affects germination in green alder (Alnus viridis (Chaix) DC. subsp. crispa (Aiton) Turrill) seed collected from northwestern Alberta. Nativ. Plants J. 2016, 17, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. In Arabidopsis Book, 1st ed.; BioOne: Washington, DC, USA, 2008; Volume 6, p. 119. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Bose, B. Impact of micronutrient seed priming on germination, growth, development, nutritional status and yield aspects of plants. J. Plant Nutr. 2019, 42, 2577–2599. [Google Scholar] [CrossRef]
- Sher, A.; Sarwar, T.; Nawaz, A.; Ijaz, M.; Sattar, A.; Ahmad, S. Methods of Seed Priming. In Priming and Pretreatment of Seeds and Seedlings: Implication in Plant Stress Tolerance and Enhancing Productivity in Crop Plants; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Singapore, 2019; pp. 1–10. [Google Scholar] [CrossRef]
- Farahani, H.A.; Maroufi, K.; Sani, B. Seedling Production in Nigella (Nigella sativa L.) Under Nanopriming Method. In Proceedings of the ICABBBE 2012: International Conference on Agricultural, Biotechnology, Biological and Biosystems Engineering, Amsterdam, The Netherlands, 29 January 2012. [Google Scholar]
- Guha, T.; Ravikumar, K.V.G.; Mukherjee, A.; Mukherjee, A.; Kundu, R. Nanopriming with zero valent iron (nZVI) enhances germination and growth in aromatic rice cultivar (Oryza sativa cv. Gobindabhog L.). Plant Physiol. Biochem. 2018, 127, 403–413. [Google Scholar] [CrossRef]
- Li, R.; He, J.; Xie, H.; Wang, W.; Bose, S.K.; Sun, Y.; Hu, J.; Yin, H. Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int. J. Biol. Macromol. 2019, 126, 91–100. [Google Scholar] [CrossRef]
- Mittal, J.; Osheen, S.; Gupta, A.; Kumar, R. Carbon Nanomaterials in Agriculture. In Nanoscience for Sustainable Agriculture; Pudake, R.N., Chauhan, N., Kole, C., Eds.; Springer International Publishing: Cham, Sweitzerland, 2019; pp. 153–170. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Anand, A.; Nagarajan, S. Biochemical and biophysical changes associated with magnetopriming in germinating cucumber seeds. Plant Physiol. Biochem. 2012, 57, 67–73. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef]
- Chang, X.; Song, Z.; Xu, Y.; Gao, M. Effects of carbon nanotubes on growth of wheat seedlings and Cd uptake. Chemosphere 2020, 240, 124931. [Google Scholar] [CrossRef]
- Raja, K.; Sowmya, R.; Sudhagar, R.; Moorthy, P.S.; Govindaraju, K.; Subramanian, K.S. Biogenic ZnO and Cu nanoparticles to improve seed germination quality in blackgram (Vigna mungo). Mater. Lett. 2019, 235, 164–167. [Google Scholar] [CrossRef]
- Rai, P.K.; Kumar, V.; Lee, S.; Raza, N.; Kim, K.-H.; Ok, Y.S.; Tsang, D.C.W. Nanoparticle-plant interaction: Implications in energy, environment, and agriculture. Environ. Int. 2018, 119, 1–19. [Google Scholar] [CrossRef]
- Kottegoda, N.; Munaweera, I.; Madusanka, N.; Karunaratne, V. A green slow-release fertilizer composition based on urea-modified hydroxyapatite nanoparticles encapsulated wood. Curr. Sci. 2011, 101, 73–78. [Google Scholar]
- Zheng, L.; Hong, F.; Lu, S.; Liu, C. Effect of nano-TiO2 on strength of naturally aged seeds and growth of spinach. Biol. Trace Elem. Res. 2005, 104, 83–92. [Google Scholar] [CrossRef]
- Hao, Y.; Xu, B.; Ma, C.; Shang, J.; Gu, W.; Li, W.; Hou, T.; Xiang, Y.; Cao, W.; Xing, B.; et al. Synthesis of novel mesoporous carbon nanoparticles and their phytotoxicity to rice (Oryza sativa L.). J. Saudi Chem. Soc. 2019, 23, 75–82. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, Y.; Mao, J.; Chen, B. Effects of biochar nanoparticles on seed germination and seedling growth. Environ. Pollut. 2019, 256, 113409. [Google Scholar] [CrossRef]
