
A New Look at the Effects of Engineered ZnO and TiO2 Nanoparticles: Evidence from Transcriptomics Studies



Abstract
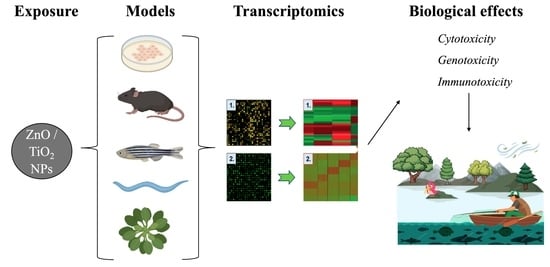
:
1. Introduction
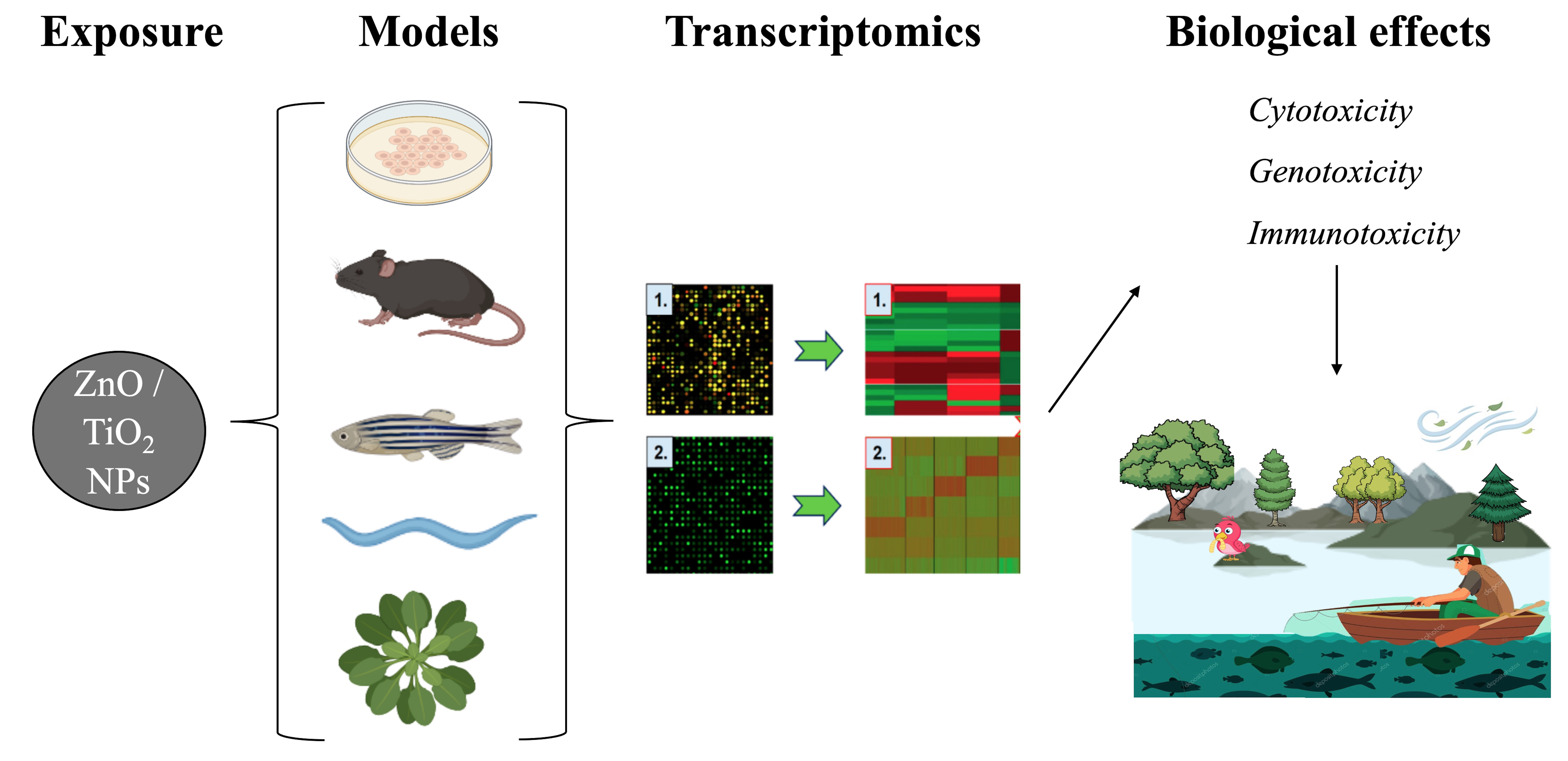
2. Synthetic and Biological Identities of ZnO and TiO2 NPs
2.1. Material Intrinsic Properties
2.1.1. Size
2.1.2. Surface Modifications
2.1.3. Dissolution
2.2. Context-Dependent Properties Relevant for Humans
2.2.1. Port of Entry
2.2.2. Bio-Corona
2.2.3. Cellular Interactions and Trojan Horse Effect
2.2.4. Subcellular Localization
2.3. Context-Dependent Properties Relevant for the Environment
3. Transcriptomic Profiling Relevant to Human Toxicology
3.1. ZnO and TiO2 NPs Exposure In Vitro
3.1.1. Cellular Stress and Cell Death
3.1.2. Developmental and Hereditary Modifications
