
Cellular Responses Induced by Zinc in Zebra Mussel Haemocytes. Loss of DNA Integrity as a Cellular Mechanism to Evaluate the Suitability of Nanocellulose-Based Materials in Nanoremediation


 , ,
, ,  , , ,
, , ,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Equipments
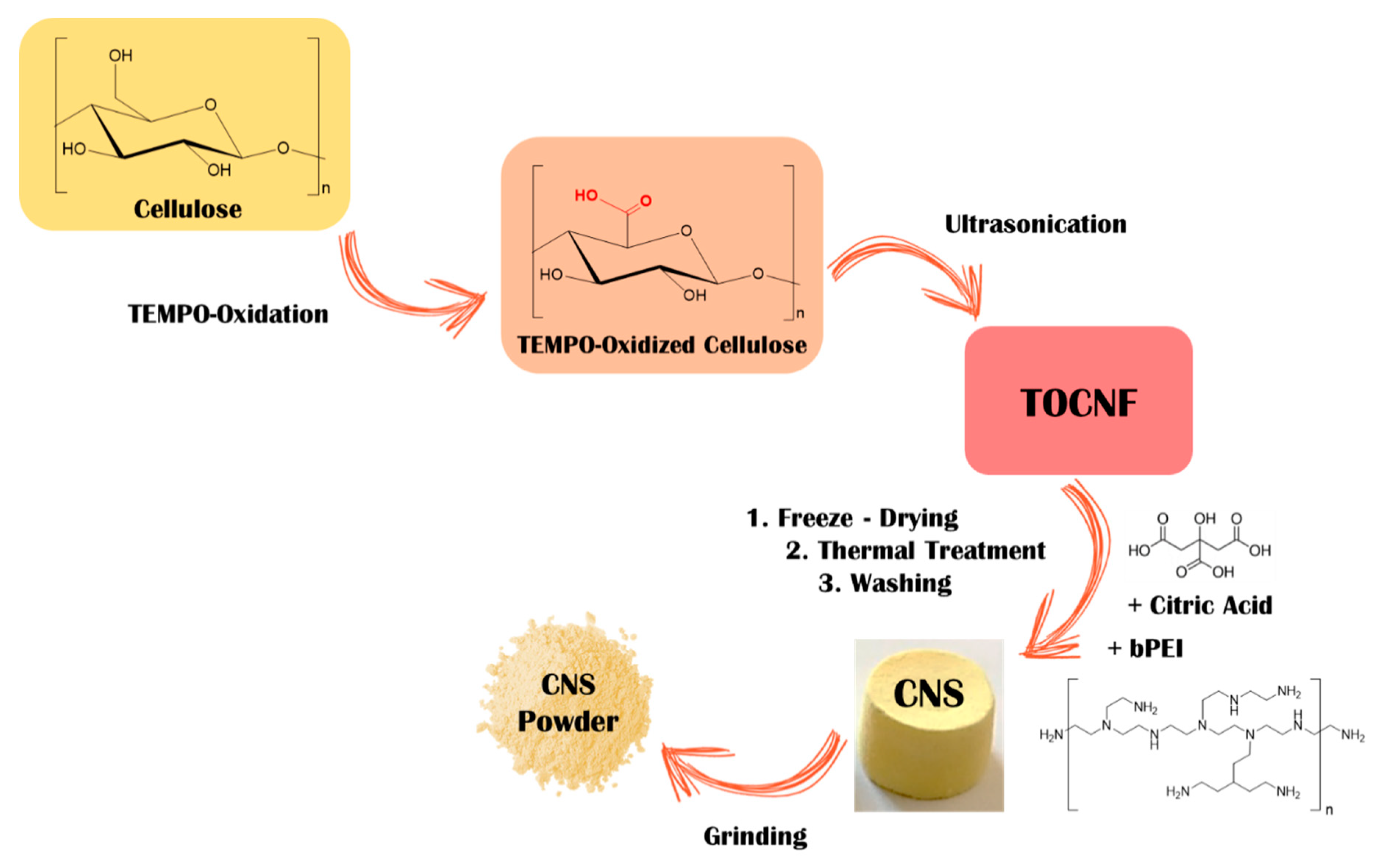
2.2. Preparation and Characterization of the Cellulose-Based Nanosponge (CNS)
2.2.1. Synthesis of TEMPO-Oxidized Cellulose