- Canas, J.E.; Long, M.; Nations, S.; Vadan, R.; Dai, L.; Luo, M.; Ambikapathi, R.; Lee, E.H.; Olszyk, D. Effects of Functionalized and Nonfunctionalized Single-Walled Carbon Nanotubes on Root Elongation of Select Crop Species. Environ. Toxicol. Chem. 2008, 27, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Shojaei, T.R.; Salleh, M.A.M.; Tabatabaei, M.; Mobli, H.; Aghbashlo, M.; Rashid, S.A.; Tan, T. Chapter 11—Applications of Nanotechnology and Carbon Nanoparticles in Agriculture. In Synthesis, Technology and Applications of Carbon Nanomaterials; Rashid, S.A., Raja Othman, R.N.I., Hussein, M.Z., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–277. [Google Scholar] [CrossRef]
- Haghighi, M.; Silva, J.A.T. The effect of carbon nanotubes on the seed germination and seedling growth of four vegetable species. J. Crop Sci. Biotechnol. 2014, 17, 201–208. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, B.; Wang, Q.; Shi, X.; Xiao, Z.; Lin, J.; Fang, X. Carbon nanotubes as molecular transporters for walled plant cells. Nano Lett. 2009, 9, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, D.K.; Dasgupta-Schubert, N.; Cendejas, L.M.V.; Villegas, J.; Montoya, L.C.; Garcia, S.E.B. Interfacing carbon nanotubes (CNT) with plants: Enhancement of growth, water and ionic nutrient uptake in maize (Zea mays) and implications for nanoagriculture. Appl. Nanosci. 2014, 4, 577–591. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, S.; Kartoolinejad, D.; Naghdi, R. Effects of priming with multi-walled carbon nanotubes on seed physiological characteristics of Hopbush (Dodonaeaviscosa L.) under drought stress. Int. J. Environ. Stud. 2017, 74, 528–539. [Google Scholar] [CrossRef]
- Park, S.; Ahn, Y.-J. Multi-walled carbon nanotubes and silver nanoparticles differentially affect seed germination, chlorophyll content, and hydrogen peroxide accumulation in carrot (Daucus carota L.). Biocatal. Agric. Biotechnol. 2016, 8, 257–262. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Zia ur Rehman, M.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Sierra-Almeida, A. Assessing the importance of cold-stratification for seed germination in alpine plant species of the High-Andes of central Chile. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 125–131. [Google Scholar] [CrossRef]
- Kim, S.Y.; Warpeha, K.M.; Huber, S.C. The brassinosteroid receptor kinase, BRI1, plays a role in seed germination and the release of dormancy by cold stratification. J. Plant Physiol. 2019, 241, 153031. [Google Scholar] [CrossRef] [PubMed]
- Rosbakh, S.; Hülsmann, L.; Weinberger, I.; Bleicher, M.; Poschlod, P. Bleaching and cold stratification can break dormancy and improve seed germination in Cyperaceae. Aquat. Bot. 2019, 158, 103128. [Google Scholar] [CrossRef]
- Su, L.; Lan, Q.; Pritchard, H.W.; Xue, H.; Wang, X. Reactive oxygen species induced by cold stratification promote germination of Hedysarum scoparium seeds. Plant Physiol. Biochem. 2016, 109, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Samples, V. Accumulation Parameters and Seasonal Trends for PCBs in Temperate and Boreal Forest Plant Species. Environ. Sci. Technol. 2008, 42, 5911–5916. [Google Scholar]
- Savithramma, N.; Ankanna, S.; Bhumi, G. Effect of Nanoparticles on Seed Germination and Seedling Growth of Boswellia Ovalifoliolata—An Endemic and Endangered Medicinal Tree Taxon. Nano Vis. 2012, 2, 61–68. [Google Scholar] [CrossRef]
