In Vitro Interactions of TiO2 Nanoparticles with Earthworm Coelomocytes: Immunotoxicity Assessment

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Handling, Sample Collection, and Culture Medium Preparation
2.2. TiO2 NPs Characterization
2.3. Electron Microscopy Analyses
2.3.1. Cell Preparation
2.3.2. Scanning Electron Microscopy (SEM)
2.3.3. Transmission Electron Microscopy (TEM)
2.4. Flow Cytometry Assays
2.5. Malondialdehyde (MDA) Production and Alkaline Comet Assay
2.6. mRNA Levels Quantification
2.7. Statistical Analyses
3. Results
3.1. TiO2 NPs Characterization
3.2. Electron Microscopy
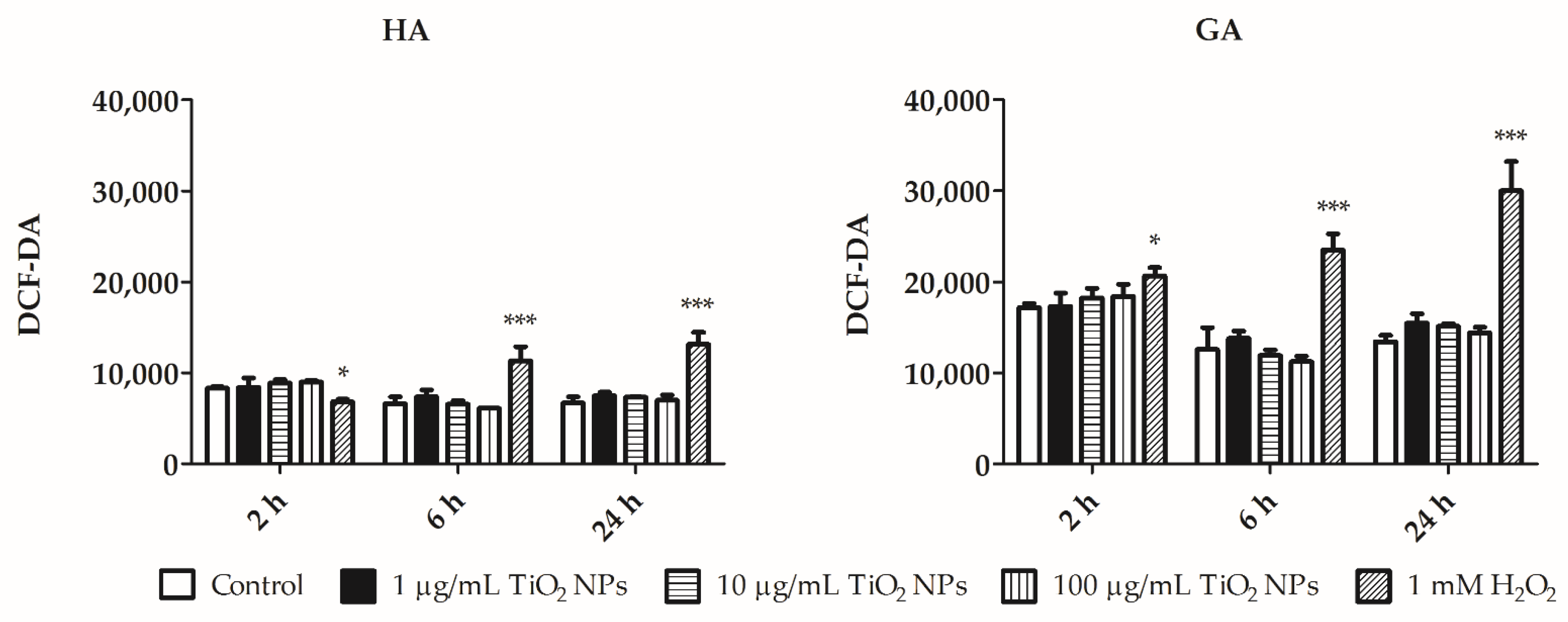
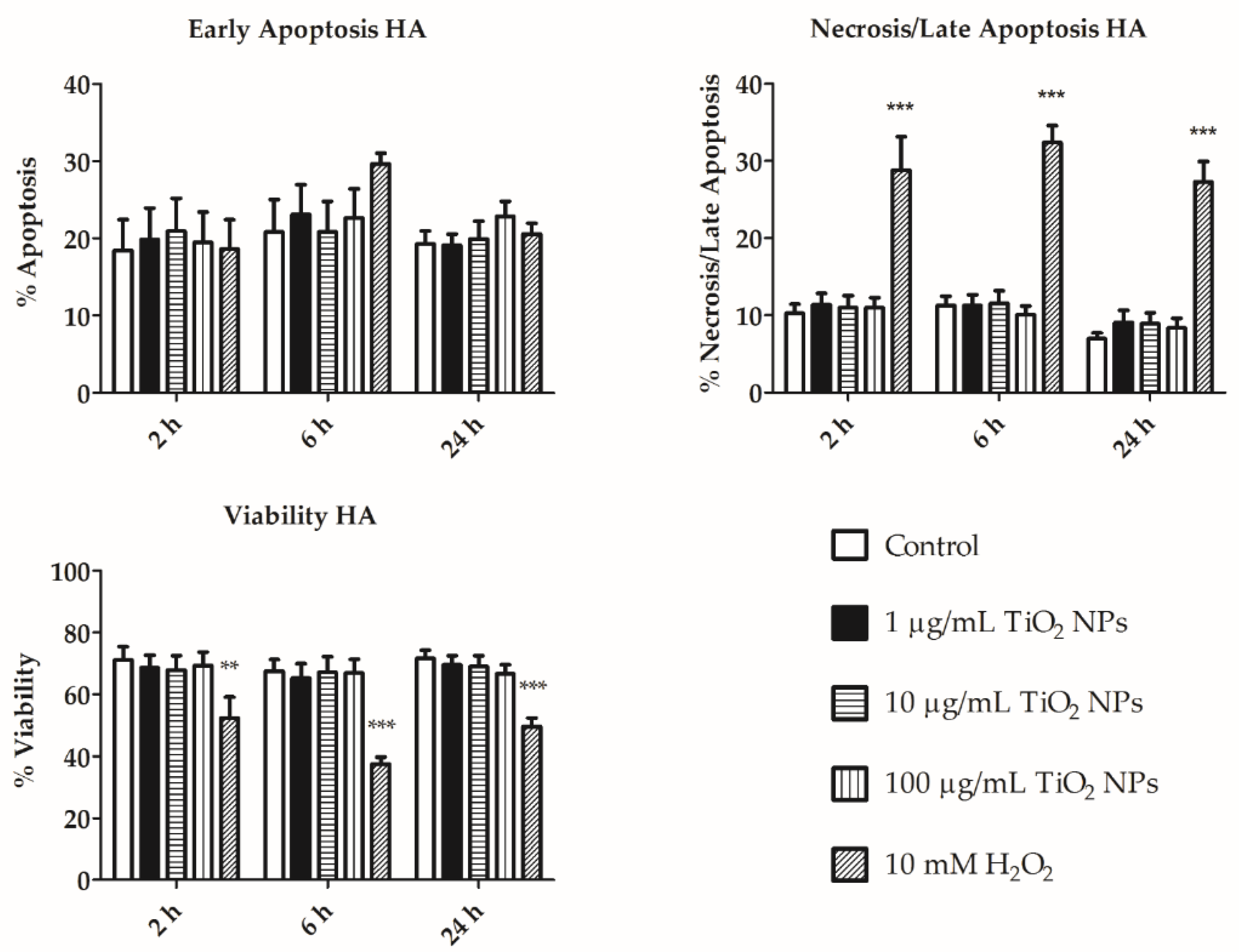
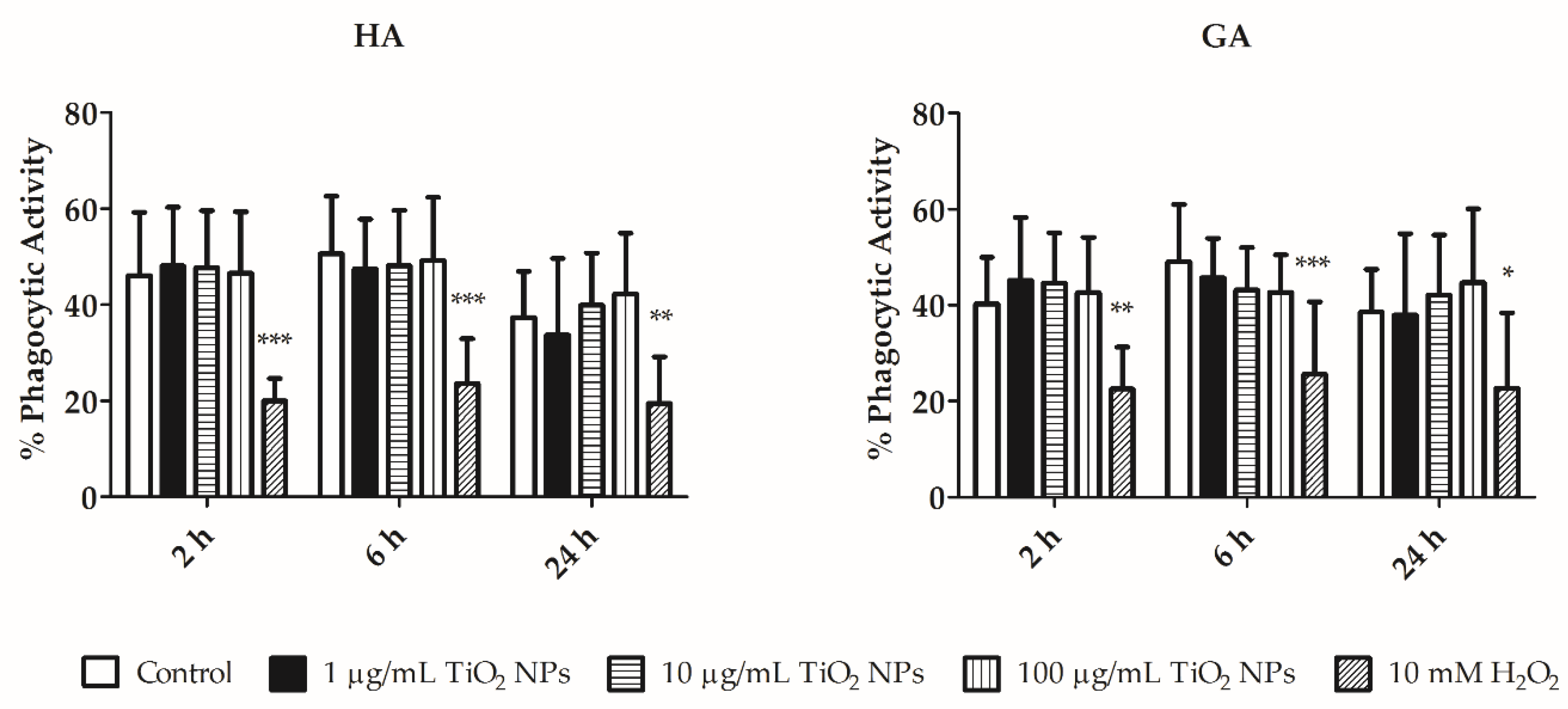
3.3. Flow Cytometry Assays
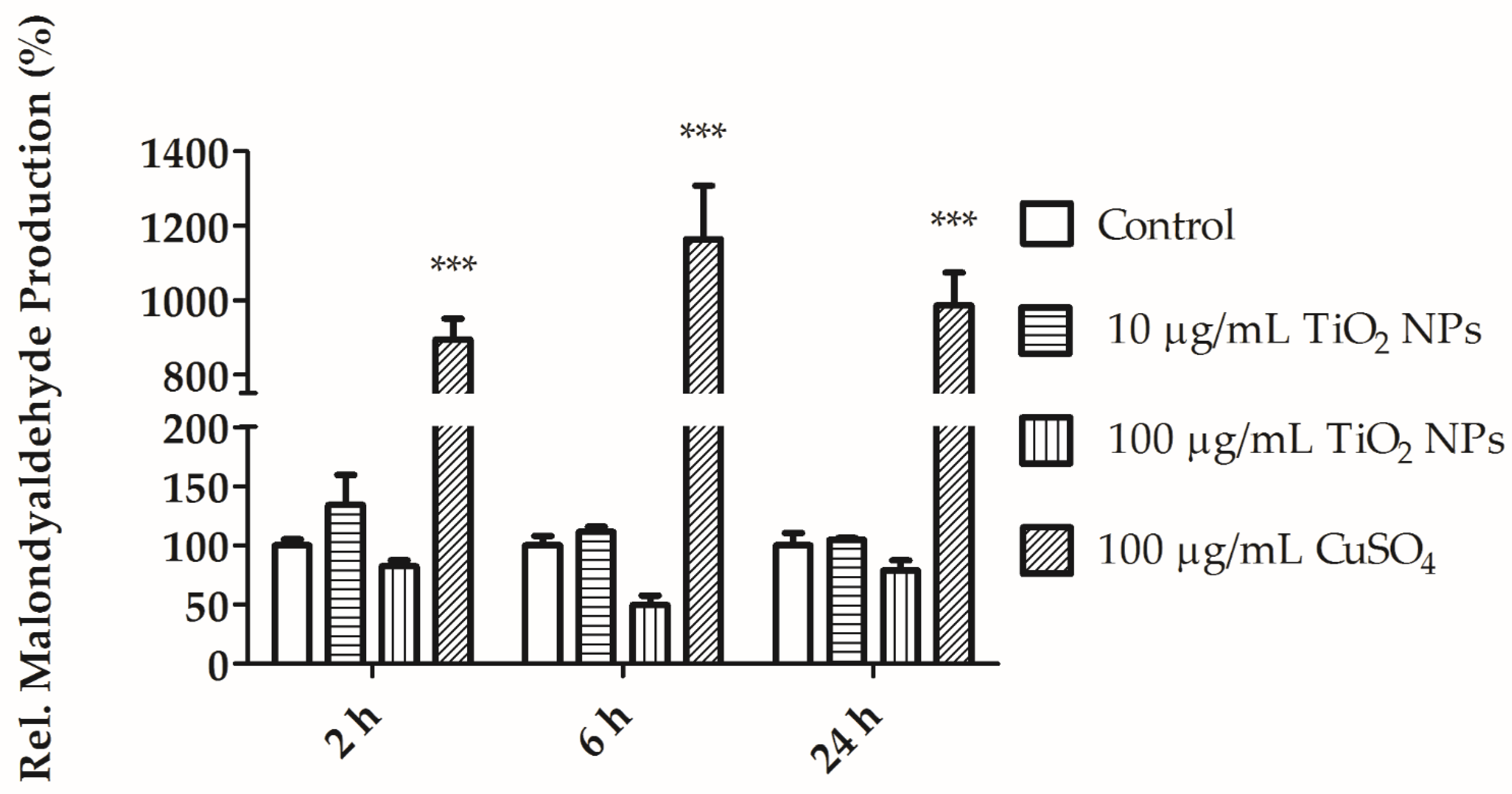
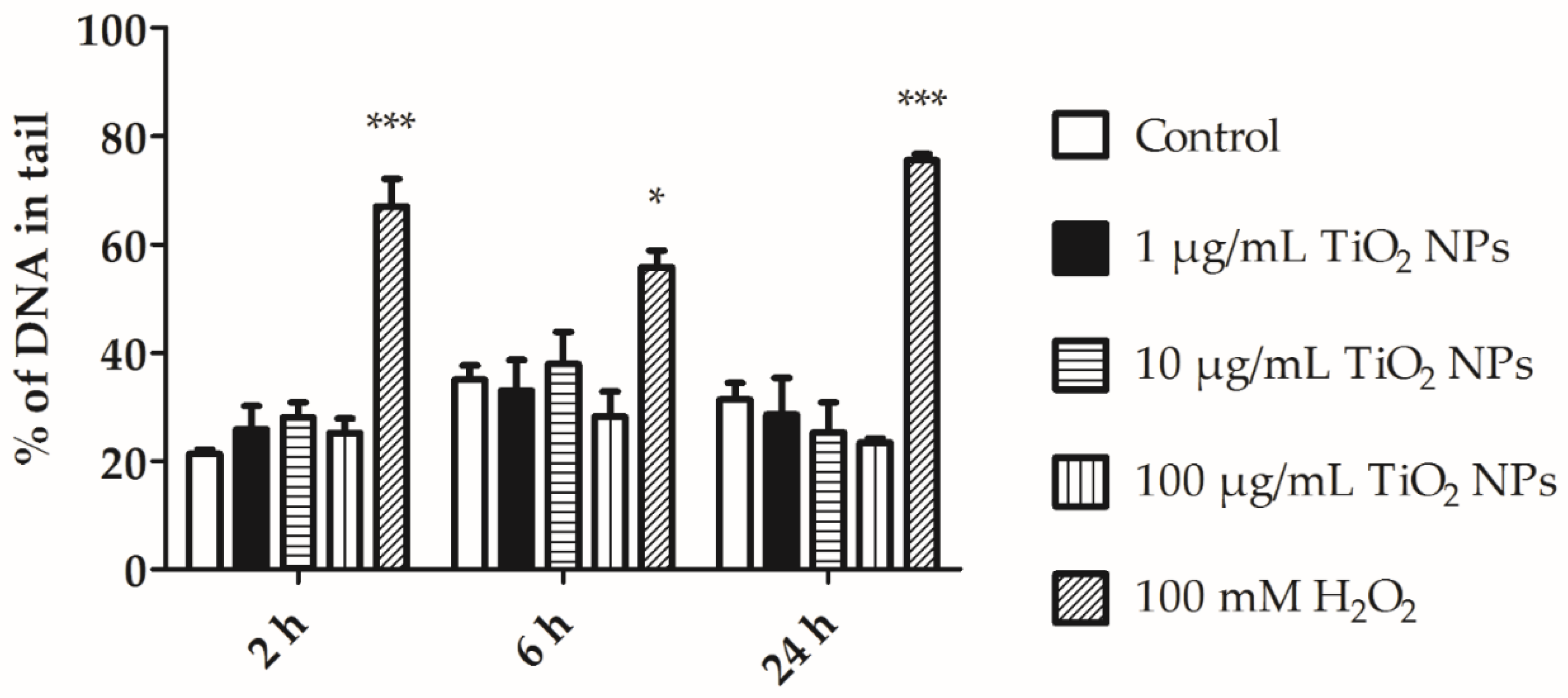
3.4. MDA and Alkaline Comet Assay
3.5. mRNA Levels of Detoxification, Immune, Antioxidant, and Signal Transduction Molecules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, S.M.; Tripathi, M. A review of TiO2 nanoparticles. Chin. Sci. Bull. 2011, 56, 1639. [Google Scholar] [CrossRef] [Green Version]
- Gautam, A.; Ray, A.; Mukherjee, S.; Das, S.; Pal, K.; Das, S.; Karmakar, P.; Ray, M.; Ray, S. Immunotoxicity of copper nanoparticle and copper sulfate in a common Indian earthworm. Ecotoxicol. Environ. Saf. 2018, 148, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.I.; An, Y.-J. Ecotoxicological Effects of Nanomaterials on Earthworms: A Review. Hum. Ecol. Risk Assess. 2015, 21, 1566–1575. [Google Scholar] [CrossRef]
- OECD. Guideline for the Testing of Chemicals. In No. 207, Earthworm, Acute Toxicity Tests; Organisation for Economic Cooperation and Development: Paris, France, 1984. [Google Scholar]
- OECD. Guideline for the Testing of Chemicals. In No. 222, Earthworm Reproduction Test (Eisenia Fetida/Eisenia Andrei); Organisation for Economic Cooperation and Development: Paris, France, 2004. [Google Scholar]