2.2.2. Synthesis of CNS
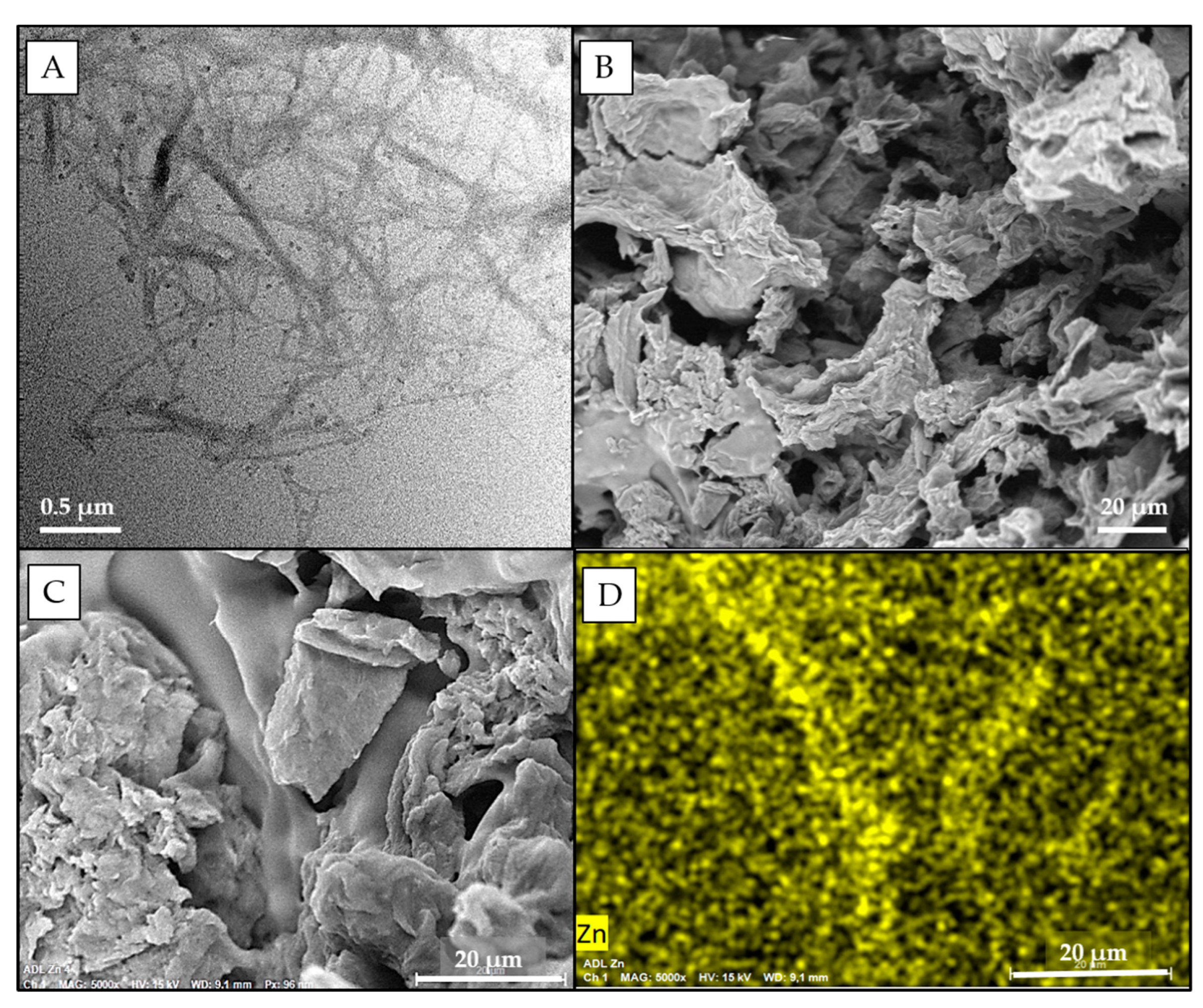
2.2.3. Characterization of TOCNF and CNS
2.3. Sampling and Maintenance Condition
2.4. In Vivo Exposure
2.5. Zinc Concentration in Water
2.6. Zn Concentrations in CNS and Zebra Mussel
2.7. Viability Assessment
2.8. Comet Assay
2.9. Cytome Assay
2.10. Acute Toxicity Test with Aliivibrio Fischeri
2.11. Statistical Analysis
3. Results and Discussion
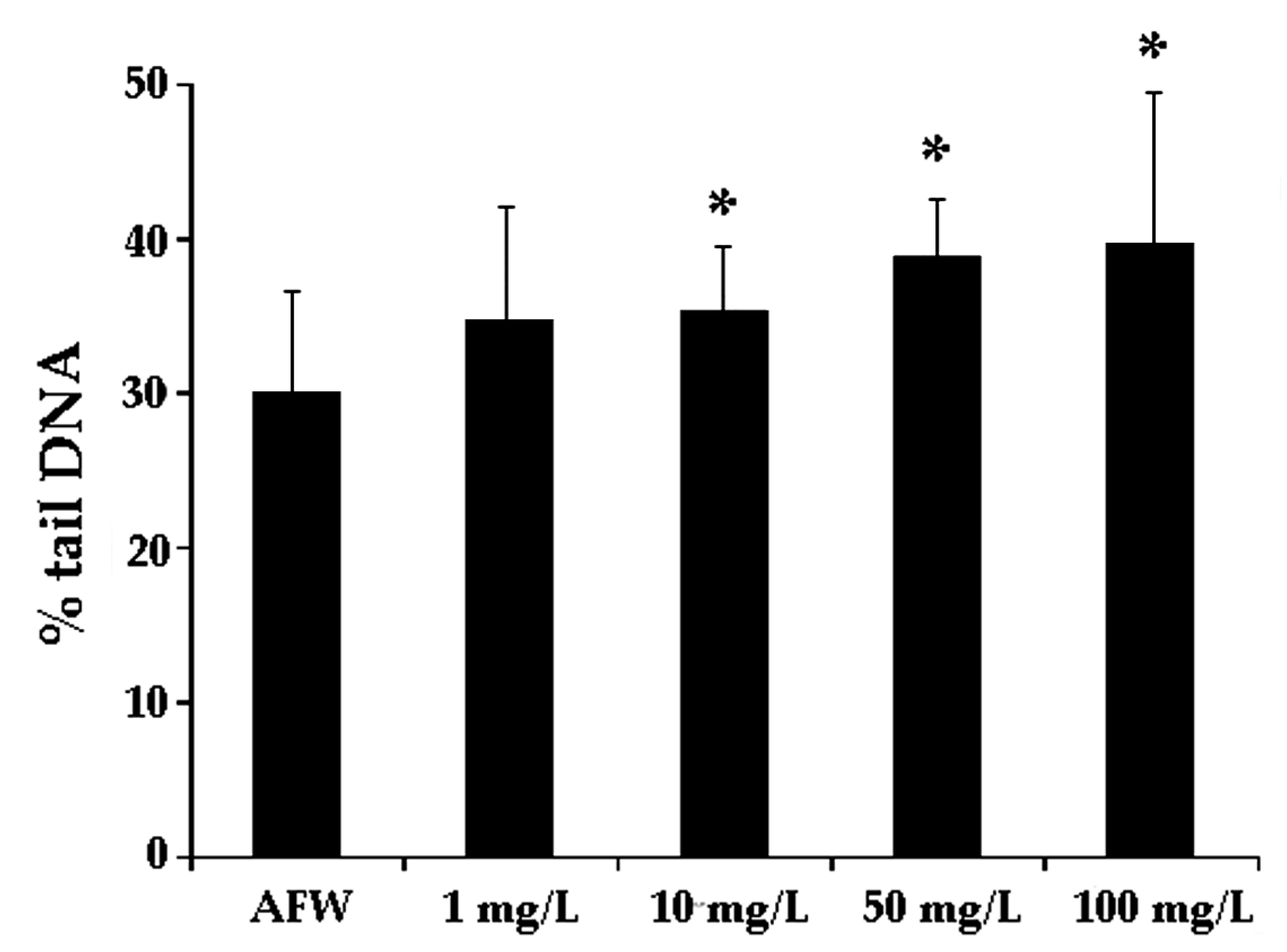