- Mahakham, W.; Theerakulpisut, P.; Maensiri, S.; Phumying, S.; Sarmah, A.K. Environmentally benign synthesis of phytochemicals-capped gold nanoparticles as nanopriming agent for promoting maize seed germination. Sci. Total Environ. 2016, 573, 1089–1102. [Google Scholar] [CrossRef]
- Naguib, D.M. Metabolic profiling during germination of hydro primed cotton seeds. Biocatal. Agric. Biotechnol. 2019, 17, 422–426. [Google Scholar] [CrossRef]
- Pachari Vera, E.; Alca, J.J.; Rondón Saravia, G.; Callejas Campioni, N.; Jachmanián Alpuy, I. Comparison of the lipid profile and tocopherol content of four Peruvian quinoa (Chenopodium quinoa Willd.) cultivars (‘Amarilla de Maranganí’, ‘Blanca de Juli’, INIA 415 ‘Roja Pasankalla’, INIA 420 ‘Negra Collana’) during germination. J. Cereal Sci. 2019, 88, 132–137. [Google Scholar] [CrossRef]
- Sun, J.; Jia, H.; Wang, P.; Zhou, T.; Wu, Y.; Liu, Z. Exogenous gibberellin weakens lipid breakdown by increasing soluble sugars levels in early germination of zanthoxylum seeds. Plant Sci. 2019, 280, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, J.; Su, L.; Sun, K.; Li, D.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. Metabolic profile analysis and identification of key metabolites during rice seed germination under low-temperature stress. Plant Sci. 2019, 289, 110282. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.H.; Bykova, N.V.; Igamberdiev, A.U. Cell signaling mechanisms and metabolic regulation of germination and dormancy in barley seeds. Crop J. 2017, 5, 459–477. [Google Scholar] [CrossRef]
- Ali, A.S.; Elozeiri, A.A. Metabolic Processes During Seed Germination. In Advances in Seed Biology; Jimenez-Lopez, J.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- Li-Shar, H.; Claus, G. Lipid and Fatty Acid Changes During Germination of Alfalfa Seeds. Phytochemistry 1990, 29, 1441–1445. [Google Scholar]
- Chadoeufhannel, R.; Taylorson, R.B. Changes in Lipid Fatty-Acids Associated with Dormancy Breaking in Amaranthus-Albus Seeds. J. Seed Technol. 1987, 11, 15–22. [Google Scholar]
- Smith, R.L. Seed Dormancy in Panicum-Maximum Jacq. Trop. Agric. 1979, 56, 233–239. [Google Scholar]
- ISTA. International Rules for Seed Testing, 2006 ed.; International Seed Testing Association: Bassersdorf, Switzerland, 2006. [Google Scholar]
- Paiva, E.P.D.; Torres, S.B.; Almeida, J.P.N.D.; Sá, F.V.D.S.; Oliveira, R.R.T. Tetrazolium test for the viability of gherkin seeds. Rev. Ciênc. Agron. 2017, 48, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.M.C.; Torres, S.B.; Sousa, E.C.; Sousa, D.M.M.; Pereira, K.T.O.; Paiva, E.P.D.; Matias, J.R.; Santos, B.R.V.D. Viability of Carica papaya L. Seeds by the Tetrazolium Test. J. Agric. Sci. 2018, 10, 335. [Google Scholar] [CrossRef] [Green Version]
- Vieira, R.D.; Neto, A.S.; de Bittencourt, S.R.M.; Panobianco, M. Electrical conductivity of the seed soaking solution and soybean seedling emergence. Sci. Agric. 2004, 61, 164–168. [Google Scholar] [CrossRef]
- Bligh, E.; Dyer, W. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Nadeem, M.; Pham, T.H.; Thomas, R.; Galagedara, L.; Kavanagh, V.; Zhu, X.; Ali, W.; Cheema, M. Potential role of root membrane phosphatidic acid in superior agronomic performance of silage-corn cultivated in cool climate cropping systems. Physiol. Plant. 2019. [Google Scholar] [CrossRef]
- Pham, T.H.; Zaeem, M.; Fillier, T.A.; Nadeem, M.; Vidal, N.P.; Manful, C.; Cheema, S.; Cheema, M.; Thomas, R.H. Targeting Modified Lipids during Routine Lipidomics Analysis using HILIC and C30 Reverse Phase Liquid Chromatography coupled to Mass Spectrometry. Sci. Rep. 2019, 9, 5048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, N.P.; Pham, H.T.; Manful, C.; Pumphrey, R.; Nadeem, M.; Cheema, M.; Galagedara, L.; Leke-Aladekoba, A.; Abbey, L.; Thomas, R. The use of natural media amendments to produce kale enhanced with functional lipids in controlled environment production system. Sci. Rep. 2018, 8, 14771. [Google Scholar] [CrossRef] [PubMed]
