Investigation of the Electrical Properties of Microtubule Ensembles under Cell-Like Conditions

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tubulin Reconstitution
2.2. MT Polymerisation and Stabilisation
2.3. Fluorescence Imaging of MTs
2.4. Electrode Design and Device Construction
2.5. Impedance Measurements
2.6. Data Analysis
3. Results
3.1. Validation of Parallel-Plate Contact Device to Measure Dielectric Properties of Physiologically Relevant Ionic Solutions
3.2. The Effect of Microtubule Networks on Solution Capacitance at Physiologically Relevant Conditions
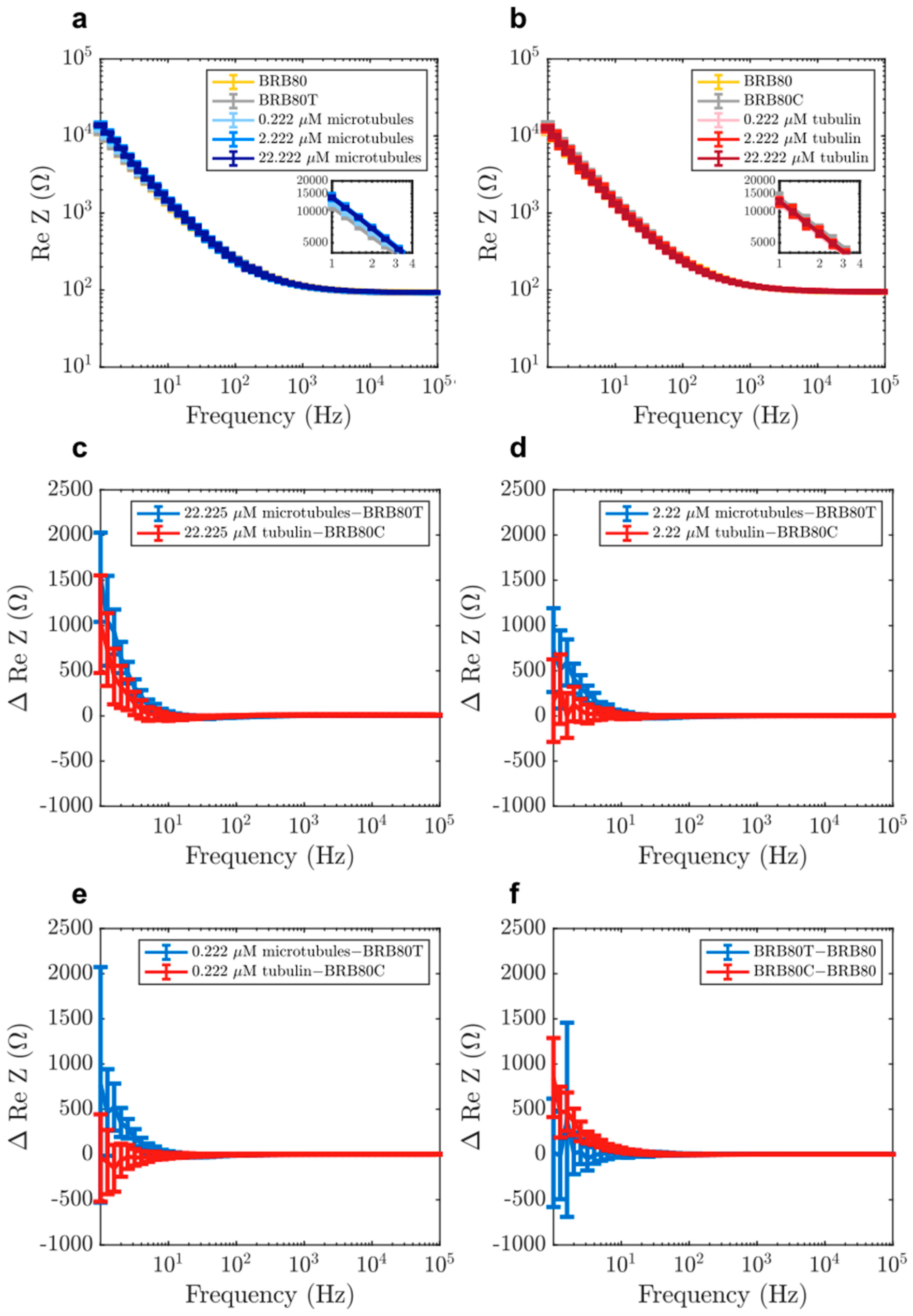
3.3. The Effect of Microtubule Networks on Solution Resistance at Physiologically Relevant Conditions
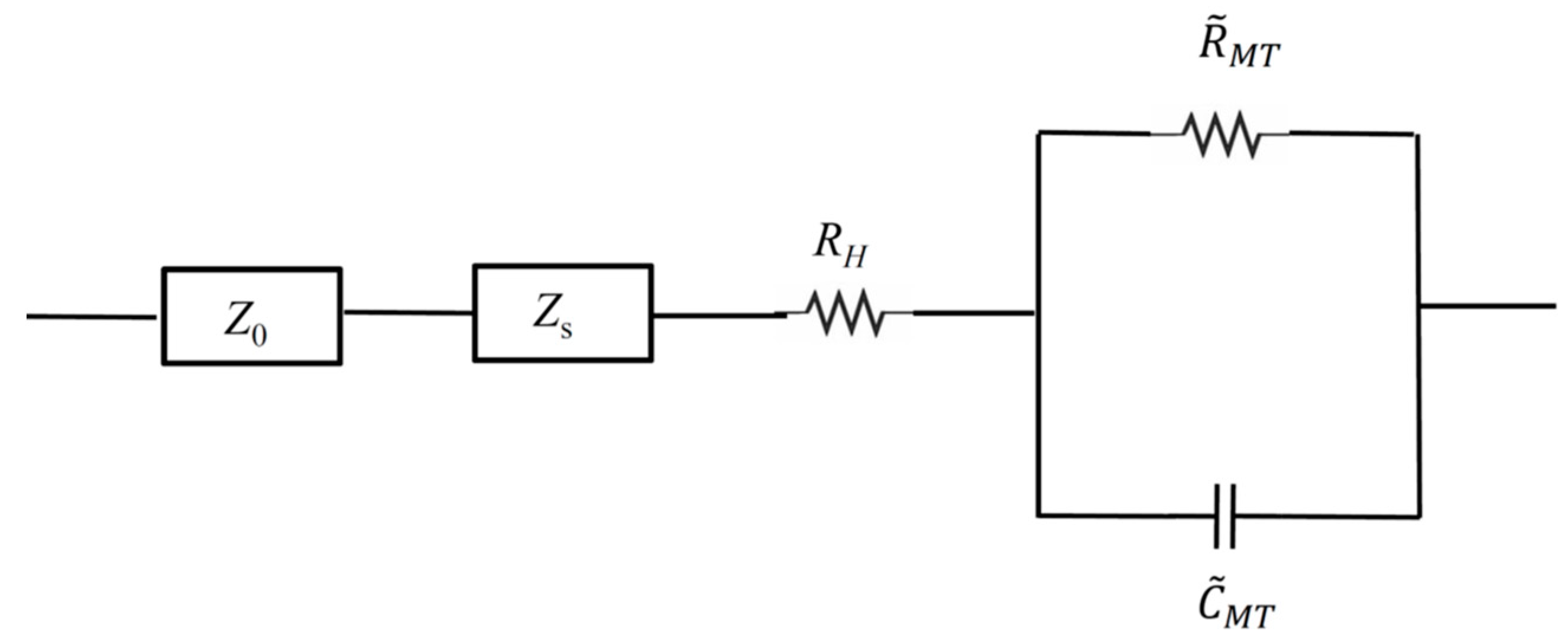
3.4. The Microtubule Network as an RC Circuit in Parallel
4. Discussion
4.1. The Physical Underpinnings of An Increased Capacitance
4.2. Implications for the Cell
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mershin, A.; Kolomenski, A.A.; Schuessler, H.A.; Nanopoulos, D.V. Tubulin dipole moment, dielectric constant and quantum behavior: Computer simulations, experimental results and suggestions. Biosystems 2004, 77, 73–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Heuvel, M.; De Graaff, M.; Lemay, S.; Dekker, C. Electrophoresis of individual microtubules in microchannels. Proc. Natl. Acad. Sci. USA 2007, 104, 7770–7775. [Google Scholar] [CrossRef] [Green Version]
- Timmons, J.J.; Preto, J.; Tuszynski, J.A.; Wong, E.T. Tubulin’s response to external electric fields by molecular dynamics simulations. PLoS ONE 2018, 13, e0202141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preto, J.; Pettini, M.; Tuszynski, J.A. Possible role of electrodynamic interactions in long-distance biomolecular recognition. Phys. Rev. E 2015, 91, 052710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barvitenko, N.; Lawen, A.; Aslam, M.; Pantaleo, A.; Saldanha, C.; Skverchinskaya, E.; Regolini, M.; Tuszynski, J.A. Integration of intracellular signaling: Biological analogues of wires, processors and memories organized by a centrosome 3D reference system. Biosystems 2018, 173, 191–206. [Google Scholar] [CrossRef]
- Friesen, D.E.; Craddock, T.J.; Kalra, A.P.; Tuszynski, J.A. Biological wires, communication systems, and implications for disease. Biosystems 2015, 127, 14–27. [Google Scholar] [CrossRef]