3.1.3. Changes in Immune Responses
3.2. ZnO and TiO2 NPs Exposure In Vivo
3.2.1. Ingestion
3.2.2. Dermal Contact
3.2.3. Inhalation
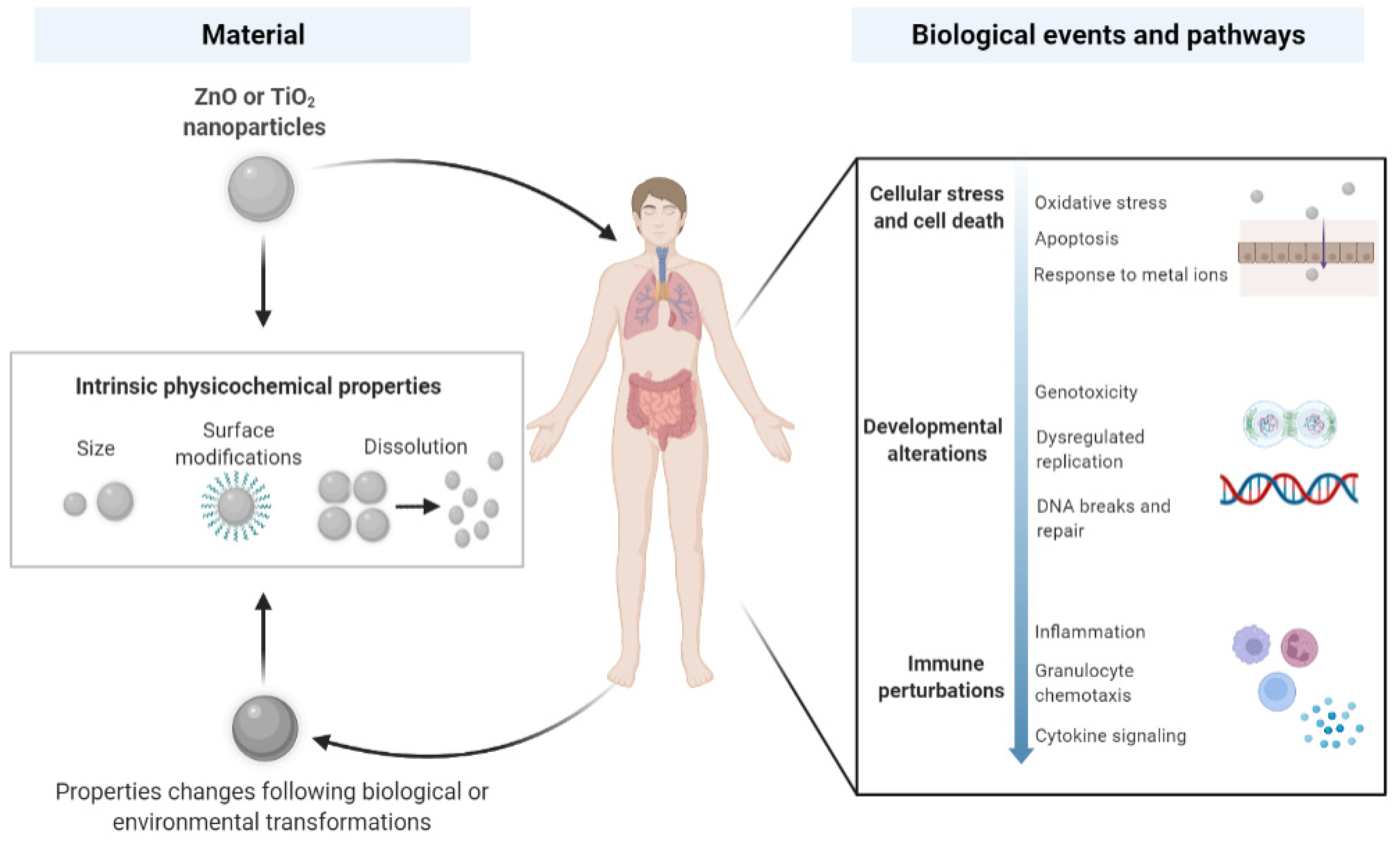
4. Transcriptomic Studies in Environmental Toxicology
4.1. Danio rerio
4.2. Caenorhabditis elegans
4.3. Arabidopsis thaliana
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F., Jr.; Rejeski, D.; Hull, M.S. Nanotechnology in the real world: Redeveloping the nanomaterial consumer products inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Types and Uses of Nanomaterials, Including Safety Aspects; Commission Staff Working Paper SWD 288 Final. Brussels, Belgium, 2012. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=SWD:2012:0288:FIN:EN:PDF (accessed on 1 March 2022).
