
Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato

 ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Nanomaterial Characterization
2.3. Preparation of Sweet Potato Inoculant and Inoculation
2.4. Treatment Groups
2.5. SEM (Scanning Electron Microscope) Assessment
2.6. Determination of Hormonal Contents of Sweet Potato
2.7. Determination of Enzyme Activity of Sweet Potato
2.8. Biochemical Analysis
2.8.1. Total Flavone Content
2.8.2. Total Phenol Content
2.8.3. Ascorbic Acid Content
2.9. Data Analysis
3. Results and Discussion
3.1. NMs Characterization
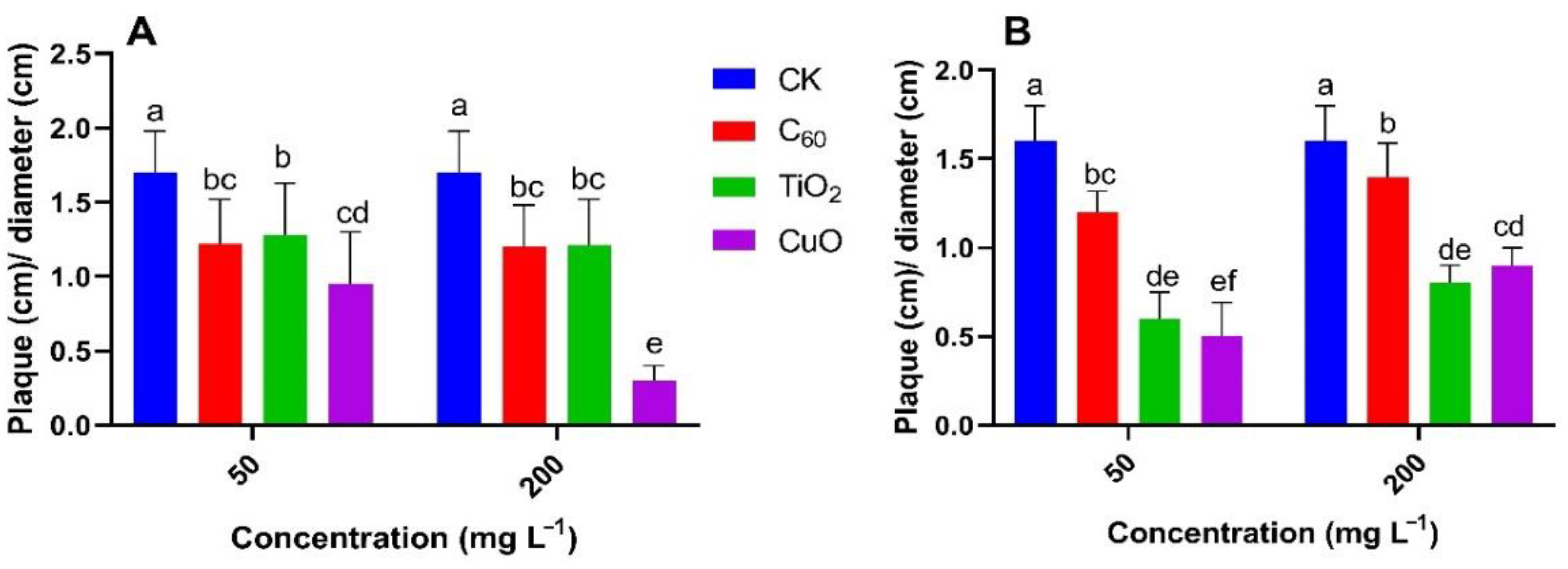
3.2. NMs Treatment Suppressed the Soft Rot Symptoms and Disease Development
3.3. NMs Application Enhances Hormonal Contents of Sweet Potato
3.4. Activation of the Antioxidant Activities in Sweet Potato over Application of NMs
3.5. Nutrients Content of Sweet Potatoes
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, W. Global pesticide use: Profile, trend, cost/benefit and more. Proc. Int. Acad. Ecol. Environ. Sci. 2018, 8, 1. [Google Scholar] [CrossRef]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Renu, S.; Abhilasha, A.L. Incidence of post-harvest fungal pathogens in guava and banana in allahabad. J. Hortic. Sci. 2009, 4, 85–89. [Google Scholar]
- Khalid, S.; Naseer, A.; Shahid, M.; Shah, G.M.; Ullah, M.I.; Waqar, A.; Abbas, T.; Imran, M.; Rehman, F. Assessment of nutritional loss with food waste and factors governing this waste at household level in Pakistan. J. Clean. Prod. 2019, 206, 1015–1024. [Google Scholar] [CrossRef]
- Abountiolas, M.; Kelly, K.; Yagiz, Y.; Li, Z.; Mahnken, G.; Borejsza-Wysocki, W.; Marshall, M.; Sims, C.A.; Peres, N.; do Nascimento Nunes, M.C.N. Sensory quality, physicochemical attributes, polyphenol profiles, and residual fungicides in strawberries from different disease-control treatments. J. Agric. Food Chem. 2018, 66, 6986–6996. [Google Scholar] [CrossRef] [PubMed]
- Jiaxin, L.; Yingli, M.; Yinfeng, X. Stimulatory Effect of Fe3O4 Nanoparticles on the growth and yield of pseudostellaria heterophylla via improved photosynthetic performance. HortScience 2021, 56, 753–761. [Google Scholar] [CrossRef]
- Xing, K.; Xing, Y.; Liu, Y.; Zhang, Y.; Shen, X.; Li, X.; Miao, X.; Feng, Z.; Peng, X.; Qin, S. Fungicidal effect of chitosan via inducing membrane disturbance against Ceratocystis fimbriata. Carbohydr. Polym. 2018, 192, 95–103. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Liu, Y.; Li, X.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Qin, S.; Xing, K. Volatile organic compounds produced by pseudomonas chlororaphis subsp. aureofaciens SPS-41 as biological fumigants to control ceratocystis fimbriata in postharvest sweet potatoes. J. Agric. Food Chem. 2019, 67, 3702–3710. [Google Scholar] [CrossRef]
- Rattner, B.A.; Lazarus, R.S.; Elliott, J.E.; Shore, R.F.; van den Brink, N.V.D. Adverse outcome pathway and risks of anticoagulant rodenticides to predatory wildlife. Environ. Sci. Technol. 2014, 48, 8433–8445. [Google Scholar] [CrossRef] [Green Version]