- Sekulić, D.L.; Satarić, B.M.; Tuszynski, J.A.; Satarić, M.V. Nonlinear ionic pulses along microtubules. Eur. Phys. J. E 2011, 34, 49. [Google Scholar] [CrossRef]
- Satarić, M.; Ilić, D.; Ralević, N.; Tuszynski, J.A. A nonlinear model of ionic wave propagation along microtubules. Eur. Biophys. J. 2009, 38, 637–647. [Google Scholar] [CrossRef]
- del Rocío Cantero, M.; Etchegoyen, C.V.; Perez, P.L.; Scarinci, N.; Cantiello, H.F. Bundles of Brain Microtubules Generate Electrical Oscillations. Sci. Rep. 2018, 8, 11899. [Google Scholar] [CrossRef]
- del Rocío Cantero, M.; Perez, P.L.; Smoler, M.; Etchegoyen, C.V.; Cantiello, H.F. Electrical oscillations in two-dimensional microtubular structures. Sci. Rep. 2016, 6, 27143. [Google Scholar] [CrossRef] [PubMed]
- Cifra, M.; Pokorný, J.; Havelka, D.; Kučera, O. Electric field generated by axial longitudinal vibration modes of microtubule. BioSystems 2010, 100, 122–131. [Google Scholar] [CrossRef]
- Uppalapati, M.; Huang, Y.M.; Jackson, T.N.; Hancock, W.O. Microtubule alignment and manipulation using AC electrokinetics. Small 2008, 4, 1371–1381. [Google Scholar] [CrossRef]
- Minoura, I.; Muto, E. Dielectric measurement of individual microtubules using the electroorientation method. Biophys. J. 2006, 90, 3739–3748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, R.; Böhm, K.; Wollweber, L.; Tuszynski, J.; Unger, E. Analysis of the migration behaviour of single microtubules in electric fields. Biochem. Biophys. Res. Commun. 2002, 293, 602–609. [Google Scholar] [CrossRef]
- Isozaki, N.; Ando, S.; Nakahara, T.; Shintaku, H.; Kotera, H.; Meyhöfer, E.; Yokokawa, R. Control of microtubule trajectory within an electric field by altering surface charge density. Sci. Rep. 2015, 5, 7669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujovne, I.; van den Heuvel, M.; Shen, Y.; de Graaff, M.; Dekker, C. Velocity modulation of microtubules in electric fields. Nano Lett. 2008, 8, 4217–4220. [Google Scholar] [CrossRef] [PubMed]
- Kirson, E.D.; Gurvich, Z.; Schneiderman, R.; Dekel, E.; Itzhaki, A.; Wasserman, Y.; Schatzberger, R.; Palti, Y. Disruption of cancer cell replication by alternating electric fields. Cancer Res. 2004, 64, 3288–3295. [Google Scholar] [CrossRef] [Green Version]
- Giladi, M.; Schneiderman, R.S.; Voloshin, T.; Porat, Y.; Munster, M.; Blat, R.; Sherbo, S.; Bomzon, Z.; Urman, N.; Itzhaki, A. Mitotic spindle disruption by alternating electric fields leads to improper chromosome segregation and mitotic catastrophe in cancer cells. Sci. Rep. 2015, 5, 18046. [Google Scholar] [CrossRef]