- Sawma, J.T.; Mohler, C.L. Evaluating Seed Viability by an Unimbibed Seed Crush Test in Comparison with the Tetrazolium Test. Weed Technol. 2017, 16, 781–786. [Google Scholar] [CrossRef]
- Anghinoni, F.B.; Braccini, A.L.; Anghinoni, G.; Jourdan, P.; Braz, G.B.; Albrecht, A.J.; Rubem, S.D.O. Improving small weed Seeds viability assessment using tetrazolium test. J. Agric. Sci. 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Fogaça, C.A.; Krohn, N.G.; de Aquino Souza, M.; de Paula, R.C. Teste de tetrazólio em sementes de Copaifera langsdorffii E schizolobium parahyba. Floresta 2011, 41, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Mishra, M.; Singh, N.I.; Chauhan, J.S. Determination of Integrity of Seed Membrane Systems and Mineral Ions Leakage in Horse Gram. World J. Agric. Sci. 2011, 7, 476–479. [Google Scholar]
- Piwowarczyk, B.; Tokarz, K.; Kamińska, I. Responses of grass pea seedlings to salinity stress in In Vitro culture conditions. Plant Cell Tissue Organ Cult. 2016, 124, 227–240. [Google Scholar] [CrossRef]
- Matthews, S.; Wagner, M.-H.; Kerr, L.; Powell, A.A. Potential for early counts of radicle emergence and leakage of electrolytes as quick tests to predict the percentage of normal seedlings. Seed Sci. Technol. 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Benamar, A.; Tallon, C.; Macherel, D. Membrane integrity and oxidative properties of mitochondria isolated from imbibing pea seeds after priming or accelerated ageing. Seed Sci. Res. 2003, 13, 35–45. [Google Scholar] [CrossRef]
- Kim, D.H. Practical methods for rapid seed germination from seed coat-imposed dormancy of Prunus yedoensis. Sci. Hortic. 2019, 243, 451–456. [Google Scholar] [CrossRef]
- Venier, P.; Carrizo García, C.; Cabido, M.; Funes, G. Survival and germination of three hard-seeded Acacia species after simulated cattle ingestion: The importance of the seed coat structure. S. Afr. J. Bot. 2012, 79, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Smreciu, A.; Gould, K. Field emergence of native boreal forest species on reclaimed sites in northeastern Alberta. Nativ. Plants J. 2015, 16, 204–226. [Google Scholar] [CrossRef]
- Poff, K.E.; Sharma, J.; Richards, M. Cold-Moist Stratification Improves Germination in a Temperate Terrestrial Orchid. Castanea 2016, 81, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.; Basu, R.; Das, S.; Nandy, P. Beneficial role of carbon nanotubes on mustard plant growth: An agricultural prospect. J. Nanopart. Res. 2011, 13, 4519–4528. [Google Scholar] [CrossRef]
- Zhao, X.; Joo, J.C.; Kim, D.; Lee, J.; Kim, J.Y. Estimation of the Seedling Vigor Index of Sunflowers Treated with Various Heavy Metals. J. Bioremed. Biodegrad. 2016, 7, 353. [Google Scholar] [CrossRef]
- Beerelli, K.; Balakrishnan, D.; Surapaneni, M.; Addanki, K.; Rao, Y.V.; Neelamraju, S. Evaluation of yield and seedling vigour related traits of Swarna/Oryza nivara backcross introgression lines under three environment conditions. Ecol. Genet. Genom. 2019, 11, 100036. [Google Scholar] [CrossRef]
- Rehman, H.U.; Iqbal, H.; Basra, S.M.A.; Afzal, I.; Farooq, M.; Wakeel, A.; Wang, N. Seed priming improves early seedling vigor, growth and productivity of spring maize. J. Integr. Agric. 2015, 14, 1745–1754. [Google Scholar] [CrossRef]