- Merel, S.; Benzing, S.; Gleiser, C.; Di Napoli-Davis, G.; Zwiener, C. Occurrence and overlooked sources of the biocide carbendazim in wastewater and surface water. Environ. Pollut. 2018, 239, 512–521. [Google Scholar] [CrossRef]
- Vallejo, A.; Millán, L.; Abrego, Z.; Sampedro, M.C.; Sánchez-Ortega, A.; Unceta, N.; Gómez-Caballero, A.; Goicolea, M.A.; Diez-Navajas, A.M.; Barrio, R.J. Fungicide distribution in vitiviniculture ecosystems according to different application strategies to reduce environmental impact. Sci. Total Environ. 2019, 687, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Flores, L.; Banjac, Z.; Farré, M.; Larrañaga, A.; Mas-Martí, E.; Muñoz, I.; Barceló, D.; Elosegi, A. Effects of a fungicide (imazalil) and an insecticide (diazinon) on stream fungi and invertebrates associated with litter breakdown. Sci. Total Environ. 2014, 476, 532–541. [Google Scholar] [CrossRef]
- de Souza, R.M.; Seibert, D.; Quesada, H.B.; de Jesus Bassetti, F.; Fagundes-Klen, M.R.; Bergamasco, R. Occurrence, impacts and general aspects of pesticides in surface water: A review. Process. Saf. Environ. Prot. 2020, 135, 22–37. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, M.; Pan, S.; Pan, C.; Li, Y.; Tian, J. Perillaldehyde controls postharvest black rot caused by Ceratocystis fimbriata in sweet potatoes. Front. Microbiol. 2018, 9, 1102. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, M.; Zafar, M.; Afsheen, S.; Iqbal, T. A review on ag-nanostructures for enhancement in shelf time of fruits. J. Inorg. Organomet. Polym. Mater. 2020, 30, 1475–1482. [Google Scholar] [CrossRef]
- Muñoz-Pina, S.; Ros-Lis, J.V.; Argüelles, Á.; Coll, C.; Martínez-Máñez, R.; Andrés, A. Full inhibition of enzymatic browning in the presence of thiol-functionalised silica nanomaterial. Food Chem. 2018, 241, 199–205. [Google Scholar] [CrossRef]
- Hao, Y.; Cao, X.; Ma, C.; Zhang, Z.; Zhao, N.; Ali, A.; Hou, T.; Xiang, Z.; Zhuang, J.; Wu, S.; et al. Potential applications and antifungal activities of engineered nanomaterials against gray mold disease agent botrytis cinerea on rose petals. Front. Plant. Sci. 2017, 8, 1332. [Google Scholar] [CrossRef] [Green Version]
- Chandra, H.; Patel, D.; Kumari, P.; Jangwan, J.S.; Yadav, S. Phyto-mediated synthesis of zinc oxide nanoparticles of Berberis aristata: Characterization, antioxidant activity and antibacterial activity with special reference to urinary tract pathogens. Mater. Sci. Eng. C 2019, 102, 212–220. [Google Scholar] [CrossRef]