- Tuszynski, J.; Wenger, C.; Friesen, D.; Preto, J. An overview of sub-cellular mechanisms involved in the action of TTFields. Int. J. Environ. Res. Public Health 2016, 13, 1128. [Google Scholar] [CrossRef] [Green Version]
- Santelices, I.B.; Friesen, D.E.; Bell, C.; Hough, C.M.; Xiao, J.; Kalra, A.; Kar, P.; Freedman, H.; Rezania, V.; Lewis, J.D. Response to Alternating Electric Fields of Tubulin Dimers and Microtubule Ensembles in Electrolytic Solutions. Sci. Rep. 2017, 7, 9594. [Google Scholar] [CrossRef] [Green Version]
- Shelden, E.; Wadsworth, P. Observation and quantification of individual microtubule behavior in vivo: Microtubule dynamics are cell-type specific. J. Cell Biol. 1993, 120, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Van de Water, L.; Olmsted, J. The quantitation of tubulin in neuroblastoma cells by radioimmunoassay. J. Biol. Chem. 1980, 255, 10744–10751. [Google Scholar] [PubMed]
- Olmsted, J.; Borisy, G. Ionic and nucleotide requirements for microtubule polymerization in vitro. Biochemistry 1975, 14, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Weisenberg, R.C. Microtubule formation in vitro in solutions containing low calcium concentrations. Science 1972, 177, 1104–1105. [Google Scholar] [CrossRef]
- Böhm, K.J.; Mavromatos, N.E.; Michette, A.; Stracke, R.; Unger, E. Movement and alignment of microtubules in electric fields and electric-dipole-moment estimates. Electromagn. Biol. Med. 2005, 24, 319–330. [Google Scholar] [CrossRef]
- Umnov, M.; Palusinski, O.; Deymier, P.; Guzman, R.; Hoying, J.; Barnaby, H.; Yang, Y.; Raghavan, S. Experimental evaluation of electrical conductivity of microtubules. J. Mater. Sci. 2007, 42, 373–378. [Google Scholar] [CrossRef]
- van Eunen, K.; Bouwman, J.; Daran-Lapujade, P.; Postmus, J.; Canelas, A.B.; Mensonides, F.I.; Orij, R.; Tuzun, I.; van den Brink, J.; Smits, G.J. Measuring enzyme activities under standardized in vivo-like conditions for systems biology. Febs J. 2010, 277, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Theillet, F.-X.; Binolfi, A.; Frembgen-Kesner, T.; Hingorani, K.; Sarkar, M.; Kyne, C.; Li, C.; Crowley, P.B.; Gierasch, L.; Pielak, G.J. Physicochemical properties of cells and their effects on intrinsically disordered proteins (IDPs). Chem. Rev. 2014, 114, 6661–6714. [Google Scholar] [CrossRef]