- OECD. Guidelines for the testing of chemicals. In No. 317, Bioaccumulation in Terrestrial Oligochaetes; Organisation for Economic Cooperation and Development: Paris, France, 2010. [Google Scholar]
- Valerio-Rodríguez, M.F.; Trejo-Téllez, L.I.; Aguilar-González, M.A.; Medina-Pérez, G.; Zúñiga-Enríquez, J.C.; Ortegón-Pérez, A.; Fernández-Luqueño, F. Effects of ZnO, TiO2 or Fe2O3 Nanoparticles on the Body Mass, Reproduction, and Survival of Eisenia fetida. Pol. J. Environ. Stud. 2020, 29, 2383–2394. [Google Scholar] [CrossRef]
- Šíma, P. Annelid coelomocytes and haemocytes: Roles in cellular immune reactions. In Immunology of Annelids; CRC Press: Boca Raton, FL, USA, 1994; pp. 115–165. [Google Scholar]
- Hayashi, Y.; Engelmann, P.; Foldbjerg, R.; Szabo, M.; Somogyi, I.; Pollak, E.; Molnar, L.; Autrup, H.; Sutherland, D.S.; Scott-Fordsmand, J.; et al. Earthworms and humans in vitro: Characterizing evolutionarily conserved stress and immune responses to silver nanoparticles. Environ. Sci. Technol. 2012, 46, 4166–4173. [Google Scholar] [CrossRef]
- Kwak, J.I.; Park, J.-W.; An, Y.-J. Effects of silver nanowire length and exposure route on cytotoxicity to earthworms. Environ. Sci. Pollut. Res. 2017, 24, 14516–14524. [Google Scholar] [CrossRef]
- Gupta, S.; Kushwah, T.; Yadav, S. Earthworm coelomocytes as nanoscavenger of ZnO NPs. Nanoscale Res. Lett. 2014, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wang, N.; Zhu, M.; Lu, J.; Zhong, H.; Xue, X.; Guo, S.; Li, M.; Wei, X.; Tao, Y.; et al. TiO2 nanoparticles cause mitochondrial dysfunction, activate inflammatory responses, and attenuate phagocytosis in macrophages: A proteomic and metabolomic insight. Redox Biol. 2018, 15, 266–276. [Google Scholar] [CrossRef]