- Xing, K.; Li, T.J.; Liu, Y.F.; Zhang, J.; Zhang, Y.; Shen, X.Q.; Li, X.Y.; Miao, X.M.; Feng, Z.Z.; Peng, X.; et al. Antifungal and eliciting properties of chitosan against Ceratocystis fimbriata in sweet potato. Food Chem. 2018, 268, 188–195. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Li, L.; Cheng, M.; Zhang, L. Optimization of konjac glucomannan/carrageenan/nano-SiO2 coatings for extending the shelf-life of Agaricus bisporus. Int. J. Biol. Macromol. 2019, 122, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Ruffo Roberto, S.; Youssef, K.; Hashim, A.F.; Ippolito, A. Nanomaterials as alternative control means against postharvest diseases in fruit crops. Nanomaterials 2019, 9, 1752. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Yuan, W.; Ma, C.; White, J.C.; Zhang, Z.; Adeel, M.; Zhou, T.; Rui, Y.; Xing, B. Engineered nanomaterials suppress turnip mosaic virus infection in tobacco (Nicotiana benthamiana). Environ. Sci. Nano 2018, 5, 1685–1693. [Google Scholar] [CrossRef]
- Adeel, M.; Farooq, T.; White, J.C.; Hao, Y.; He, Z.; Rui, Y. Carbon-based nanomaterials suppress tobacco mosaic virus (TMV) infection and induce resistance in Nicotiana benthamiana. J. Hazard. Mater. 2021, 404, 124167. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.; Adeel, M.; He, Z.; Umar, M.; Shakoor, N.; da Silva, W.; Elmer, W.; White, J.C.; Rui, Y. Nanotechnology and plant viruses: An emerging disease management approach for resistant pathogens. ACS Nano 2021, 15, 6030–6037. [Google Scholar] [CrossRef] [PubMed]
- Adu-Kwarteng, E.; Sakyi-Dawson, E.O.; Ayernor, G.S.; Truong, V.-D.; Shih, F.F.; Daigle, K. Variability of sugars in staple-type sweet potato (ipomoea batatas) cultivars: The effects of harvest time and storage. Int. J. Food Prop. 2014, 17, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.C.; Chen, C.-C.; Lin, K.-H.; Chao, P.-Y.; Lin, H.-H.; Huang, M.-Y. Bioactive compounds, antioxidants, and health benefits of sweet potato leaves. Molecules 2021, 26, 1820. [Google Scholar] [CrossRef]
- Abegunde, O.K.; Mu, T.-H.; Chen, J.-W.; Deng, F.-M. Physicochemical characterization of sweet potato starches popularly used in Chinese starch industry. Food Hydrocoll. 2013, 33, 169–177. [Google Scholar] [CrossRef]
- Oduola, A.A.; Awojobi, K.O.; Adeyemo, S.M. Microorganisms associated with post harvest spoilage of sweet potatoes in Ile-Ife, Nigeria. J. Microbiol. Res. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Yang, J.W.; Yu, G.D.; Nam, S.S.; Lee, S.Y.; Paul, N.C. First report of fusarium rot caused by Fusarium circinatum on sweet potato (Ipomoea batatas). J. Plant. Pathol. 2019, 101, 407. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N. Managing Postharvest Diseases: Classical versus New Technologies. In Bio-Management of Postharvest Diseases and Mycotoxigenic Fungi; CRC Press: Boca Raton, FL, USA, 2020; pp. 7–56. [Google Scholar]
- Edmunds, B.A.; Holmes, G.J. Evaluation of alternative decay control products for control of postharvest Rhizopus soft rot of sweetpotatoes. Plant Health Prog. 2009, 10, 26. [Google Scholar] [CrossRef]
- Hao, Y.; Ma, C.; Zhang, Z.; Song, Y.; Cao, W.; Guo, J.; Zhou, G.; Rui, Y.; Liu, L.; Xing, B. Carbon nanomaterials alter plant physiology and soil bacterial community composition in a rice-soil-bacterial ecosystem. Environ. Pollut. 2018, 232, 123–136. [Google Scholar] [CrossRef]
