

Different Strategies to Attenuate the Toxic Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells

 , ,
, ,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
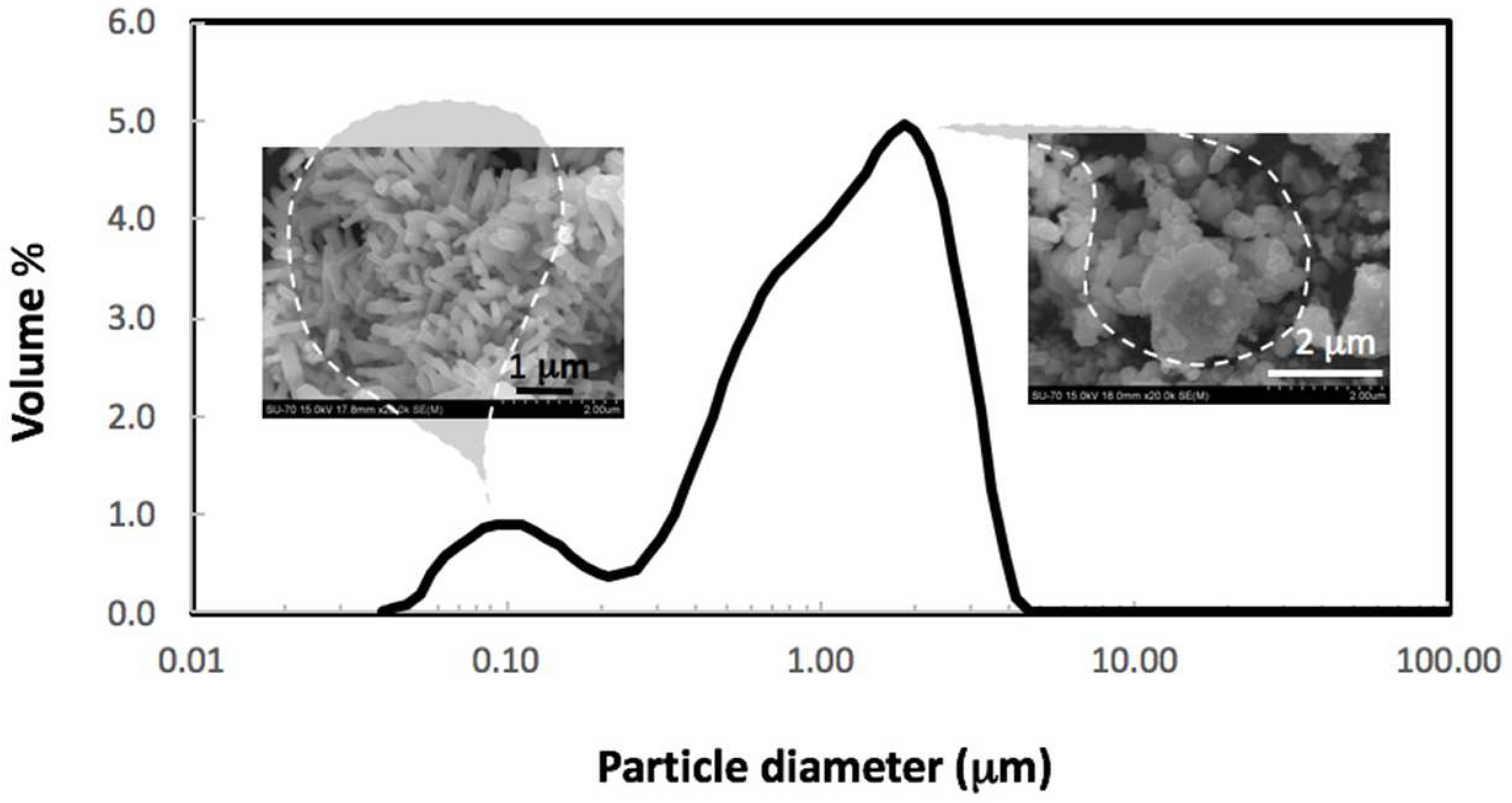
2.1. Characterization of ZnO NPs
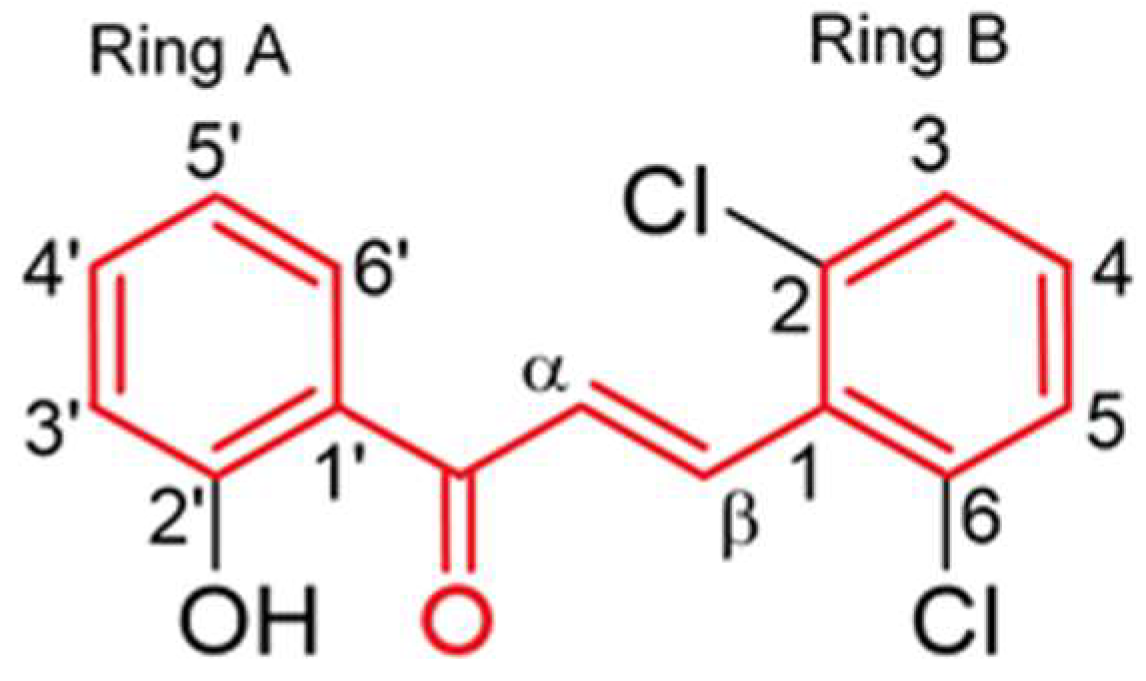
2.2. Synthesis of (E)-3-(2,6-Dichlorophenyl)-1-(2-hydroxyphenyl)prop-2-en-1-one (1)
2.3. Cell Culture
2.4. Experimental Design
2.5. Cell Viability Assay
2.6. SDS-PAGE and Immunoblotting
2.7. Statistical Analysis
3. Results
3.1. Characterization of ZnO NPs
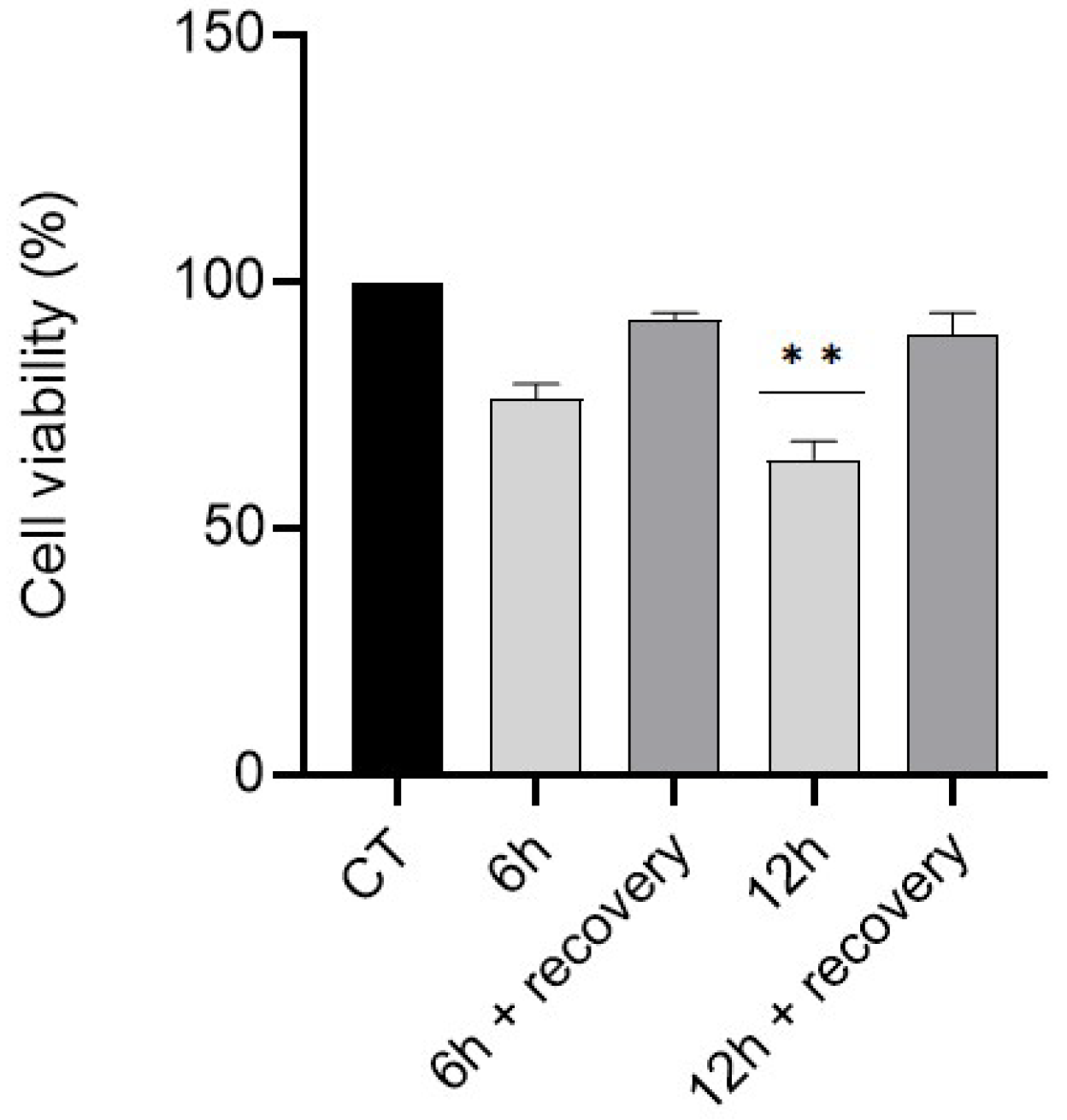
3.2. Effect of a Recovery Period on GC-1 Cells Survival
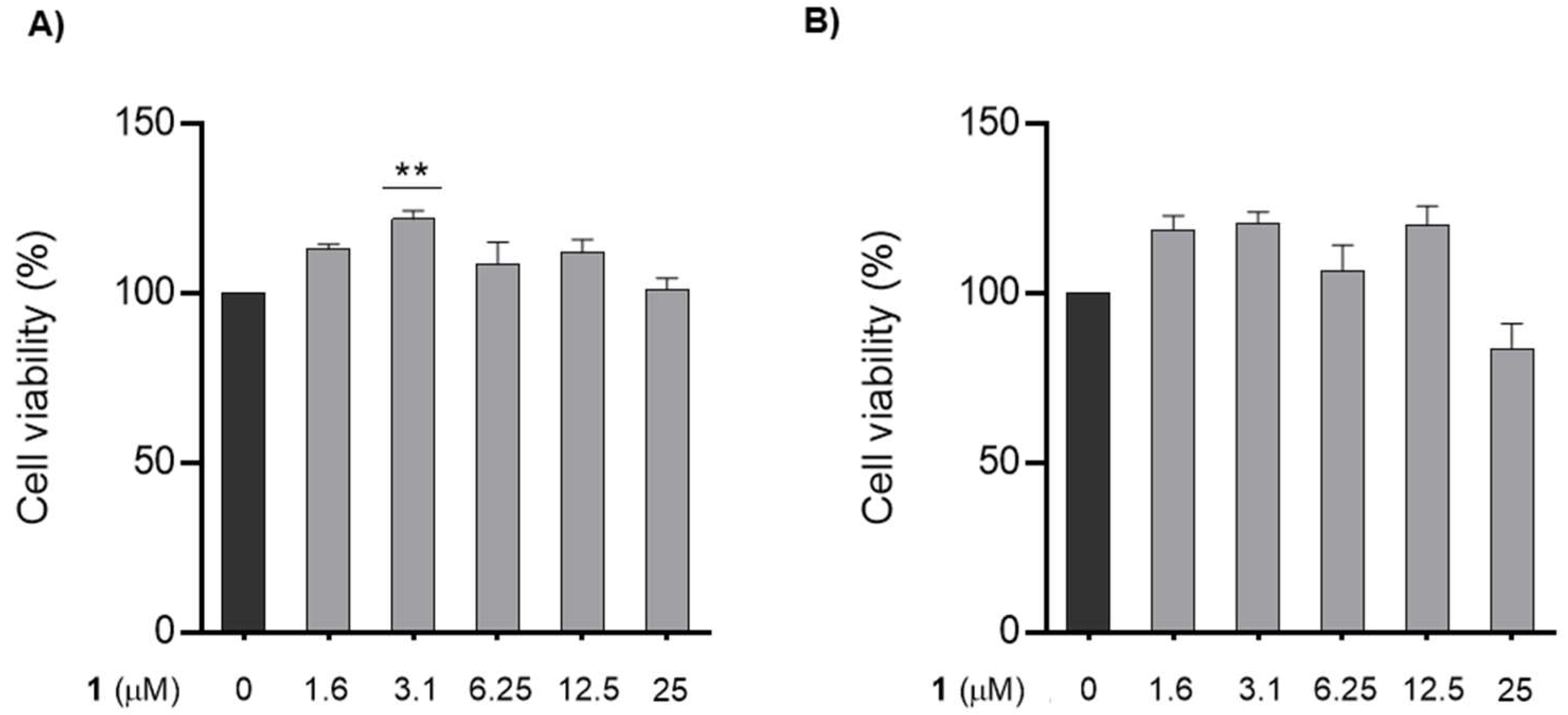
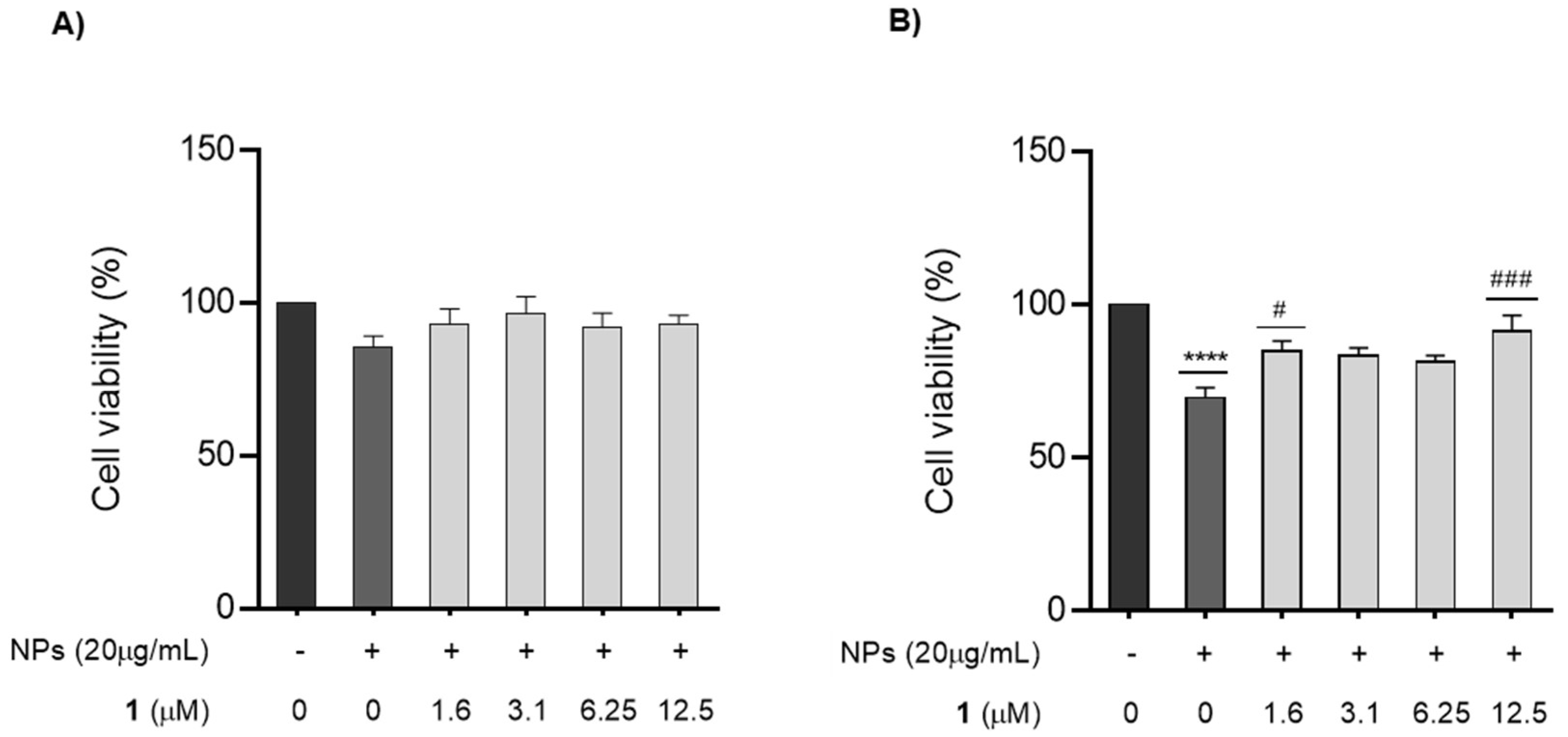
3.3. Effect of Chalcone 1 on GC-1 Cells Survival
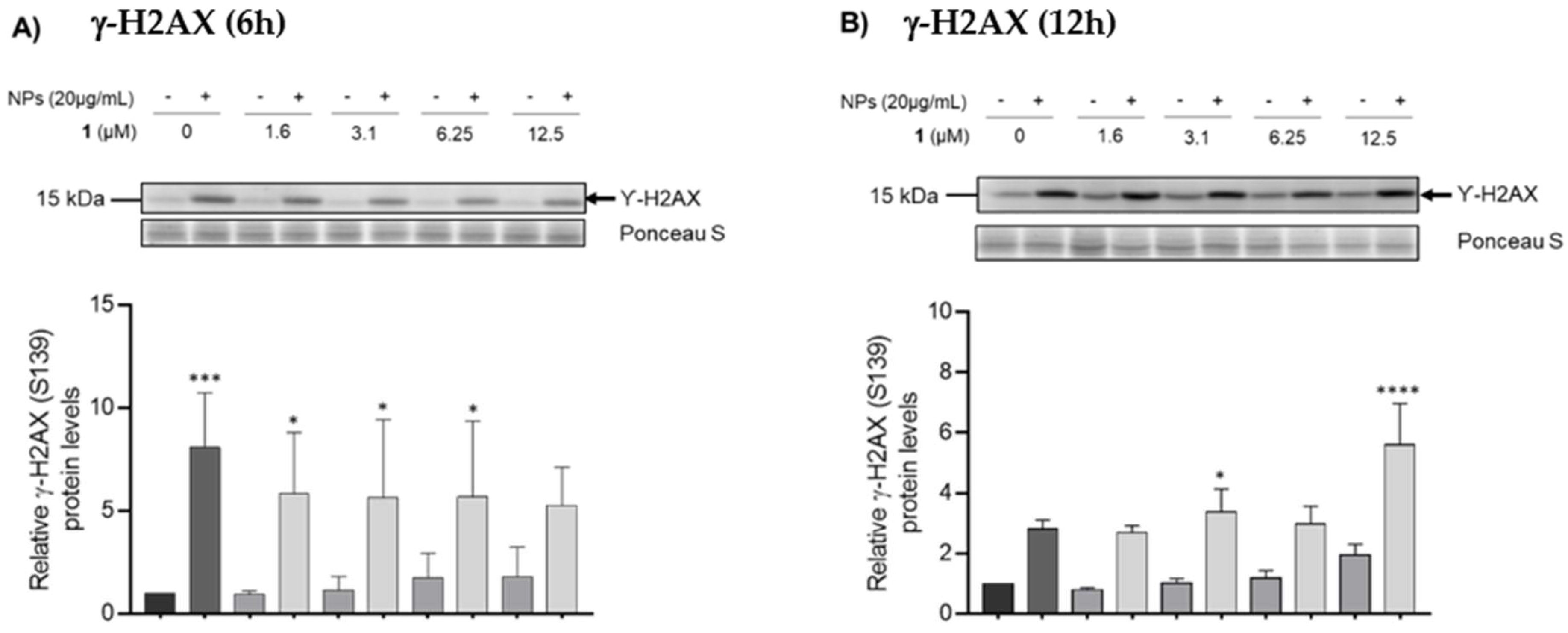
3.4. Counteracting DNA Damage Using Chalcone 1
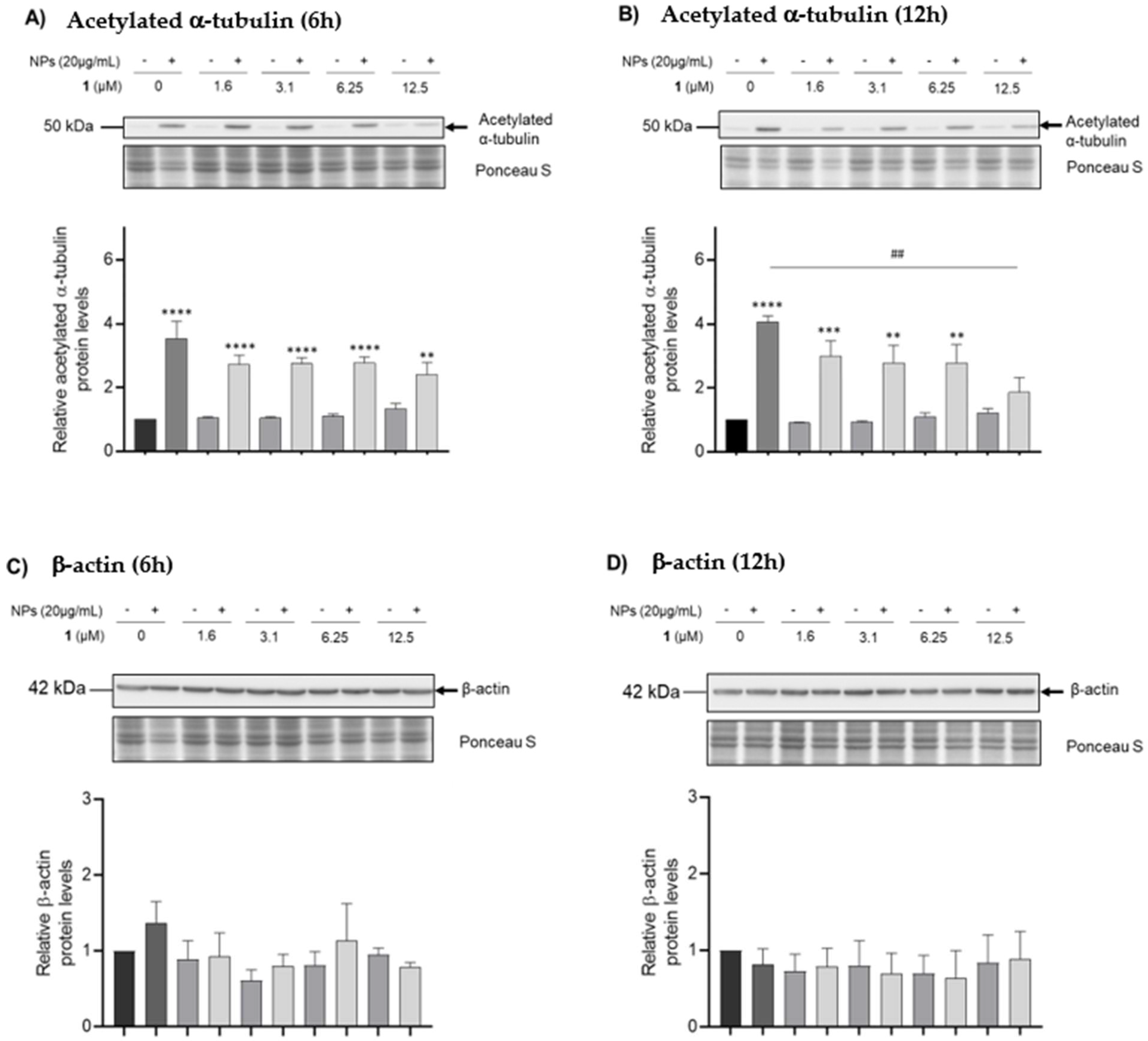
3.5. Analysis of Cytoskeleton Dynamics
4. Discussion
5. Conclusions and Future Trends
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pinho, A.R.; Martins, F.; Costa, M.E.V.; Senos, A.M.R.; Silva, O.A.B.D.C.E.; Pereira, M.D.L.; Rebelo, S. In Vitro Cytotoxicity Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells. Cells 2020, 9, 1081. [Google Scholar] [CrossRef] [PubMed]
- Barkhoradi, A.; Hekmatimoghaddam, S.; Jebali, A.; Khalili, M.; Talebi, A.; Noorani, M. Effect of Zinc Oxide Nanoparticles on Viability of Human Spermatozoa. Iran J. Reprod. Med. 2013, 11, 767–771. [Google Scholar]
- Shen, J.; Yang, D.; Zhou, X.; Wang, Y.; Tang, S.; Yin, H.; Wang, J.; Chen, R.; Chen, J. Role of Autophagy in Zinc Oxide Nanoparticles-Induced Apoptosis of Mouse LEYDIG Cells. Int. J. Mol. Sci. 2019, 20, 4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalpana, V.N.; Rajeswari, V.D. A Review on Green Synthesis, Biomedical Applications, and Toxicity Studies of ZnO NPs. Bioinorg. Chem. Appl. 2018, 2018, 3569758. [Google Scholar] [CrossRef] [PubMed]
- Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Ripolles-Avila, C.; Rodríguez-Jerez, J.J. Antimicrobial Activity and Prevention of Bacterial Biofilm Formation of Silver and Zinc Oxide Nanoparticle-Containing Polyester Surfaces at Various Concentrations for Use. Foods 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staroń, A.; Długosz, O.; Pulit-Prociak, J.; Banach, M. Analysis of the Exposure of Organisms to the Action of Nanomaterials. Materials 2020, 13, 349. [Google Scholar] [CrossRef] [Green Version]