- Alijagic, A.; Gaglio, D.; Napodano, E.; Russo, R.; Costa, C.; Benada, O.; Kofronova, O.; Pinsino, A. Titanium dioxide nanoparticles temporarily influence the sea urchin immunological state suppressing inflammatory-relate gene transcription and boosting antioxidant metabolic activity. J. Hazard. Mater. 2020, 384, 121389. [Google Scholar] [CrossRef]
- Bigorgne, E.; Foucaud, L.; Caillet, C.; Giamberini, L.; Nahmani, J.; Thomas, F.; Rodius, F. Cellular and molecular responses of E. fetida coelomocytes exposed to TiO2 nanoparticles. J. Nanopart. Res. 2012, 14. [Google Scholar] [CrossRef]
- Bigorgne, E.; Foucaud, L.; Lapied, E.; Labille, J.; Botta, C.; Sirguey, C.; Falla, J.; Rose, J.; Joner, E.J.; Rodius, F.; et al. Ecotoxicological assessment of TiO2 byproducts on the earthworm Eisenia fetida. Environ. Pollut. 2011, 159, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Lapied, E.; Nahmani, J.Y.; Moudilou, E.; Chaurand, P.; Labille, J.; Rose, J.; Exbrayat, J.M.; Oughton, D.H.; Joner, E.J. Ecotoxicological effects of an aged TiO2 nanocomposite measured as apoptosis in the anecic earthworm Lumbricus terrestris after exposure through water, food and soil. Environ. Int. 2011, 37, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.; Cooper, E.L. The Role of Opsonins in Phagocytosis by Coelomocytes of the Earthworm, Lumbricus Terrestris. Dev. Comp. Immunol. 1981, 5, 415–425. [Google Scholar] [CrossRef]
- Hayashi, Y.; Miclaus, T.; Scavenius, C.; Kwiatkowska, K.; Sobota, A.; Engelmann, P.; Scott-Fordsmand, J.J.; Enghild, J.J.; Sutherland, D.S. Species Differences Take Shape at Nanoparticles: Protein Corona Made of the Native Repertoire Assists Cellular Interaction. Environ. Sci. Technol. 2013, 47, 14367–14375. [Google Scholar] [CrossRef] [PubMed]
- Brunelli, A.; Pojana, G.; Callegaro, S.; Marcomini, A. Agglomeration and sedimentation of titanium dioxide nanoparticles (n-TiO2) in synthetic and real waters. J. Nanoparticle Res. 2013, 15. [Google Scholar] [CrossRef]