- Fritzsche, W.; Böhm, K.; Unger, E.; Köhler, J.M. Making electrical contact to single molecules. Nanotechnology 1998, 9, 177. [Google Scholar] [CrossRef]
- Fritzsche, W.; Köhler, J.M.; Böhm, K.J.; Unger, E.; Wagner, T.; Kirsch, R.; Mertig, M.; Pompe, W. Wiring of metallized microtubules by electron beam-induced structuring. Nanotechnology 1999, 10, 331. [Google Scholar] [CrossRef]
- Gell, C.; Bormuth, V.; Brouhard, G.J.; Cohen, D.N.; Diez, S.; Friel, C.T.; Helenius, J.; Nitzsche, B.; Petzold, H.; Ribbe, J. Microtubule dynamics reconstituted in vitro and imaged by single-molecule fluorescence microscopy. In Methods in Cell Biology; Elsevier: New York, NY, USA, 2010; Volume 95, pp. 221–245. [Google Scholar]
- Taylor, S.; Gileadi, E. Physical interpretation of the Warburg impedance. Corrosion 1995, 51, 664–671. [Google Scholar] [CrossRef]
- Ramos-Barrado, J.; Galan Montenegro, P.; Cambón, C.C. A generalized Warburg impedance for a nonvanishing relaxation process. J. Chem. Phys. 1996, 105, 2813–2815. [Google Scholar] [CrossRef] [Green Version]
- Sanabria, H.; Miller, J.H., Jr. Relaxation processes due to the electrode-electrolyte interface in ionic solutions. Phys. Rev. E 2006, 74, 051505. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.F.; Vieira, A.L.; Mukai, H.; Andrade, C.M.; Fernandes, P.R. Electric impedance of aqueous KCl and NaCl solutions: Salt concentration dependence on components of the equivalent electric circuit. J. Mol. Liq. 2017, 241, 530–539. [Google Scholar] [CrossRef]
- Lenzi, E.; Fernandes, P.; Petrucci, T.; Mukai, H.; Ribeiro, H.; Lenzi, M.; Gonçalves, G. Anomalous diffusion and electrical response of ionic solutions. Int. J. Electrochem. Sci 2013, 8, 2849–2862. [Google Scholar]
- Wilson, L.; Miller, H.P.; Farrell, K.W.; Snyder, K.B.; Thompson, W.C.; Purich, D.L. Taxol stabilization of microtubules in vitro: Dynamics of tubulin addition and loss at opposite microtubule ends. Biochemistry 1985, 24, 5254–5262. [Google Scholar] [CrossRef]
- Arnal, I.; Wade, R.H. How does taxol stabilize microtubules? Curr. Biol. 1995, 5, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Mikhaylova, M.; Cloin, B.M.; Finan, K.; Van Den Berg, R.; Teeuw, J.; Kijanka, M.M.; Sokolowski, M.; Katrukha, E.A.; Maidorn, M.; Opazo, F. Resolving bundled microtubules using anti-tubulin nanobodies. Nat. Commun. 2015, 6, 7933. [Google Scholar] [CrossRef]
- Herold, C.; Leduc, C.; Stock, R.; Diez, S.; Schwille, P. Long-Range Transport of Giant Vesicles along Microtubule Networks. ChemPhysChem 2012, 13, 1001–1006. [Google Scholar] [CrossRef]