- Özgür, M.E. The Protective Effects of Vitamin C and Trolox on Kinematic and Oxidative Stress Indices in Rainbow Trout Sperm Cells against Flower-like ZnO Nanoparticles. Aquac. Res. 2019, 50, 2838–2845. [Google Scholar] [CrossRef]
- Özgür, M.E.; Ulu, A.; Noma, S.A.A.; Özcan, İ.; Balcıoğlu, S.; Ateş, B.; Köytepe, S. Melatonin Protects Sperm Cells of Capoeta Trutta from Toxicity of Titanium Dioxide Nanoparticles. Environ. Sci. Pollut. Res. 2020, 27, 17843–17853. [Google Scholar] [CrossRef]
- Hussein, M.M.A.; Ali, H.A.; Saadeldin, I.M.; Ahmed, M.M. Querectin Alleviates Zinc Oxide Nanoreprotoxicity in Male Albino Rats. J. Biochem. Mol. Toxicol. 2016, 30, 489–496. [Google Scholar] [CrossRef]
- Martins, T.; Silva, V.L.M.; Silva, A.M.S.; Lima, J.L.F.C.; Fernandes, E.; Ribeiro, D. Chalcones as Scavengers of HOCl and Inhibitors of Oxidative Burst: Structure-Activity Relationship Studies. Med. Chem. 2022, 18, 88–96. [Google Scholar] [CrossRef]
- Sousa, A.; Lucas, M.; Ribeiro, D.; Correia, C.M.; Silva, V.L.M.; Silva, A.M.S.; Fernandes, E.; Freitas, M. Chalcones as Modulators of Neutrophil Oxidative Burst under Physiological and High Glucose Conditions. J. Nat. Prod. 2020, 83, 3131–3140. [Google Scholar] [CrossRef] [PubMed]
- Stepanić, V.; Matijašić, M.; Horvat, T.; Verbanac, D.; Kučerová-Chlupáčová, M.; Saso, L.; Žarković, N. Antioxidant Activities of Alkyl Substituted Pyrazine Derivatives of Chalcones—In Vitro and In Silico Study. Antioxidants 2019, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gacche, R.N.; Dhole, N.A.; Kamble, S.G.; Bandgar, B.P.; Dhole, N.A.; Kamble, S.G.; Bandgar, B.P. In-Vitro Evaluation of Selected Chalcones for Antioxidant Activity. J. Enzyme Inhib. Med. Chem. 2008, 23, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.M.M.; Silva, A.M.S. Chalcones as Versatile Synthons for the Synthesis of 5- and 6-Membered Nitrogen Heterocycles. Curr. Org. Chem. 2014, 18, 2750–2775. [Google Scholar]
- Jasim, H.A.; Nahar, L.; Jasim, M.A.; Moore, S.A.; Ritchie, K.J.; Sarker, S.D. Chalcones: Synthetic Chemistry Follows Where Nature Leads. Biomolecules 2021, 11, 1203. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z.; States, U. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem Rev. 2018, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, L.; Cheng, C.; Li, G.; Qiu, P.; Wu, J. Design, Synthesis and Biological Evaluation of Chalcone Analogues with Novel Dual Antioxidant Mechanisms as Potential Anti-Ischemic Stroke Agents. Acta Pharm. Sin. B 2019, 9, 335–350. [Google Scholar] [CrossRef]