- Alijagic, A.; Benada, O.; Kofronova, O.; Cigna, D.; Pinsino, A. Sea Urchin Extracellular Proteins Design a Complex Protein Corona on Titanium Dioxide Nanoparticle Surface Influencing Immune Cell Behavior. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Müllerová, I. Imaging of specimens at optimized low and very low energies in scanning electron microscopes. Scanning 2001, 23, 379–394. [Google Scholar] [CrossRef]
- Benada, O.; Pokorný, V. Modification of the Polaron sputter-coater unit for glow-discharge activation of carbon support films. J. Electron Microsc. Tech. 1990, 16, 235–239. [Google Scholar] [CrossRef]
- Semerad, J.; Cvancarova, M.; Filip, J.; Kaslik, J.; Zlota, J.; Soukupova, J.; Cajthaml, T. Novel assay for the toxicity evaluation of nanoscale zero-valent iron and derived nanomaterials based on lipid peroxidation in bacterial species. Chemosphere 2018, 213, 568–577. [Google Scholar] [CrossRef]
- Roubalova, R.; Prochazkova, P.; Hanc, A.; Dvorak, J.; Bilej, M. Mutual interactions of E. andrei earthworm and pathogens during the process of vermicomposting. Environ. Sci. Pollut. Res. Int. 2019, 10.1007/s11356-019-04329-5. [Google Scholar] [CrossRef]
- Magdolenova, Z.; Bilaničová, D.; Pojana, G.; Fjellsbø, L.M.; Hudecova, A.; Hasplova, K.; Marcomini, A.; Dusinska, M. Impact of agglomeration and different dispersions of titanium dioxide nanoparticles on the human related in vitro cytotoxicity and genotoxicity. J. Environ. Monit. 2012, 14, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Jin, X.; George, S.; Xia, T.; Meng, H.; Wang, X.; Suarez, E.; Zhang, H.; Hoek, E.M.V.; Godwin, H.A.; et al. Dispersion and Stability Optimization of TiO2 Nanoparticles in Cell Culture Media. Environ. Sci. Technol. 