- Kulić, I.M.; Brown, A.E.; Kim, H.; Kural, C.; Blehm, B.; Selvin, P.R.; Nelson, P.C.; Gelfand, V.I. The role of microtubule movement in bidirectional organelle transport. Proc. Natl. Acad. Sci. USA 2008, 105, 10011–10016. [Google Scholar] [CrossRef] [Green Version]
- Margolis, R.L.; Wilson, L. Addition of colchicine-tubulin complex to microtubule ends: The mechanism of substoichiometric colchicine poisoning. Proc. Natl. Acad. Sci. USA 1977, 74, 3466–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmsted, J.; Borisy, G. Characterization of microtubule assembly in porcine brain extracts by viscometry. Biochemistry 1973, 12, 4282–4289. [Google Scholar] [CrossRef] [PubMed]
- Satarić, M.; Sekulić, D.; Živanov, M. Solitonic ionic currents along microtubules. J. Comput. Theor. Nanosci. 2010, 7, 2281–2290. [Google Scholar] [CrossRef]
- Freedman, H.; Rezania, V.; Priel, A.; Carpenter, E.; Noskov, S.Y.; Tuszynski, J.A. Model of ionic currents through microtubule nanopores and the lumen. Phys. Rev. E 2010, 81, 051912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanabria, H.; Miller, J.H., Jr.; Mershin, A.; Luduena, R.F.; Kolomenski, A.A.; Schuessler, H.A.; Nanopoulos, D.V. Impedance spectroscopy of α-β tubulin heterodimer suspensions. Biophys. J. 2006, 90, 4644–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Guo, W. Ion Permeability of a Microtubule in Neuron Environment. J. Phys. Chem. Lett. 2018, 9, 2009–2014. [Google Scholar] [CrossRef]
- Priel, A.; Tuszyński, J. A nonlinear cable-like model of amplified ionic wave propagation along microtubules. Epl (Europhys. Lett.) 2008, 83, 68004. [Google Scholar] [CrossRef]
- Tuszyński, J.; Brown, J.; Crawford, E.; Carpenter, E.; Nip, M.; Dixon, J.; Satarić, M. Molecular dynamics simulations of tubulin structure and calculations of electrostatic properties of microtubules. Math. Comput. Model. 2005, 41, 1055–1070. [Google Scholar] [CrossRef]
- Sekulić, D.L.; Satarić, M.V. Microtubule as nanobioelectronic nonlinear circuit. Serb. J. Electr. Eng. 2012, 9, 107–119. [Google Scholar] [CrossRef]
- Manning, G.S. Limiting laws and counterion condensation in polyelectrolyte solutions I. Colligative properties. J. Chem. Phys. 1969, 51, 924–933. [Google Scholar] [CrossRef]
- Tuszyński, J.; Portet, S.; Dixon, J.; Luxford, C.; Cantiello, H. Ionic wave propagation along actin filaments. Biophys. J. 2004, 86, 1890–1903. [Google Scholar] [CrossRef] [Green Version]