- Rozmer, Z.; Perjési, P. Naturally Occurring Chalcones and Their Biological Activities. Phytochem. Rev. 2016, 15, 87–120. [Google Scholar] [CrossRef]
- Rocha, S.; Sousa, A.; Ribeiro, D.; Correia, C.M.; Silva, V.L.M.; Santos, C.M.M.; Silva, A.M.S.; Araújo, A.N.; Fernandes, E.; Freitas, M. A Study towards Drug Discovery for the Management of Type 2 Diabetes: Mellitus through Inhibition of the Carbohydrate-Hydrolyzing Enzymes α-Amylase and α-Glucosidase by Chalcone Derivatives. Food Funct. 2019, 10, 5510–5520. [Google Scholar] [CrossRef] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phaniendra, A.; Babu, D. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Ind. J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebi, A.R.; Khorsandi, L.; Moridian, M. The Effect of Zinc Oxide Nanoparticles on Mouse Spermatogenesis. J. Assist. Reprod. Genet. 2013, 30, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Donnell, B.O.; Huo, L.; Polli, J.R.; Qiu, L.; Collier, D.N.; Zhang, B.; Pan, X.; Carolina, N.; Services, W.P.; Carolina, N. ZnO Nanoparticles Enhanced Germ Cell Apoptosis in Caenorhabditis Elegans, in Comparison with ZnCl 2. Toxicol. Sci. 2017, 156, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Abbasalipourkabir, R.; Moradi, H.; Zarei, S.; Asadi, S.; Salehzadeh, A.; Ghafourikhosroshahi, A.; Mortazavi, M.; Ziamajidi, N. Toxicity of Zinc Oxide Nanoparticles on Adult Male Wistar Rats. Food Chem. Toxicol. 2015, 84, 154–160. [Google Scholar] [CrossRef]
- Simko, M.; Gazso, A.; Fiedeler, U.; Nentwich, M. Nanoparticles, Free Radicals and Oxidative Stress. NanoTrust Doss. 2011, 12, 1–3. [Google Scholar]
- Lakshmipriya, T.; Gopinath, S. Introduction to Nanoparticles and Analytical Devices. In Nanoparticles in Analytical and Medical Devices; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–29. [Google Scholar]
- Pinho, A.; Rebelo, S.; Pereira, M. The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility. Materials 2020, 13, 849. [Google Scholar] [CrossRef] [Green Version]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative Stress and Male Infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef]