2010, 44, 7309–7314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-García, E.; Pérez-Arizti, J.A.; Márquez-Ramírez, S.G.; Delgado-Buenrostro, N.L.; Chirino, Y.I.; Iglesias, G.G.; López-Marure, R. Titanium dioxide nanoparticles induce strong oxidative stress and mitochondrial damage in glial cells. Free Radic. Biol. Med. 2014, 73, 84–94. [Google Scholar] [CrossRef]
- Semerad, J.; Moeder, M.; Filip, J.; Pivokonsky, M.; Filipova, A.; Cajthaml, T. Oxidative stress in microbes after exposure to iron nanoparticles: Analysis of aldehydes as oxidative damage products of lipids and proteins. Environ. Sci. Pollut. Res. 2019, 26, 33670–33682. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Velasco, N.; Gandariasbeitia, M.; Irizar, A.; Soto, M. Uptake route and resulting toxicity of silver nanoparticles in Eisenia fetida earthworm exposed through Standard OECD Tests. Ecotoxicology 2016, 25, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Semerad, J.; Pacheco, N.I.N.; Grasserova, A.; Prochazkova, P.; Pivokonsky, M.; Pivokonska, L.; Cajthaml, T. In Vitro Study of the Toxicity Mechanisms of Nanoscale Zero-Valent Iron (nZVI) and Released Iron Ions Using Earthworm Cells. Nanomaterials 2020, 10, 2189. [Google Scholar] [CrossRef] [PubMed]
- Homa, J.; Stalmach, M.; Wilczek, G.; Kolaczkowska, E. Effective activation of antioxidant system by immune-relevant factors reversely correlates with apoptosis of Eisenia andrei coelomocytes. J. Comp. Physiol. B 2016, 186, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Tumminello, R.A.; Fuller-Espie, S.L. Heat stress induces ROS production and histone phosphorylation in celomocytes of Eisenia hortensis. Invertebr. Surviv. J. 2013, 10, 50–57. [Google Scholar]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, L.; Wang, J.; Wang, J.; Du, Z.; Li, B.; Zhou, T.; Cheng, C.; Wang, Z. Evaluating subchronic toxicity of fluoxastrobin using earthworms (Eisenia fetida). Sci. Total Environ. 2018, 642, 567–573. [Google Scholar] [CrossRef]