3.1. Dose-Effect Experiments
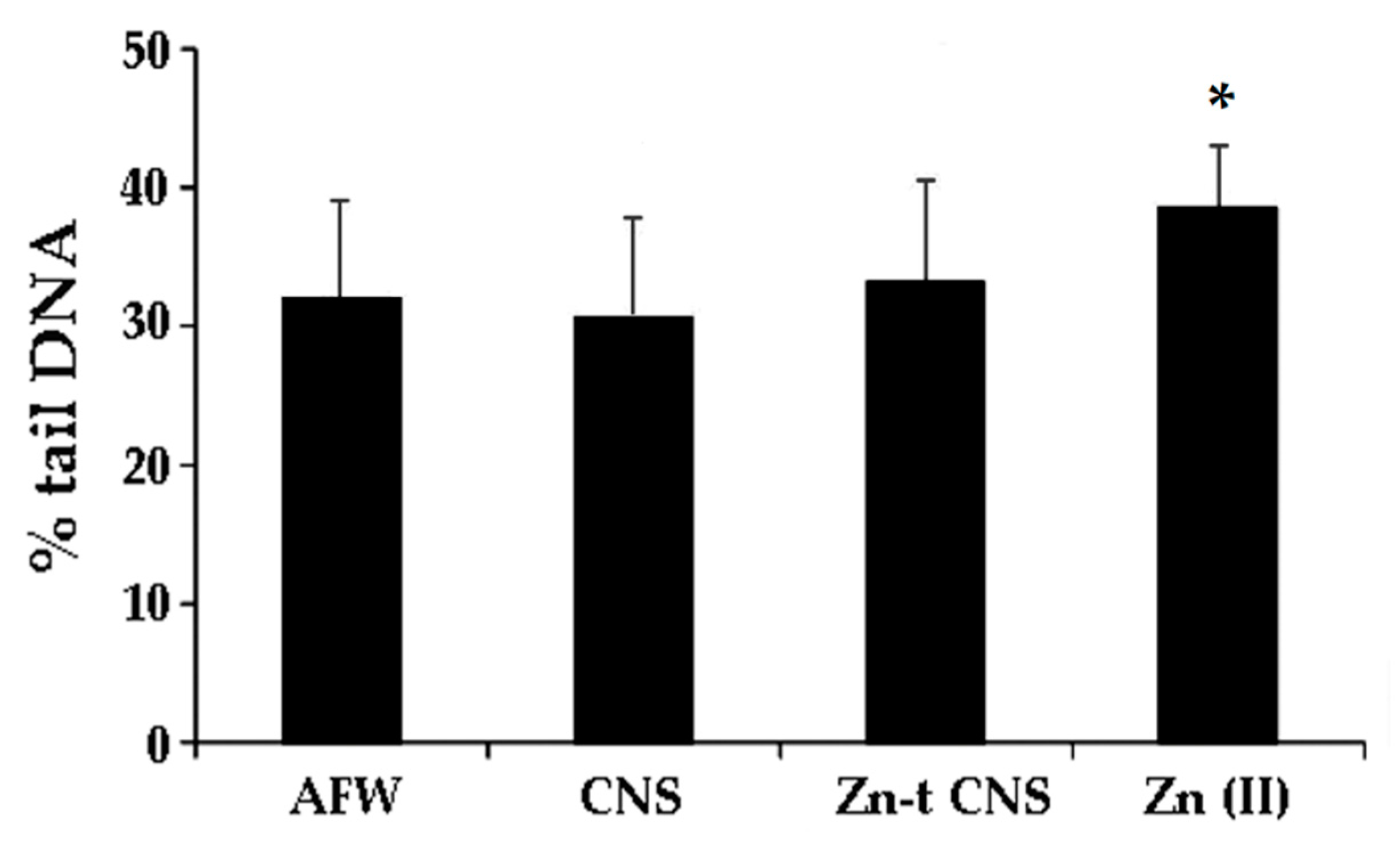
3.2. Combined Experiments
3.2.1. Synthesis and Characterization of Cellulose Nanosponges
3.2.2. Zn(II) Adsorption Efficiency of CNS
3.2.3. Genotoxicity
3.2.4. Acute Toxicity Test with Aliivibrio fischeri
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Zinc Association. Available online: https://www.zinc.org/environment/ (accessed on 29 June 2021).