- Bodaghi, H.; Mostofi, Y.; Oromiehie, A.; Ghanbarzadeh, B.; Hagh, Z.G. Synthesis of clay–TiO2 nanocomposite thin films with barrier and photocatalytic properties for food packaging application. J. Appl. Polym. Sci. 2015, 132, 14. [Google Scholar] [CrossRef]
- Maneerat, C.; Hayata, Y. Antifungal activity of TiO2 photocatalysis against Penicillium expansum in vitro and in fruit tests. Int. J. Food Microbiol. 2006, 107, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.F.; Li, Q.; Li, X.Y.; Li, H.M.; Tang, Z.H.; Hu, L.; Cao, Q.H.; Xie, Y.P.; Wang, X. Selection of parents for breeding edible varieties of sweet potato with high carotene content. Agric. Sci. China 2009, 8, 1166–1173. [Google Scholar] [CrossRef]
- Ocsoy, I.; Paret, M.L.; Ocsoy, M.A.; Kunwar, S.; Chen, T.; You, M.; Tan, W. Nanotechnology in plant disease management: Dna-directed silver nanoparticles on graphene oxide as an antibacterial against xanthomonas perforans. ACS Nano 2013, 7, 8972–8980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-biotechnology in agriculture: Use of nanomaterials to promote plant growth and stress tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Liu, C.; Fan, G.; Liu, C.; Sun, X. Preventing viral disease by ZnONPs through directly deactivating TMV and activating plant immunity in nicotiana benthamiana. Environ. Sci. Nano 2019, 6, 3653–3669. [Google Scholar] [CrossRef]
- Nafady, N.A.; Alamri, S.A.M.; Hassan, E.A.; Hashem, M.; Mostafa, Y.S.; Abo-Elyousr, K.A.M. Application of ZnO-nanoparticles to manage Rhizopus soft rot of sweet potato and prolong shelf-life. Folia Hortic. 2019, 31, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Shende, S.; Bhagat, R.; Raut, R.; Rai, M.; Gade, A. Myco-fabrication of copper nanoparticles and its effect on crop pathogenic fungi. IEEE Trans. NanoBioscience 2021, 20, 146–153. [Google Scholar] [CrossRef]
- Ci, D.; Tian, M.; Song, Y.; Du, Q.; Quan, M.; Xuan, A.; Yu, J.; Yuan, Z.; Zhang, D. Indole-3-acetic acid has long-term effects on long non-coding RNA gene methylation and growth in Populus tomentosa. Mol. Genet. Genom. 2019, 294, 1511–1525. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D. Brassinosteroid signal transduction: From receptor kinase activation to transcriptional networks regulating plant development. Plant Cell. 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Chen, W.; Yao, Q.; Guo, B.; Valliyodan, B.; Wang, Z.; Nguyen, H.T. Genome-wide transcriptional profiling for elucidating the effects of brassinosteroids on Glycine max during early vegetative development. Sci. Rep. 2019, 9, 16085. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol. 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Roychoudhury, A. Melatonin application reduces fluoride uptake and toxicity in rice seedlings by altering abscisic acid, gibberellin, auxin and antioxidant homeostasis. Plant Physiol. Biochem. 2019, 145, 164–173. [Google Scholar] [CrossRef]