- Poon, W.-L.; Lee, J.C.-Y.; Leung, K.S.; Alenius, H.; El-Nezami, H.; Karisola, P. Nanosized silver, but not titanium dioxide or zinc oxide, enhances oxidative stress and inflammatory response by inducing 5-HETE activation in THP-1 cells. Nanotoxicology 2020, 14, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Reeves, J.F.; Davies, S.J.; Dodd, N.J.F.; Jha, A.N. Hydroxyl radicals (OH) are associated with titanium dioxide (TiO2) nanoparticle-induced cytotoxicity and oxidative DNA damage in fish cells. Mutat. Res.-Fund. Mol. Mech. Mut. 2008, 640, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.W.; Li, M.; Cui, Y.B.; Li, D.S.; Chen, J.; Yang, L.Y. Toxicological effects of TiO2 and ZnO nanoparticles in soil on earthworm Eisenia fetida. Soil Biol. Biochem. 2010, 42, 586–591. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, X.; Liu, Y.; Zhang, J.; Lin, D. Integration of transcriptomics and metabolomics reveals the responses of earthworms to the long-term exposure of TiO2 nanoparticles in soil. Sci. Total Environ. 2020, 719, 137492. [Google Scholar] [CrossRef] [PubMed]
- Candas, D.; Li, J.J. MnSOD in oxidative stress response-potential regulation via mitochondrial protein influx. Antiox. Redox. Sign. 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, V.; Wilson, C.L.; Hayward, S.L.; Kidambi, S. Titanium Dioxide Nanoparticles Trigger Loss of Function and Perturbation of Mitochondrial Dynamics in Primary Hepatocytes. PLoS ONE 2015, 10, e0134541. [Google Scholar] [CrossRef]
- Bernard, F.; Brulle, F.; Douay, F.; Lemiere, S.; Demuynck, S.; Vandenbulcke, F. Metallic trace element body burdens and gene expression analysis of biomarker candidates in Eisenia fetida, using an “exposure/depuration” experimental scheme with field soils. Ecotoxicol. Environ. Saf. 2010, 73, 1034–1045. [Google Scholar] [CrossRef]