- Jha, A.N.; Cheung, V.V.; Foulkes, M.E.; Hill, S.J.; Depledge, M.H. Detection of genotoxins in the marine environment: Adoption and evaluation of an integrated approach using the embryo-larval stages of the marine mussel Mytilus edulis. Mutat. Res. 2000, 464, 213–228. [Google Scholar] [CrossRef]
- Frenzilli, G. Nanotechnology for Environmental and Biomedical Research. Nanomaterials 2020, 10, 2220. [Google Scholar] [CrossRef]
- Karn, B.; Kuiken, T.; Otto, M. Nanotechnology and in situ remediation: A review of the benefits and potential risks. Environ. Health Perspect. 2009, 117, 1813–1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M.; Floyd, M.; Bajpai, S. Nanotechnology for site remediation. Remediation 2008, 19, 99–108. [Google Scholar] [CrossRef]
- Corsi, I.; Winther-Nielsen, M.; Sethi, R.; Punta, C.; Della Torre, C.; Libralato, G.; Lofrano, G.; Sabatini, L.; Aiello, M.; Fiordi, L.; et al. Ecofriendly nanotechnologies and nanomaterials for environmental applications: Key issue and consensus recommendations for sustainable and ecosafe nanoremediation. Ecotoxicol. Environ. Saf. 2018, 154, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Bennett, S.; Keller, A.A.; Pease, S.; Lenihan, H.S. TiO2 Nanoparticles Are Phototoxic to Marine Phytoplankton. PLoS ONE 2012, 7, e30321. [Google Scholar] [CrossRef] [Green Version]
- Mathur, A.; Parashar, A.; Chandrasekaran, N.; Mukherjee, A. Nano-TiO2 enhances biofilm formation in a bacterial isolate from activated sludge of a waste water treatment plant. Int. Biodeterior. Biodegrad. 2017, 116, 17–25. [Google Scholar] [CrossRef]
- Aruoja, V.; Dubourguier, H.C.; Kasemets, K.; Kahru, A. Toxicity of nanoparticles of CuO, ZnO and TiO2 to microalgae Pseudokirchneriella subcapitata. Sci. Total Environ. 2009, 407, 1461–1468. [Google Scholar] [CrossRef]
- Mortimer, M.; Kasemets, K.; Kahru, A. Toxicity of ZnO and CuO nanoparticles to ciliated protozoa Tetrahymena thermophila. Toxicology 2010, 269, 182–189. [Google Scholar] [CrossRef]
- Hanna, S.K.; Miller, R.J.; Lenihan, H.S. Deposition of carbon nanotubes by a marine suspension feeder revealed by chemical and isotopic tracers. J. Hazard. Mater. 2014, 279, 32–37. [Google Scholar] [CrossRef]
- Callaghan, N.I.; MacCormack, T.J. Ecophysiological perspectives on engineered nanomaterial toxicity in fish and crustaceans. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 193, 30–41. [Google Scholar] [CrossRef]
- Oughton, D.; Koin, P.; Bleyl, S.; Filip, J.; Skácelová, P.; Klaas, N.; Von der Kammer, F.; Gondikas, A. Development and Application of Analytical Methods for Monitoring Nanoparticles in Remdation: NanoRem 5 CLAIRE; London, UK, 2017; Volume 6, pp. 32–44. Available online: file:///C:/Users/amministratore/Downloads/NanoRem_TB_DL_6-2_monitoring_analytical_toolbox.pdf (accessed on 23 August 2021).
- Carpenter, A.W.; de Lannoy, C.-F.; Wiesner, M.R. Cellulose nanomaterials in water treatment technologies. Environ. Sci. Technol. 2015, 49, 5277–5287. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-M.; Ting, S.S.; Ong, H.L.; Omar, M. Surface functionalized nanocellulose as a veritable inclusionary material in contemporary bioinspired applications: A review. J. Appl. Polym. Sci. 2018, 135, 46065. [Google Scholar] [CrossRef] [Green Version]
- Bossa, N.; Carpenter, A.W.; Kumar, N.; de Lannoy, C.-F.; Wiesner, M. Cellulose nanocrystal zero-valent iron nanocomposites for groundwater remediation. Environ. Sci. Nano 2017, 4, 1294–1303. [Google Scholar] [CrossRef]
- Fiorati, A.; Grassi, G.; Graziano, A.; Liberatori, G.; Pastori, N.; Melone, L.; Bonciani, L.; Pontorno, L.; Punta, C.; Corsi, I. Eco-design of nanostructured cellulose sponges for sea-water decontamination from heavy metal ions. J. Clean. Prod. 2020, 246. [Google Scholar] [CrossRef]
- Liberatori, G.; Grassi, G.; Guidi, P.; Bernardeschi, M.; Fiorati, A.; Scarcelli, V.; Genovese, M.; Faleri, C.; Protano, G.; Frenzilli, G.; et al. Effect-Based Approach to Assess Nanostructured Cellulose Sponge Removal Efficacyof Zinc Ions from Seawater to Prevent Ecological Riscks. Nanomaterial 2020, 10, 1283. [Google Scholar] [CrossRef]
- Guidi, P.; Bernardeschi, M.; Palumbo, M.; Genovese, M.; Scarcelli, V.; Fiorati, A.; Riva, L.; Punta, C.; Corsi, I.; Frenzilli, G. Suitability of a Cellulose-Based Nanomaterial for the Remediation of Heavy Metal Contaminated Freshwaters: A Case-Study Showing the Recovery of Cadmium Induced DNA Integrity Loss, Cell Proliferation Increase, Nuclear Morphology and Chromosomal Alterations on Dreissena polymorpha. Nanomaterials 2020, 10, 1837. [Google Scholar] [CrossRef]