- Li, Z.; Yan, D.; Xue, Z.; Gu, Y.; Li, S.; Liu, L.; Zhang, H.; Wang, Z.; Yang, J.; Gu, J. Regulations of plant growth and development by cytokinins and their applications in rice production. Chin. J. Rice Sci. 2018, 32, 311–324. [Google Scholar] [CrossRef]
- Shang, H.; Ma, C.; Li, C.; Zhao, J.; Elmer, W.; White, J.C.; Xing, B. Copper oxide nanoparticle-embedded hydrogels enhance nutrient supply and growth of lettuce (lactuca sativa) infected with Fusarium oxysporum f. sp. lactucae. Environ. Sci. Technol. 2021, in press. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Yang, Y. PEG 6000-stimulated drought stress improves the attributes of in vitro growth, steviol glycosides production, and antioxidant activities in stevia rebaudiana bertoni. Plants 2020, 9, 1552. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Ao, Q.; Yang, Y. Engineered ZnO and CuO nanoparticles ameliorate morphological and biochemical response in tissue culture regenerants of candyleaf (Stevia rebaudiana). Molecules 2020, 25, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokerst, J.C.; Adkins, J.A.; Bisha, B.; Mentele, M.M.; Goodridge, L.D.; Henry, C.S. Development of a paper-based analytical device for colorimetric detection of select foodborne pathogens. Anal. Chem. 2012, 84, 2900–2907. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.F.; Xu, Y.J.; Lai, X.H.; Chang, J.L.; Peng, D.H. Effects of endophytic bacteria 252 and 254 on peroxidase (POD) and catalase (CAT) activities of wheat seedlings under salt stress. J. Appl. Ecol. 2018, 28, 2984–2992. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [Green Version]
- Sha, W.; Jiang, Z.; Zhang, M.; Ma, T. Cloning and expression analysis of drought-resistance related gene RcSOD in racomitrium canescens. Genom. Appl. Biol. 2019, 38, 2144–2152. [Google Scholar] [CrossRef]
- Song, G.; Gao, Y.; Wu, H.; Hou, W.; Zhang, C.; Ma, H. Physiological effect of anatase TiO2 nanoparticles on lemna minor. Environ. Toxicol. Chem. 2012, 31, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Miura, C.; Sugawara, K.; Neriya, Y.; Minato, N.; Keima, T.; Himeno, M.; Maejima, K.; Komatsu, K.; Yamaji, Y.; Oshima, K.; et al. Functional characterization and gene expression profiling of superoxide dismutase from plant pathogenic phytoplasma. Gene 2012, 510, 107–112. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef]
- Thangavelu, R.M.; Gunasekaran, D.; Jesse, M.I.; S.U., M.R.; Sundarajan, D.; Krishnan, K. Nanobiotechnology approach using plant rooting hormone synthesized silver nanoparticle as “nanobullets” for the dynamic applications in horticulture—An in vitro and ex vitro study. Arab. J. Chem. 2018, 11, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Alghuthaymi, M.; Abd-Elsalam, K.A.; Paraliker, P.; Rai, M. Mono and hybrid nanomaterials: Novel strategies to manage postharvest diseases. Multifunct. Hybrid Nanomater. Sustain. Agric.-Food Ecosyst. 2020, 13, 287–317. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Packer, J.E.; Slater, T.F.; Willson, R.L. Direct observation of a free radical interaction between vitamin E and vitamin C. Nature 1979, 278, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Osmond, C.B.; Foyer, C.H.; Bock, G.; Smirnoff, N. Ascorbate biosynthesis and function in photoprotection. Philos. Trans. R. Soc. B 2000, 355, 1455–1464. [Google Scholar] [CrossRef]