- Homa, J.; Zorska, A.; Wesolowski, D.; Chadzinska, M. Dermal exposure to immunostimulants induces changes in activity and proliferation of coelomocytes of Eisenia andrei. J. Comp. Physiol. B 2013, 183, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Bodó, K.; Ernszt, D.; Németh, P.; Engelmann, P. Distinct immune-and defense-related molecular fingerprints in sepatated coelomocyte subsets of Eisenia andrei earthworms. Invertebr. Surviv. J. 2018, 15, 338–345. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UV/Vis (nm) a | Z-Avg. (nm) b | ζ (mV) c | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2 h | 6 h | 24 h | 2 h | 6 h | 24 h | 2 h | 6 h | 24 h | |
| Distilled water | 300–370 | 300–370 | 300–370 | 581 ± 23.30 | 570 ± 2.75 | 480 ± 64.3 | −26.8 ± 2.99 | −31.7 ± 0.921 | −32.9 ± 2.59 |
| R-RPMI 1640 medium | 320–380 | 320–380 | 320–380 | 31.34 ± 1.55 | 35.5 ± 3.94 | 597 ± 447 | −16.9 ± 0.60 | −7.87 ± 0.631 | −5.94 ± 0.45 |
| Function | Gene | TiO2 NPs (µg/mL) | Normalized Gene Expression | ||
|---|---|---|---|---|---|
| 2 h | 6 h | 24 h | |||
| Metal detoxification | Metallothionein | 1 | 5.16 ± 1.73 ** | 2.00 ± 0.32 * | 2.71 ± 0.20 * |
| 10 | 1.11 ± 0.2 | 1.97 ± 0.22 ** | 1.00 ± 0.25 | ||
| Heavy metal detoxification | Phytochelatin | 1 | 1.38 ± 0.09 | 1.02 ± 0.04 | 1.18 ± 0.08 |
| 10 | 1.00 ± 0.02 | 0.82 ± 0.02 | 0.80 ± 0.01 | ||
| Oxidative stress | Mn-SOD | 1 | 1.47 ± 0.12 | 0.85 ± 0.19 | 0.58 ± 0.05 |
| 10 | 0.93 ± 0.09 | 0.53 ± 0.01 * | 0.72 ± 0.01 | ||
| CuZn-SOD | 1 | 0.68 ± 0.05 | 0.84 ± 0.22 | 0.98 ± 0.04 | |
| 10 | 0.96 ± 0.07 | 0.71 ± 0.04 | 0.87 ± 0.01 | ||
| Catalase | 1 | 1.41 ± 0.19 | 0.87 ± 0.03 | 0.66 ± 0.03 | |
| 10 | 1.04 ± 0.13 | 0.71 ± 0.02 | 0.8 ± 0.2 | ||
| Immunity | EMAP II | 1 | 0.90 ± 0.07 | 0.94 ± 0.1 | 0.86 ± 0.02 |
| 10 | 0.84 ± 0.09 | 1.21 ± 0.01 | 1.33 ± 0.20 | ||