- Agarwal, A.; Leisegang, K.; Majzoub, A.; Henkel, R.; Finelli, R.; Selvam, M.K.P.; Tadros, N.; Parekh, N.; Ko, E.Y.; Cho, C.L.; et al. Utility of Antioxidants in the Treatment of Male Infertility: Clinical Guidelines Based on a Systematic Review and Analysis of Evidence. World J. Men’s Health 2021, 39, 233–290. [Google Scholar] [CrossRef]
- Wu, J.Z.; Cheng, C.C.; Shen, L.L.; Wang, Z.K.; Wu, S.B.; Li, W.L.; Chen, S.H.; Zhou, R.P.; Qiu, P.H. Synthetic Chalcones with Potent Antioxidant Ability on H2O2-Induced Apoptosis in PC12 Cells. Int. J. Mol. Sci. 2014, 15, 18525–18539. [Google Scholar] [CrossRef] [Green Version]
- Janković, T.; Turković, N.; Kotur-Stevuljević, J.; Vujić, Z.; Ivković, B. Differences in antioxidant potential of chalcones in human serum: In vitro study. Chem. Biol. Interact. 2020, 324, 109084. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, H.C.; Yang, X.; Tao, J.J.; Liang, G.; Wu, J.Z.; Wu, W.C.; Wang, Y.; Song, Z.M.; Zhang, X. The Novel Chalcone Analog L2H17 Protects Retinal Ganglion Cells from Oxidative Stress-Induced Apoptosis. Neural. Regen. Res. 2018, 13, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Lahsasni, S.A.; Hamad, F.; Korbi, A.; Aljaber, N.A. Synthesis, Characterization and Evaluation of Antioxidant Activities of Some Novel Chalcones Analogues. Chem. Cent. J. 2014, 8, 32. [Google Scholar] [CrossRef]
- Vh, E.S.; Setyowati, W.A.E. A Green Synthesis of Chalcones As an Antioxidant and Anticancer. IOP Conf. Ser. Mater. Sci. Eng. 2018, 299, 012077. [Google Scholar] [CrossRef]
- Verma, P.; Parte, P. Revisiting the Characteristics of Testicular Germ Cell Lines GC-1(spg) and GC-2(spd)ts. Mol. Biotechnol. 2021, 63, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Breznan, D.; Das, D.; MacKinnon-Roy, C.; Simard, B.; Kumarathasan, P.; Vincent, R. Non-specific interaction of carbon nanotubes with the resazurin assay reagent: Impact on in vitro assessment of nanoparticle cytotoxicity. Toxicol. Vitr. 2015, 29, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.I.A.; Monteiro, C.A.P.; de Oliveira, W.F.; Santos, B.S.; Fontes, A.; Filho, P.E.C. Resazurin-Based Assay to Evaluate Cell Viability After Quantum Dot Interaction. Quantum Dots 2020, 2135, 213–221. [Google Scholar] [CrossRef]
- Pina, S.; Vieira, S.; Rego, P.; Torres, P.; Silva, O.D.C.E.; Silva, E.D.C.E.; Ferreira, J. Biological responses of brushite-forming Zn- and ZnSr- substituted beta-tricalcium phosphate bone cements. Eur. Cells Mater. 2010, 20, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.; Serrano, J.B.; Müller, T.; Silva, O.A.D.C.E.; Rebelo, S. BRI2 Processing and Its Neuritogenic Role Are Modulated by Protein Phosphatase 1 Complexing. J. Cell. Biochem. 2017, 118, 2752–2763. [Google Scholar] [CrossRef]