- Pignocchi, C.; Foyer, C.H. Apoplastic ascorbate metabolism and its role in the regulation of cell signalling. Curr. Opin. Plant Biol. 2003, 6, 379–389. [Google Scholar] [CrossRef]
- Kumar, V.; Rani, A.; Dixit, A.K.; Pratap, D.; Bhatnagar, D. A comparative assessment of total phenolic content, ferric reducing-anti-oxidative power, free radical-scavenging activity, vitamin C and isoflavones content in soybean with varying seed coat colour. Food Res. Int. 2010, 43, 323–328. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Leonard, S.W.; Traber, M.G.; Reed, B.M. Vitamins C and E improve regrowth and reduce lipid peroxidation of blackberry shoot tips following cryopreservation. Plant Cell Rep. 2009, 29, 25. [Google Scholar] [CrossRef]
- Miyake, C.; Asada, K. Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxidation product monodehydroascorbate radicals in thylakoids. Plant Cell Physiol. 1992, 33, 541–553. [Google Scholar] [CrossRef]
- Polívka, T.; Frank, H.A. Molecular factors controlling photosynthetic light harvesting by carotenoids. Acc. Chem. Res. 2010, 43, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Yoneyama, K.; Yoneyama, K. The strigolactone story. Annu. Rev. Phytopathol. 2010, 48, 93–117. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.D.; Hossain, M.A. Total flavonoids content and biochemical screening of the leaves of tropical endemic medicinal plant Merremia borneensis. Arab. J. Chem. 2014, 7, 1034–1038. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, L.-J.; Adeel, M.; Shakoor, N.; Guo, K.-R.; Ma, D.-F.; Ahmad, M.A.; Lu, G.-Q.; Zhao, M.-H.; Li, S.-E.; Rui, Y.-K. Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato. Nanomaterials 2021, 11, 2572. https://doi.org/10.3390/nano11102572
Pang L-J, Adeel M, Shakoor N, Guo K-R, Ma D-F, Ahmad MA, Lu G-Q, Zhao M-H, Li S-E, Rui Y-K. Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato. Nanomaterials. 2021; 11(10):2572. https://doi.org/10.3390/nano11102572
Chicago/Turabian StylePang, Lin-Jiang, Muhammed Adeel, Noman Shakoor, Ke-Rui Guo, Dai-Fu Ma, Muhammad Arslan Ahmad, Guo-Quan Lu, Mei-Hui Zhao, Sheng-E Li, and Yu-Kui Rui. 2021. "Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato" Nanomaterials 11, no. 10: 2572. https://doi.org/10.3390/nano11102572
APA StylePang, L. -J., Adeel, M., Shakoor, N., Guo, K. -R., Ma, D. -F., Ahmad, M. A., Lu, G. -Q., Zhao, M. -H., Li, S. -E., & Rui, Y. -K. (2021). Engineered Nanomaterials Suppress the Soft Rot Disease (Rhizopus stolonifer) and Slow Down the Loss of Nutrient in Sweet Potato. Nanomaterials, 11(10), 2572. https://doi.org/10.3390/nano11102572