| Fetidin/lysenin | 1 | 0.64 ± 0.08 | 0.62 ± 0.13 | 0.70 ± 0.04 | |
| 10 | 0.65 ± 0.05 | 2.20 ± 0.2 ** | 0.81 ± 0.19 | ||
| Lumbricin | 1 | 1.33 ± 0.05 | 0.75 ± 0.10 | 1.84 ± 0.02 | |
| 10 | 0.84 ± 0.10 | 2.10 ± 0.43 * | 1.92 ± 0.55 | ||
| Signal Transduction | MEKK I | 1 | 1.40 ± 0.19 | 1.47 ± 0.44 | 1.73 ± 0.04 * |
| 10 | 1.00 ± 0.15 | 1.96 ± 0.11 * | 1.33 ± 0.03 | ||
| PKC I | 1 | 1.52 ± 0.30 | 1.08 ± 0.19 | 1.43 ± 0.06 | |
| 10 | 1.10 ± 0.16 | 0.33 ± 0.04 ** | 0.58 ± 0.11 * | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro Pacheco, N.I.; Roubalova, R.; Semerad, J.; Grasserova, A.; Benada, O.; Kofronova, O.; Cajthaml, T.; Dvorak, J.; Bilej, M.; Prochazkova, P. In Vitro Interactions of TiO2 Nanoparticles with Earthworm Coelomocytes: Immunotoxicity Assessment. Nanomaterials 2021, 11, 250. https://doi.org/10.3390/nano11010250
Navarro Pacheco NI, Roubalova R, Semerad J, Grasserova A, Benada O, Kofronova O, Cajthaml T, Dvorak J, Bilej M, Prochazkova P. In Vitro Interactions of TiO2 Nanoparticles with Earthworm Coelomocytes: Immunotoxicity Assessment. Nanomaterials. 2021; 11(1):250. https://doi.org/10.3390/nano11010250
Chicago/Turabian StyleNavarro Pacheco, Natividad Isabel, Radka Roubalova, Jaroslav Semerad, Alena Grasserova, Oldrich Benada, Olga Kofronova, Tomas Cajthaml, Jiri Dvorak, Martin Bilej, and Petra Prochazkova. 2021. "In Vitro Interactions of TiO2 Nanoparticles with Earthworm Coelomocytes: Immunotoxicity Assessment" Nanomaterials 11, no. 1: 250. https://doi.org/10.3390/nano11010250
APA StyleNavarro Pacheco, N. I., Roubalova, R., Semerad, J., Grasserova, A., Benada, O., Kofronova, O., Cajthaml, T., Dvorak, J., Bilej, M., & Prochazkova, P. (2021). In Vitro Interactions of TiO2 Nanoparticles with Earthworm Coelomocytes: Immunotoxicity Assessment. Nanomaterials, 11(1), 250. https://doi.org/10.3390/nano11010250