- Pereira, C.D.; Martins, F.; Santos, M.; Müeller, T.; da Cruz E Silva, O.A.B.; Rebelo, S. Nuclear Accumulation of LAP1:TRF2 Complex. during DNA Damage Response Uncovers a Novel Role for LAP1. Cells 2020, 9, 1804. [Google Scholar] [CrossRef]
- Divito, M.; Miller, W.; Uzarski, J.; Wertheim, J. Essential Design Considerations for the Resazurin Reduction Assay to Noninvasively Quantify Cell Expansion within Perfused Extracellular Matrix Scaffolds. Biomaterials 2017, 129, 163–175. [Google Scholar] [CrossRef]
- Zhao, H.; Huang, X.; Halicka, H.D. Detection of Histone H2AX Phosphorylation on Ser-139 as an Indicator of DNA Damage. Curr. Protoc. Cytom. 2019, 89, e55. [Google Scholar] [CrossRef] [PubMed]
- García-Hevia, L.; Valiente, R.; Martín-Rodríguez, R.; Renero-Lecuna, C.; González, J.; Rodríguez-Fernández, L.; Aguado, F.; Villegas, J.C.; Fanarraga, M.L. Nano-ZnO Leads to Tubulin Macrotube Assembly and Actin Bundling, Triggering Cytoskeletal Catastrophe and Cell Necrosis. Nanoscale 2016, 8, 10963–10973. [Google Scholar] [CrossRef]
- Valdivia, A.; Duran, C.; Martin, A.S. The Role of Nox-Mediated Oxidation in the Regulation of Cytoskeletal Dynamics The Role of Nox-Mediated Oxidation in the Regulation of Cytoskeletal Dynamics. Curr. Pharm. Des. 2015, 21, 6009–6022. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, N.; Yu, Y.; Li, Y.; Li, Y.-B.; Yu, Y.-B.; Zhou, X.-Q.; Sun, Z.-W. Exposure to Silica Nanoparticles Causes Reversible Damage of the Spermatogenic Process in Mice. PLoS ONE 2014, 9, e101572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Zhang, Y.; Zhang, J.; Mu, Q.; Zhang, W.; Elizabeth, R.; Snyder, S.E.; Yan, B.; Chemistry, I. Repeated Carbon Nanotube Administrations in Male Mice Cause Reversible Testis Damage without Affecting Fertility. Nat. Nanotechnol. 2011, 5, 683–689. [Google Scholar] [CrossRef]