- Corsi, I.; Fiorati, A.; Grassi, G.; Bartolozzi, I.; Daddi, T.; Melone, L.; Punta, C. Environmentally Sustainable and Ecosafe Polysaccharide-Based Materials for Water Nano-Treatment: An Eco-Design Study. Materials 2018, 11, 1228. [Google Scholar] [CrossRef] [Green Version]
- Riva, L.; Pastori, N.; Panozzo, A.; Antonelli, M.; Punta, C. Nanostructured Cellulose-Based Sorbent Materials for Water Decontamination from Organic Dyes. Nanomaterials 2020, 10, 1570. [Google Scholar] [CrossRef]
- Fiorati, A.; Turco, G.; Travan, A.; Caneva, E.; Pastori, N.; Cametti, M.; Punta, C.; Melone, L. Mechanical and Drug Release Properties of Sponges from Cross-linked Cellulose Nanofibers. ChemPlusChem 2017, 82, 848–858. [Google Scholar] [CrossRef]
- Melone, L.; Rossi, B.; Pastori, N.; Panzeri, W.; Mele, A.; Punta, C. TEMPO-Oxidized Cellulose Cross-Linked with Branched Polyethyleneimine: Nanostructured Adsorbent Sponges for Water Remediation. Chempluschem 2015, 80, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Isogai, A.; Saitoa, T.; Fukuzumia, H. TEMPO-oxidized cellulose nanofibers. Nanoscale 2011, 3, 71–85. [Google Scholar] [CrossRef]
- Pierre, G.; Punta, C.; Delattre, C.; Melone, L.; Dubessay, P.; Fiorati, A.; Pastori, N.; Galante, Y.M.; Michaud, P. TEMPO-mediated oxidation of polysaccharides: An ongoing story. Carbohyd. Polym. 2017, 165, 71–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo Stampino, P.; Riva, L.; Punta, C.; Elegir, G.; Bussini, D.; Dotelli, G. Comparative Life Cycle Assessment of Cellulose Nanofibres Production Routes from Virgin and Recycled Raw Materials. Molecules 2021, 26, 2558. [Google Scholar] [CrossRef] [PubMed]
- Binelli, A.; Cogni, D.; Parolini, M.; Riva, C.; Provini, A. In vivo experiments for the evaluation of genotoxic and cytotoxic effects of Triclosan in Zebra mussel hemocytes. Aquat. Toxicol. 2009, 91, 238–244. [Google Scholar] [CrossRef]
- Hietanen, B.; Sunila, I.; Kristoff, R. Toxic effects of zinc on the common mussel Mytilus edulis L. (Bivalvia) in brackish water. I. Physiological and histopathological studies. Ann. Zool. Fenn. 1988, 25, 341–347. [Google Scholar]
- Ministry of Foreign Affairs and International Cooperation. Available online: https://www.esteri.it/mae/doc/prontuario_navigazione_2016/documenti_prontuario/11.normativa%20sanitaria%20e%20presidi%20medici%20di%20bordo/decreto%20del%20presidente%20della%20repubblica%2024%20maggio%201988%20n.%20236.pdf (accessed on 29 June 2021).
- Official Gazette of Italian Republic. Available online: https://www.gazzettaufficiale.it/eli/id/1999/10/01/099G0410/sg (accessed on 29 June 2021).
- Palacio, S.M.; Espinoza-Quiñones, F.R.; Manozzo Galante, R.; Zenatti, D.C.; Seolatto, A.A.; Lorenz, E.K.; Zacarkim, C.E.; Rossi, N.; Rizzutto, M.; Tabacniks, M.H. Correlation between Heavy Metal Ions (Copper, Zinc, Lead) Concentrations and Root Length of Allium cepa L. in Polluted River Water. Braz. Arch. Biol. Technol. 2005, 48, 191–196. [Google Scholar] [CrossRef]
- Matos, L.A.; Cunha, A.C.S.; Sousa, A.A.; Maranhão, J.P.R.; Santos, N.R.S.; Gonçalves, M.d.M.C.; Dantas, S.M.M.d.M.; e Sousa, J.M.d.C.; Peron, A.P.; da Silva, F.C.C.; et al. The influence of heavy metals on toxicogenetic damage in a Brazilian tropical river. Chemosphere 2017, 185, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Guidi, P.; Bernardeschi, M.; Scarcelli, V.; Cantafora, E.; Beneteddi, M.; Falleni, A.; Frenzilli, G. Lysosomal, genetic and chromosomal damage in haemocytes of the freshwater bivalve (Unio pictorum) exposed to polluted sediments from the River Cecina (Italy). Chem. Ecol. 2017, 33, 516–527. [Google Scholar] [CrossRef]
- Spielmann, H.; Balls, M.; Dupuis, J.; Pape, W.J.; Pechovitch, G.; De Silva, O.; Holzhutter, H.G.; Clothier, R.; Desolle, P.; Gerberick, F.; et al. The International EU/COLIPA in vitro phototoxicity validation study: Results of phase II (Blind Trial). Part 1: The 3T3 NRU phototoxicity test. Toxicol. In Vitro 1998, 12, 305–327. [Google Scholar] [CrossRef]
- Guidi, P.; Frenzilli, G.; Benedetti, M.; Bernardeschi, M.; Falleni, A.; Fattorini, D.; Regoli, F.; Scarcelli, V.; Nigro, M. Antioxidant, genotoxic and lysosomal biomarkers in the freshwater bivalve (Unio pictorum) transplanted in a metal polluted river basin. Aquat. Toxicol. 2010, 100, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Kumaravel, T.S.; Jha, A.N. Reliable Comet assay measurements for detecting DNA damage induced by ionising radiation and chemicals. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2006, 605, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO 11348-1:2007 Water Quality. Available online: https://www.iso.org/obp/ui/#iso:std:iso:11348:-2:ed-2:v1:en (accessed on 29 June 2021).