- Priel, A.; Ramos, A.J.; Tuszynski, J.A.; Cantiello, H.F. A biopolymer transistor: Electrical amplification by microtubules. Biophys. J. 2006, 90, 4639–4643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aouaichia, M.; McCullen, N.; Bowen, C.R.; Almond, D.P.; Budd, C.; Bouamrane, R. Understanding the anomalous frequency responses of composite materials using very large random resistor-capacitor networks. Eur. Phys. J. B 2017, 90, 39. [Google Scholar] [CrossRef] [Green Version]
- Panteny, S.; Stevens, R.; Bowen, C. The frequency dependent permittivity and AC conductivity of random electrical networks. Ferroelectrics 2005, 319, 199–208. [Google Scholar] [CrossRef]
- Parri, H.R.; Gould, T.M.; Crunelli, V. Spontaneous astrocytic Ca 2+ oscillations in situ drive NMDAR-mediated neuronal excitation. Nat. Neurosci. 2001, 4, 803. [Google Scholar] [CrossRef]
- Regula, C.; Berlin, R. Microtubule assembly and disassembly at alkaline pH. J. Cell Biol. 1981, 89, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Borisy, G.; Marcum, J.; Olmsted, J.; Murphy, D.; Johnson, K. Purification of tubulin and associated high molecular weight proteins from porcine brain and characterization of microtubule assembly in vitro. Ann. New York Acad. Sci. USA 1975, 253, 107–132. [Google Scholar] [CrossRef]
- Larsson, H.; Wallin, M.; Edström, A. Induction of a sheet polymer of tubulin by Zn2+. Exp. Cell Res. 1976, 100, 104–110. [Google Scholar] [CrossRef]
- Nicholson, W.V.; Lee, M.; Downing, K.H.; Nogales, E. Cryo-electron microscopy of GDP-tubulin rings. Cell Biochem. Biophys. 1999, 31, 175–183. [Google Scholar] [CrossRef]
- Zhai, Y.; Kronebusch, P.J.; Simon, P.M.; Borisy, G.G. Microtubule dynamics at the G2/M transition: Abrupt breakdown of cytoplasmic microtubules at nuclear envelope breakdown and implications for spindle morphogenesis. J. Cell Biol. 1996, 135, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Borisy, G.G. Quantitative determination of the proportion of microtubule polymer present during the mitosis-interphase transition. J. Cell Sci. 1994, 107, 881–890. [Google Scholar] [PubMed]