- Castellini, C.; Ruggeri, S.; Mattioli, S.; Bernardini, G.; Macchioni, L.; Moretti, E.; Collodel, G. Long-Term Effects of Silver Nanoparticles on Reproductive Activity of Rabbit Buck. Syst. Biol. Reprod. Med. 2014, 60, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quevedo, A.; Lynch, I.; Valsami-Jones, E. Cellular Repair Mechanisms Triggered by Exposure to Silver Nanoparticles to Ionic Silver in Embryonic Zebrafish Cells. Environ. Sci. Nano 2021, 8, 2507–2522. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Al-Kahtani, M.A.; Alsyaad, K.M.; Shalaby, F.M.; Saadeldin, I.M.; Alshammari, F.A.; Mobashar, M.; Suleiman, M.H.; Ali, A.H.H.; Taqi, M.O.; et al. Combined Supplementation of Nano-Zinc Oxide and Thyme Oil Improves the Nutrient Digestibility and Reproductive Fertility in the Male Californian Rabbits. Animals 2020, 10, 2234. [Google Scholar] [CrossRef]
- Hamam, E.; Awadalla, A.; Shokeir, A.; Aboul-Naga, A. Zinc Oxide Nanoparticles Attenuate Prepubertal Exposure to Cisplatin- Induced Testicular Toxicity and Spermatogenesis Impairment in Rats. Toxicology 2022, 28, 153102. [Google Scholar] [CrossRef]
- Mesallam, D.; Deraz, R.; Abdel Aal, S.; Ahmed, S. Toxicity of Subacute Oral Zinc Oxide Nanoparticles on Testes and Prostate of Adult Albino Rats and Role of Recovery. J. Histol. Histopathol. 2019, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Falagan-Lotsch, P.; Grzincic, E.M.; Murphy, C.J. One low-dose exposure of gold nanoparticles induces long-term changes in human cells. Proc. Natl. Acad. Sci. USA 2016, 113, 13318–13323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mironava, T.; Hadjiargyrou, M.; Simon, M.; Jurukovski, V.; Rafailovich, M.H. Gold Nanoparticles Cellular Toxicity and Recovery: Effect of Size, Concentration and Exposure Time. Nanotoxicology 2010, 4, 120–137. [Google Scholar] [CrossRef] [PubMed]
- Oh, N.; Park, J.-H. Endocytosis and Exocytosis of Nanoparticles in Mammalian Cells. Int. J. Nanomed. 2014, 9, 51–63. [Google Scholar]
- Fröhlich, E. Cellular Screening Methods for the Study of Nanoparticle-Induced Lysosomal Damage. In Lysosomes—Associated Diseases and Methods to Study their Function; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Borgese, M.; Rossi, F.; Bonfanti, P.; Colombo, A.; Mantecca, P.; Valdatta, L.; Bernardini, G.; Gornati, R. Recovery Ability of Human Adipose Stem Cells Exposed to Cobalt Nanoparticles: Outcome of Dissolution. Nanomedicine 2020, 15, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Hu, W.; Lu, C.; Cheng, K.; Tang, M. Chemosphere Mechanisms Underlying Nickel Nanoparticle Induced Reproductive Toxicity and Chemo-Protective Effects of Vitamin C in Male Rats. Chemosphere 2019, 218, 259–265. [Google Scholar] [CrossRef]