- Isprambiente.gov.it. Available online: https://www.isprambiente.gov.it/contentfiles/00006700/6770-manuale-apat-icram-2007.pdf (accessed on 29 June 2021).
- Lovell, D.P.; Omori, T. Statistical issues in the use of the comet assay. Mutagenesis 2008, 23, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Nacci, D.E.; Cayula, S.; Jackim, E. Detection of DNA damage in individual cells from marine organisms using the single cell gel assay. Aquat. Toxicol. 1996, 35, 197–210. [Google Scholar] [CrossRef]
- Buschini, A.; Carboni, P.; Martino, A.; Poli, P.; Ross, C. Effects of temperature on baseline and genotoxicant-induced DNA damage in haemocytes of Dreissena polymorpha. Mutat. Res. 2003, 537, 81–92. [Google Scholar] [CrossRef]
- Mersch, J.; Beauvais, M.-N.; Nagel, P. Induction of micronuclei in haemocytes and gill cells of zebra mussels, Dreissena polymorpha, exposed to clastogens. Mutat. Res. Genet. Toxicol. 1996, 371, 47–55. [Google Scholar] [CrossRef]
- Pavlica, M.; Klobučar, G.I.V.; Mojaš, N.; Erben, R.; Papeš, D. Detection of DNA damage in haemocytes of zebra mussel using comet assay. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 490, 209–214. [Google Scholar] [CrossRef]
- Banu, B.S.; Danadevi, K.; Rahman, M.F.; Ahuja, Y.R.; Kaiser, J. Genotoxic effect of monocrotophos to sentinel species using the Comet assay. Food Chem. Toxicol. 2001, 39, 361–366. [Google Scholar] [CrossRef]
- Farombi, E.O.; Adelowo, O.A.; Ajimoko, Y.R. Biomarkers of oxidative stress and heavy metal levels as indicators of environmental pollution in African catfish (Clarias gariepinus) from Nigeria Ogun River. Int. J. Environ. Res. Public Health 2007, 4, 158–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalon, M.C.S.; Rechenmacher, C.; Siebel, A.M.; Kayser, M.L.; Rodrigues, M.T.; Maluf, S.W.; Rodrigues, M.A.S.; Silva, L.B. Evaluation of Sinos River water genotoxicity using the comet assay in fish. Braz. J. Biol. 2010, 70, 1217–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.F.; Chen, S.Y.; Qu, A.O.; Adeleye, A.O.; Di, Y.N. Utilization of isolated marine mussel cells as an in vitro model to assess xenobiotics induced genotoxicity. Toxicol. In Vitro 2017, 44, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Pearson, H.B.; Dallas, L.J.; Comber, S.D.W.; Braungardt, C.B.; Worsfold, P.J.; Jha, A.N. Mixtures of tritiated water, zinc and dissolved organic carbon: Assessing interactive bioaccumulation and genotoxic effects in marine mussels, Mytilus galloprovincialis. J. Environ. Radioact. 2018, 187, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Bagdonas, E.; Vosylienė, M.Z. A study of toxicity and genotoxicity of copper, zinc and their mixture to rainbow trout (Oncorhynchus mykiss). Biologija 2006, 1, 8–13. [Google Scholar]
- Majone, F.; Beltrame, C.; Brunetti, R. Frequencies of micronuclei detected on Mytilus galloprovincialis by different staining techniques after treatment with zinc chloride. Mutat. Res. 1988, 209, 131–134. [Google Scholar] [CrossRef]
- Fernández, B.; Campillo, J.A.; Martínez-Gómez, C.; Benedicto, J. Micronuclei and other nuclear abnormalities in mussels (Mytilus galloprovincialis) as biomarkers of cyto-genotoxic pollution in mediterranean waters. Environ. Mol. Mutagen. 2011, 52, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Varotto, L.; Domeneghetti, S.; Rosani, U.; Manfrin, C.; Cajaraville, M.P.; Raccanelli, S.; Pallavicini, A.; Venier, P. DNA Damage and Transcriptional Changes in the Gills of Mytilus galloprovincialis Exposed to Nanomolar Doses of Combined Metal Salts (Cd, Cu, Hg). PLoS ONE 2013, 8, e54602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binelli, A.; Pedriali, A.; Riva, C.; Parolini, M. Illicit drugs as new environmental pollutants: Cyto-genotoxic effects of cocaine on the biological model Dreissena polymorpha. Chemosphere 2012, 86, 906–911. [Google Scholar] [CrossRef]
- Kononenko, V.; Repar, N.; Marušič, N.; Drašler, B.; Romih, T.; Hočevar, S.; Drobne, D. Comparative in vitro genotoxicity study of ZnO nanoparticles, ZnO macroparticles and ZnCl2 to MDCK kidney cells: Size matters. Toxicol. In Vitro 2017, 40, 256–263. [Google Scholar] [CrossRef]
- Paladini, G.; Venuti, V.; Almásy, L.; Melone, L.; Crupi, V.; Majolino, D.; Pastori, N.; Fiorati, A.; Punta, C. Cross-linked cellulose nano-sponges: A small angle neutron scattering (SANS) study. Cellulose 2019, 26, 9005–9019. [Google Scholar] [CrossRef]