- Ménard, L.-P.; Ruel, J.-C.; Thiffault, N. Abundance and impacts of competing species on conifer regeneration following careful logging in the Eastern Canadian Boreal forest. Forests 2019, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef]
- Srinivasan, C.; Saraswathi, R. Nano-agriculture–carbon nanotubes enhance tomato seed germination and plant growth. Curr. Sci. 2010, 99, 274–275. [Google Scholar]
- Yu, X.; Li, A.; Li, W. How membranes organize during seed germination: Three patterns of dynamic lipid remodelling define chilling resistance and affect plastid biogenesis. Plant Cell Environ. 2015, 38, 1391–1403. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-X.; Xin, X.; Yin, G.-K.; He, J.-J.; Zhou, Y.-C.; Chen, J.-Y.; Lu, X.-X. Membrane phospholipids remodeling upon imbibition in Brassica napus L. seeds. Biochem. Biophys. Res. Commun. 2019, 515, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Sicilia, C.; Silvestre, S.; Haslam, R.P.; Michaelson, L.V. Lipid remodelling: Unravelling the response to cold stress in Arabidopsis and its extremophile relative Eutrema salsugineum. Plant Sci. 2017, 263, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Mamode Cassim, A.; Gouguet, P.; Gronnier, J.; Laurent, N.; Germain, V.; Grison, M.; Boutté, Y.; Gerbeau-Pissot, P.; Simon-Plas, F.; Mongrand, S. Plant lipids: Key players of plasma membrane organization and function. Progress Lipid Res. 2019, 73, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Galazzi, R.M.; Lopes Júnior, C.A.; de Lima, T.B.; Gozzo, F.C.; Arruda, M.A.Z. Evaluation of some effects on plant metabolism through proteins and enzymes in transgenic and non-transgenic soybeans after cultivation with silver nanoparticles. J. Proteom. 2019, 191, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Kaliamurthi, S.; Selvaraj, G.; Cakmak, Z.E.; Korkmaz, A.D.; Cakmak, T. The relationship between Chlorella sp. and zinc oxide nanoparticles: Changes in biochemical, oxygen evolution, and lipid production ability. Process Biochem. 2019, 85, 43–50. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Zapata, L.; Chalbi, N.; Carvajal, M. Multiwalled carbon nanotubes enter broccoli cells enhancing growth and water uptake of plants exposed to salinity. J. Nanobiotechnol. 2016, 14, 42. [Google Scholar] [CrossRef] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.H.; Sobze, J.-M.; Pham, T.H.; Nadeem, M.; Liu, C.; Galagedara, L.; Cheema, M.; Thomas, R. Carbon Nanoparticles Functionalized with Carboxylic Acid Improved the Germination and Seedling Vigor in Upland Boreal Forest Species. Nanomaterials 2020, 10, 176. https://doi.org/10.3390/nano10010176
Ali MH, Sobze J-M, Pham TH, Nadeem M, Liu C, Galagedara L, Cheema M, Thomas R. Carbon Nanoparticles Functionalized with Carboxylic Acid Improved the Germination and Seedling Vigor in Upland Boreal Forest Species. Nanomaterials. 2020; 10(1):176. https://doi.org/10.3390/nano10010176
Chicago/Turabian StyleAli, Md. Hossen, Jean-Marie Sobze, Thu Huong Pham, Muhammad Nadeem, Chen Liu, Lakshman Galagedara, Mumtaz Cheema, and Raymond Thomas. 2020. "Carbon Nanoparticles Functionalized with Carboxylic Acid Improved the Germination and Seedling Vigor in Upland Boreal Forest Species" Nanomaterials 10, no. 1: 176. https://doi.org/10.3390/nano10010176
APA StyleAli, M. H., Sobze, J. -M., Pham, T. H., Nadeem, M., Liu, C., Galagedara, L., Cheema, M., & Thomas, R. (2020). Carbon Nanoparticles Functionalized with Carboxylic Acid Improved the Germination and Seedling Vigor in Upland Boreal Forest Species. Nanomaterials, 10(1), 176. https://doi.org/10.3390/nano10010176