- Hsu, H.T.; Tseng, Y.T.; Wong, W.J.; Liu, C.M.; Lo, Y.C. Resveratrol Prevents Nanoparticles-Induced Inflammation and Oxidative Stress via Downregulation of PKC-α and NADPH Oxidase in Lung Epithelial A549 Cells. BMC Complement. Altern. Med. 2018, 18, 211. [Google Scholar] [CrossRef]
- Liu, Y.K.; Yang, H.W.; Wang, M.H.; Wang, W.; Liu, F.; Yang, H.L. N-Acetylcysteine Attenuates Cobalt Nanoparticle-Induced Cytotoxic Effects through Inhibition of Cell Death, Reactive Oxygen Species-Related Signaling and Cytokines Expression. Orthop. Surg. 2016, 8, 496–502. [Google Scholar] [CrossRef]
- Wang, J.; Sun, P.; Bao, Y.; Dou, B.; Song, D.; Li, Y. Vitamin E Renders Protection to PC12 Cells against Oxidative Damage and Apoptosis Induced by Single-Walled Carbon Nanotubes. Toxicol. Vitr. 2012, 26, 32–41. [Google Scholar] [CrossRef]
- Han, Z.; Yan, Q.; Ge, W.; Liu, Z.-G.; Gurunathan, S.; Felici, M.; Shen, W.; Zang, X.-F. Cytotoxic Effects of ZnO Nanoparticles on Mouse Testicular Cells. Int. J. Nanomed. 2016, 11, 5187–5203. [Google Scholar] [CrossRef] [Green Version]
- Calarco, A.; Bosetti, M.; Margarucci, S.; Fusaro, L.; Nicolì, E.; Petillo, O.; Cannas, M.; Galderisi, U.; Peluso, G. The Genotoxicity of PEI-Based Nanoparticles Is Reduced by Acetylation of Polyethylenimine Amines in Human Primary Cells. Toxicol. Lett. 2013, 218, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Wigner, P.; Zielinski, K.; Michlewska, S.; Danielska, P.; Marczak, A.; Ricci, E.J.; Santos-Oliveira, R.; Szwed, M. Disturbance of Cellular Homeostasis as a Molecular Risk Evaluation of Human Endothelial Cells Exposed to Nanoparticles. Nat. Sci. Rep. 2021, 11, 3849. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.E.M.; Bryan, M.K.O.; Stanton, P.G.; Donnell, L.O. The Cytoskeleton in Spermatogenesis. Society for Reproduction and Fertility 2019, 157, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yan, M.; Wu, S.; Wu, X.; Bu, T.; Wong, C.K.C.; Ge, R.; Sun, F.; Cheng, C.Y. Actin Binding Proteins, Actin Cytoskeleton and Spermatogenesis—Lesson from Toxicant Models. Reprod. Toxicol. 2020, 96, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Lie, Y.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Cytoskeletal Dynamics and Spermatogenesis. Philos. Trans. R. Soc. 2010, 365, 1581–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekooki, Y.; Haruo, M. Role of Tubulin Acetylation in Cellular Functions and Diseases. Med. Mol. Morphol. 2020, 53, 191–197. [Google Scholar] [CrossRef]