- Paladini, G.; Venuti, V.; Crupi, V.; Majolino, D.; Fiorati, A.; Punta, C. FTIR-ATR analysis of the H-bond network of water in branched polyethyleneimine/TEMPO-oxidized cellulose nano-fiber xerogels. Cellulose 2020, 27, 8605–8618. [Google Scholar] [CrossRef]
- Kraak, M.H.; Lavy, D.; Schoon, H.; Toussaint, M.; Peeters, W.H.; van Straalen, N.M. Ecotoxicity of mixtures of metals to the zebra mussel Dreissena polymorpha. Environ. Toxicol. Chem. 1994, 13, 109–114. [Google Scholar] [CrossRef]
- Klerks, P.L.; Fraleigh, P.C. Uptake of nickel and zinc by the zebra mussel Dreissena polymorpha. Arch. Environ. Contam. Toxicol. 1997, 32, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.S.; White, S.L. Comparative strategies of heavy metal accumulation by crustaceans: Zinc, copper and cadmium in a decapod, an amphipod and a barnacle. Hydrobiologia 1989, 174, 245–262. [Google Scholar] [CrossRef]
- Sargsyan, A.; Simonyan, A.; Hovhannisyan, G.; Arakelyan, M.; Aroutiounian, R. Application of the comet assay, micronucleus test and global DNA methylation analysis in Darevskia lizards as a sentinel organism for genotoxic monitoring of soil pollution. Mutat. Res./Genet. Toxicol Environ. Mutagen. 2019, 842, 117–124. [Google Scholar] [CrossRef]
- Stoudmann, N.; Schmutz, M.; Hirsch, C.; Nowack, B.; Som, C. Human hazard potential of nanocellulose: Quantitative insights from the literature. Nanotoxicology 2020, 14, 1241–1257. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, L.; Lu, Y.; Long, Y.; Wang, L.; Ho, K.-P.; Wong, K.-Y. Rapid determination of nanotoxicity using luminous bacteria. Anal. Sci. 2010, 26, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, K.L.E.; Devillers, J. (Eds.) Ecotoxicology of Chemicals to Photobacterium phosphoreum; Gordon & Breach Science: Langhorne, PA, USA, 1994. [Google Scholar]
- Kong, J.C.; Bitton, G.; Koopman, B.; Jung, K.H. Heavy metal toxicity testing in environmental samples. Rev. Environ. Contam. Toxicol. 1995, 142, 119–147. [Google Scholar] [CrossRef]
- Casado, M.P.; Macken, A.; Byrne, H.J. Ecotoxicological assessment of silica and polystyrene nanoparticles assessed by multitrophic test battery. Environ. Int. 2013, 51, 97–105. [Google Scholar] [CrossRef]
- Parvez, S.; Venkataraman, C.; Mukherji, S. A review on advantages of implementing luminescence inhibition test (Vibrio fischeri) for acute toxicity prediction of chemicals. Environ. Int. 2006, 32, 265–268. [Google Scholar] [CrossRef]
- Park, K.; Hee, S.Q. Effect of dust on the viability of Vibrio fischeri in the Microtox test. Ecotoxicol. Environ. Saf. 2001, 50, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Official Gazette of Italian Republic. Available online: https://www.gazzettaufficiale.it/eli/id/2016/09/06/16G00184/sg (accessed on 29 June 2021).
- Kaiser, K.L.E. Correlations of Vibrio fischeri bacteria test data with bioassay data for other organisms. Environ. Health Perspect. 1998, 106, 583–591. [Google Scholar] [PubMed]
- Bonnet, M.; Massard, C.; Veisseire, P.; Camares, O.; Awitor, K.O. Environmental toxicity and antimicrobial efficiency of titanium dioxide nanoparticles in suspension. J. Biomater. Nanobiotechnol. 2015, 6, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, M.; Kasemets, K.; Heinlaan, M.; Kurvet, I.; Kahru, A. High throughput kinetic Vibrio fischeri bioluminescence inhibition assay for study of toxic effects of nanoparticles. Toxicol. In Vitro 2008, 22, 1412–1417. [Google Scholar] [CrossRef]
- Velzeboer, I.; Hendriks, A.J.; Ragas, A.M.J.; Van de Meent, D. Aquatic ecotoxicity tests of some nanomaterials. Environ. Toxicol. Chem. 2008, 27, 1942–1947. [Google Scholar] [CrossRef]
- Lopes, I.; Ribeiro, R.; Antunes, F.E.; Rocha-Santos, T.A.P.; Rasteiro, M.G.; Soares, A.M.V.M.; Gonçalves, F.; Pereira, R. Toxicity and genotoxicity of organic and inorganic nanoparticles to the bacteria Vibrio fischeri and Salmonella typhimurium. Ecotoxicology 2012, 21, 637–648. [Google Scholar] [CrossRef]
- Minetto, D.; Libralato, G.; Volpi Ghirardini, A. Ecotoxicity of engineered TiO2 nanoparticles to saltwater organisms: An overview. Environ. Int. 2014, 66, 18–27. [Google Scholar] [CrossRef]