- Kline-Smith, S.L.; Walczak, C.E. Mitotic spindle assembly and chromosome segregation: Refocusing on microtubule dynamics. Mol. Cell 2004, 15, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Akhmanova, A.; Dogterom, M. Kinesins lead aging microtubules to catastrophe. Cell 2011, 147, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugiel, M.; Mitra, A.; Girardo, S.; Diez, S.; Schäffer, E. Measuring microtubule supertwist and defects by three-dimensional-force-clamp tracking of single kinesin-1 motors. Nano Lett. 2018, 18, 1290–1295. [Google Scholar] [CrossRef]
- Diez, S.; Reuther, C.; Dinu, C.; Seidel, R.; Mertig, M.; Pompe, W.; Howard, J. Stretching and transporting DNA molecules using motor proteins. Nano Lett. 2003, 3, 1251–1254. [Google Scholar] [CrossRef]
- Hess, H.; Saper, G. Engineering with biomolecular motors. Acc. Chem. Res. 2018, 51, 3015–3022. [Google Scholar] [CrossRef]
- Song, W.; Zhu, J.; Kong, W.; Möhwald, H.; Li, J. Different Microtubule Structures Assembled by Kinesin Motors. Langmuir 2018, 34, 9768–9773. [Google Scholar] [CrossRef]
- Tas, R.P.; Chen, C.-Y.; Katrukha, E.A.; Vleugel, M.; Kok, M.; Dogterom, M.; Akhmanova, A.; Kapitein, L.C. Guided by Light: Optical Control of Microtubule Gliding Assays. Nano Lett. 2018, 18, 7524–7528. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, K.; Kitamura, M.; Yokokawa, M.; Kanno, I.; Kotera, H.; Yokokawa, R. Colocalization of quantum dots by reactive molecules carried by motor proteins on polarized microtubule arrays. Acs Nano 2012, 7, 447–455. [Google Scholar] [CrossRef]
- Adrian, M.; Nijenhuis, W.; Hoogstraaten, R.I.; Willems, J.; Kapitein, L.C. A phytochrome-derived photoswitch for intracellular transport. Acs Synth. Biol. 2017, 6, 1248–1256. [Google Scholar] [CrossRef]
- Lam, A.T.-C.; Tsitkov, S.; Zhang, Y.; Hess, H. Reversibly bound kinesin-1 motor proteins propelling microtubules demonstrate dynamic recruitment of active building blocks. Nano Lett. 2018, 18, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Yokokawa, R.; Takeuchi, S.; Kon, T.; Nishiura, M.; Sutoh, K.; Fujita, H. Unidirectional transport of kinesin-coated beads on microtubules oriented in a microfluidic device. Nano Lett. 2004, 4, 2265–2270. [Google Scholar] [CrossRef]
- Fujimoto, K.; Morita, Y.; Iino, R.; Tomishige, M.; Shintaku, H.; Kotera, H.; Yokokawa, R. Simultaneous Observation of Kinesin-Driven Microtubule Motility and Binding of Adenosine Triphosphate Using Linear Zero-Mode Waveguides. Acs Nano 2018, 12, 11975–11985. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jia, Y.; Dong, W.; Feng, X.; Fei, J.; Li, J. Transporting a Tube in a Tube. Nano Lett. 2014, 14, 6160–6164. [Google Scholar] [CrossRef]