- Ispanixtlahuatl-Meráz, O.; Schins, R.P.F.; Chirino, Y.I. Cell type specific cytoskeleton disruption induced by engineered nanoparticles. Environ. Sci. Nano 2017, 5, 228–245. [Google Scholar] [CrossRef]
- Septiadi, D.; Crippa, F.; Moore, T.L.; Rothen-Rutishauser, B.; Petri-Fink, A. Nanoparticle-Cell Interaction: A Cell Mechanics Perspective. Adv. Mater. 2018, 30, e1704463. [Google Scholar] [CrossRef]
- Janke, C.; Montagnac, G. Causes and Consequences of Microtubule Acetylation. Curr. Biol. 2017, 27, R1287–R1292. [Google Scholar] [CrossRef] [Green Version]
- Královec, K.; Melounková, L.; Slováková, M.; Mannová, N.; Sedlák, M.; Bartáček, J.; Havelek, R. Disruption of Cell Adhesion and Cytoskeletal Networks by Thiol-Functionalized Silica-Coated Iron Oxide Nanoparticles. Int. J. Mol. Sci. 2020, 21, 9350. [Google Scholar] [CrossRef]
- Pernodet, N.; Fang, X.; Sun, Y.; Bakhtina, A.; Ramakrishnan, A.; Sokolov, J.; Ulman, A.; Rafailovich, M. Adverse Effects of Citrate/Gold Nanoparticles on Human Dermal Fibroblasts. Small 2006, 11733, 766–773. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassal, M.; Pereira, C.D.; Martins, F.; Silva, V.L.M.; Silva, A.M.S.; Senos, A.M.R.; Costa, M.E.V.; Pereira, M.d.L.; Rebelo, S. Different Strategies to Attenuate the Toxic Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells. Nanomaterials 2022, 12, 3561. https://doi.org/10.3390/nano12203561
Vassal M, Pereira CD, Martins F, Silva VLM, Silva AMS, Senos AMR, Costa MEV, Pereira MdL, Rebelo S. Different Strategies to Attenuate the Toxic Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells. Nanomaterials. 2022; 12(20):3561. https://doi.org/10.3390/nano12203561
Chicago/Turabian StyleVassal, Mariana, Cátia D. Pereira, Filipa Martins, Vera L. M. Silva, Artur M. S. Silva, Ana M. R. Senos, Maria Elisabete V. Costa, Maria de Lourdes Pereira, and Sandra Rebelo. 2022. "Different Strategies to Attenuate the Toxic Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells" Nanomaterials 12, no. 20: 3561. https://doi.org/10.3390/nano12203561
APA StyleVassal, M., Pereira, C. D., Martins, F., Silva, V. L. M., Silva, A. M. S., Senos, A. M. R., Costa, M. E. V., Pereira, M. d. L., & Rebelo, S. (2022). Different Strategies to Attenuate the Toxic Effects of Zinc Oxide Nanoparticles on Spermatogonia Cells. Nanomaterials, 12(20), 3561. https://doi.org/10.3390/nano12203561