- Baniamerian, H.; Tsapekos, P.; Alvarado-Morales, M.; Shokrollahzadeh, S.; Safavi, M.; Angelidaki, I. Effect of surfactants on photocatalytic toxicity of TiO2-based nanoparticles toward Vibrio fischeri marine bacteria. Inorg. Chem. Commun. 2020, 116, 107936. [Google Scholar] [CrossRef]
- Zeng, C.; Nguyen, C.; Boitano, S.; Field, J.A.; Shadman, F.; Sierra-Alvarez, R. Toxicity of abrasive nanoparticles (SiO2, CeO2, and Al2O3) on Aliivibrio fischeri and human bronchial epithelial cells (16HBE14o). J. Nanopart. Res. 2021, 23, 49. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Mehmood, S.; Tang, H.; Ferreira, L.F.R.; Bharagava, R.N.; Iqbal, H.M.N. Mitigation of environmentally-related hazardous pollutants from water matrices using nanostructured materials—A review. Chemosphere 2020, 253, 126770. [Google Scholar] [CrossRef] [PubMed]
- Coelho, F.P.; Rabelo, T.S.; da Cruz Felix, L.; Bila, D.M.; Ritter, E. Characterization and ecotoxicological evaluation of nanostructured chitosan particles. In Sustainable Environmental Geotechnics; Lecture Notes in Civil Engineering, Reddy, K.R., Agnihotri, A.K., Yukselen-Aksoy, Y., Dubey, B.K., Bansal, A., Eds.; Springer: Cham, Switzerland, 2020; Volume 89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | 1 mg/L | 10 mg/L | 50 mg/L | 100 mg/L | |
|---|---|---|---|---|---|
| MN | 0.95 ± 1.50 | 1.65 ± 2.06 | 1.10 ± 1.17 | 1.50 ± 1.29 | 1.95 ± 1.93 * |
| NPB | 1.05 ± 1.96 | 2.50 ± 2.80 | 2.90 ± 4.09 | 3.00 ± 3.01 | 3.70 ± 4.57 * |
| Apoptosis | 7.60 ± 10.88 | 8.90 ± 9.53 | 5.10 ± 3.70 | 13.44 ± 22.49 | 15.10 ± 14.29 |
| Necrosis | 0.00 ± 0.00 | 0.05 ± 0.22 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Exposure Group | Zn(II) T0 [mg/L] | Zn(II) T48 [mg/L] |
|---|---|---|
| AFW | 0.022 ± 0.003 | 0.02 ± 0.014 |
| Zn(II) | 28.15 ± 6.84 *# | 5.17 ± 3.60 * |
| CNS | 0.183 ± 0.11 | 0.145 ± 0.12 |
| Zn-t CNS | 1.92 ± 0.39 # | 0.45 ± 0.035 |
| Exposure Group | Zn(II) T48 [µg/g] |
|---|---|
| AFW | 121.36 ± 0.54 |
| Zn (II) | 505.50 ± 3.28 ** |
| CNS | 126.05 ± 0.44 |
| Zn-t CNS | 171.66 ± 0.44 * |
| T0 | T48 | |||||||
|---|---|---|---|---|---|---|---|---|
| EC20 (%) | Max Effect (%) | EC20 (%) | Max Effect (%) | |||||
| 5 min | 15 min | 30 min | M ± SD | 5 min | 15 min | 30 min | M ± SD | |
| AFW | >90 | >90 | >90 | −10.30 ± 2.91 | >90 | >90 | >90 | −10.46 ± 12.32 |
| CNS | >90 | >90 | >90 | 0.75 ± 0.40 | >90 | >90 | >80 | n.d. |
| Zn-t CNS | >90 | >90 | >90 | 0.82 ± 10.12 | >90 | >90 | >90 | −16.91 ± 3.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidi, P.; Bernardeschi, M.; Palumbo, M.; Scarcelli, V.; Genovese, M.; Protano, G.; Vitiello, V.; Pontorno, L.; Bonciani, L.; Buttino, I.; et al. Cellular Responses Induced by Zinc in Zebra Mussel Haemocytes. Loss of DNA Integrity as a Cellular Mechanism to Evaluate the Suitability of Nanocellulose-Based Materials in Nanoremediation. Nanomaterials 2021, 11, 2219. https://doi.org/10.3390/nano11092219
Guidi P, Bernardeschi M, Palumbo M, Scarcelli V, Genovese M, Protano G, Vitiello V, Pontorno L, Bonciani L, Buttino I, et al. Cellular Responses Induced by Zinc in Zebra Mussel Haemocytes. Loss of DNA Integrity as a Cellular Mechanism to Evaluate the Suitability of Nanocellulose-Based Materials in Nanoremediation. Nanomaterials. 2021; 11(9):2219. https://doi.org/10.3390/nano11092219
Chicago/Turabian StyleGuidi, Patrizia, Margherita Bernardeschi, Mara Palumbo, Vittoria Scarcelli, Massimo Genovese, Giuseppe Protano, Valentina Vitiello, Lorenzo Pontorno, Lisa Bonciani, Isabella Buttino, and et al. 2021. "Cellular Responses Induced by Zinc in Zebra Mussel Haemocytes. Loss of DNA Integrity as a Cellular Mechanism to Evaluate the Suitability of Nanocellulose-Based Materials in Nanoremediation" Nanomaterials 11, no. 9: 2219. https://doi.org/10.3390/nano11092219
APA StyleGuidi, P., Bernardeschi, M., Palumbo, M., Scarcelli, V., Genovese, M., Protano, G., Vitiello, V., Pontorno, L., Bonciani, L., Buttino, I., Chiaretti, G., Pellegrini, D., Fiorati, A., Riva, L., Punta, C., Corsi, I., & Frenzilli, G. (2021). Cellular Responses Induced by Zinc in Zebra Mussel Haemocytes. Loss of DNA Integrity as a Cellular Mechanism to Evaluate the Suitability of Nanocellulose-Based Materials in Nanoremediation. Nanomaterials, 11(9), 2219. https://doi.org/10.3390/nano11092219