- Wieczorek, M.; Chaaban, S.; Brouhard, G.J. Macromolecular crowding pushes catalyzed microtubule growth to near the theoretical limit. Cell. Mol. Bioeng. 2013, 6, 383–392. [Google Scholar] [CrossRef]
- Sozański, K.; Ruhnow, F.; Wiśniewska, A.; Tabaka, M.; Diez, S.; Hołyst, R. Small crowders slow down kinesin-1 stepping by hindering motor domain diffusion. Phys. Rev. Lett. 2015, 115, 218102. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tubulin Concentration (μM) | Volume of BRB80T or BRB80C (μL) | Tubulin Volume (μL) |
|---|---|---|
| 0.222 | 99.5 | 0.5 |
| 2.222 | 95 | 5 |
| 22.225 | 5 | 5 |
| [Tub] (µM). | CMT (F). | δCMT (F) | RMT (Ω) | δRMT (Ω) | RH (Ω) | δRH (Ω) | γR (Ω/L) | γC (F/L) | δγR (Ω/L) | δγC (F/L) |
|---|---|---|---|---|---|---|---|---|---|---|
| 22.222 | 1.27 × 10−5 | 1.48 × 10−7 | 9.74 × 104 | 1.18 × 104 | 2.12 | 40.61 | 3.71 × 1010 | 7.65 | 4.49 × 109 | 0.056 |
| 2.222 | 1.25 × 10−5 | 1.67 × 10−7 | 1.00 × 105 | 1.40 × 104 | 0.61 | 34.79 | 3.81 × 1010 | 4.76 | 5.33 × 109 | 0.063 |
| 0.222 | 2.01 × 10−5 | 3.38 × 10−7 | 9.97 × 104 | 2.82 × 104 | 0.41 | 31.95 | 3.80 × 1010 | 4.83 | 1.07 × 1010 | 0.12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalra, A.P.; Patel, S.D.; Bhuiyan, A.F.; Preto, J.; Scheuer, K.G.; Mohammed, U.; Lewis, J.D.; Rezania, V.; Shankar, K.; Tuszynski, J.A. Investigation of the Electrical Properties of Microtubule Ensembles under Cell-Like Conditions. Nanomaterials 2020, 10, 265. https://doi.org/10.3390/nano10020265
Kalra AP, Patel SD, Bhuiyan AF, Preto J, Scheuer KG, Mohammed U, Lewis JD, Rezania V, Shankar K, Tuszynski JA. Investigation of the Electrical Properties of Microtubule Ensembles under Cell-Like Conditions. Nanomaterials. 2020; 10(2):265. https://doi.org/10.3390/nano10020265
Chicago/Turabian StyleKalra, Aarat P., Sahil D. Patel, Asadullah F. Bhuiyan, Jordane Preto, Kyle G. Scheuer, Usman Mohammed, John D. Lewis, Vahid Rezania, Karthik Shankar, and Jack A. Tuszynski. 2020. "Investigation of the Electrical Properties of Microtubule Ensembles under Cell-Like Conditions" Nanomaterials 10, no. 2: 265. https://doi.org/10.3390/nano10020265
APA StyleKalra, A. P., Patel, S. D., Bhuiyan, A. F., Preto, J., Scheuer, K. G., Mohammed, U., Lewis, J. D., Rezania, V., Shankar, K., & Tuszynski, J. A. (2020). Investigation of the Electrical Properties of Microtubule Ensembles under Cell-Like Conditions. Nanomaterials, 10(2), 265. https://doi.org/10.3390/nano10020265