- World Health Organization. WHO Guidelines on Protecting Workers from Potential Risks of Manufactured Nanomaterials; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Lamas, B.; Martins Breyner, N.; Houdeau, E. Impacts of foodborne inorganic nanoparticles on the gut microbiota-immune axis: Potential consequences for host health. Part. Fibre Toxicol. 2020, 17, 19. [Google Scholar] [CrossRef]
- Canta, M.; Cauda, V. The investigation of the parameters affecting the ZnO nanoparticles cytotoxicity behaviour: A tutorial review. Biomater. Sci. 2020, 8, 6157–6174. [Google Scholar] [CrossRef]
- Schins, R.P.; Lightbody, J.H.; Borm, P.J.; Shi, T.; Donaldson, K.; Stone, V. Inflammatory effects of coarse and fine particulate matter in relation to chemical and biological constituents. Toxicol. Appl. Pharmacol. 2004, 195, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, Z.; Nemmar, A. Health impact of silver nanoparticles: A review of the biodistribution and toxicity following various routes of exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Cummins, E. Hazard characterization of silver nanoparticles for human exposure routes. J. Environ. Sci. Health Part A 2020, 55, 704–725. [Google Scholar] [CrossRef]
- Akter, M.; Sikder, M.T.; Rahman, M.M.; Ullah, A.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Kurasaki, M. A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives. J. Adv. Res. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Maurer, L.; Meyer, J. A systematic review of evidence for silver nanoparticle-induced mitochondrial toxicity. Environ. Sci. Nano 2016, 3, 311–322. [Google Scholar] [CrossRef]
- Marin, S.; Mihail Vlasceanu, G.; Elena Tiplea, R.; Raluca Bucur, I.; Lemnaru, M.; Minodora Marin, M.; Mihai Grumezescu, A. Applications and toxicity of silver nanoparticles: A recent review. Curr. Top. Med. Chem. 2015, 15, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Llop, J.; Estrela-Lopis, I.; Ziolo, R.F.; González, A.; Fleddermann, J.; Dorn, M.; Vallejo, V.G.; Simon-Vazquez, R.; Donath, E.; Mao, Z. Uptake, biological fate, and toxicity of metal oxide nanoparticles. Part. Part. Syst. Charact. 2014, 31, 24–35. [Google Scholar] [CrossRef]
- Aillon, K.L.; Xie, Y.; El-Gendy, N.; Berkland, C.J.; Forrest, M.L. Effects of nanomaterial physicochemical properties on in vivo toxicity. Adv. Drug Deliv. Rev. 2009, 61, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewinski, N.; Colvin, V.; Drezek, R. Cytotoxicity of nanoparticles. Small 2008, 4, 26–49. [Google Scholar] [CrossRef] [PubMed]
- Sturla, S.J.; Boobis, A.R.; FitzGerald, R.E.; Hoeng, J.; Kavlock, R.J.; Schirmer, K.; Whelan, M.; Wilks, M.F.; Peitsch, M.C. Systems toxicology: From basic research to risk assessment. Chem. Res. Toxicol. 2014, 27, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Schena, M.; Shalon, D.; Davis, R.W.; Brown, P.O. Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 1995, 270, 467–470. [Google Scholar] [CrossRef] [Green Version]
- Kukurba, K.R.; Montgomery, S.B. RNA sequencing and analysis. Cold Spring Harb. Protoc. 2015, 2015, pdb.top084970. [Google Scholar]
- Khatri, P.; Sirota, M.; Butte, A.J. Ten years of pathway analysis: Current approaches and outstanding challenges. PLoS Comput. Biol. 2012, 8, e1002375. [Google Scholar] [CrossRef] [PubMed]
- Sengul, A.B.; Asmatulu, E. Toxicity of metal and metal oxide nanoparticles: A review. Environ. Chem. Lett. 2020, 18, 1659–1683. [Google Scholar] [CrossRef]
- Dash, K.K.; Deka, P.; Bangar, S.P.; Chaudhary, V.; Trif, M.; Rusu, A. Applications of Inorganic Nanoparticles in Food Packaging: A Comprehensive Review. Polymers 2022, 14, 521. [Google Scholar] [CrossRef] [PubMed]
- Monopoli, M.P.; Åberg, C.; Salvati, A.; Dawson, K.A. Biomolecular coronas provide the biological identity of nanosized materials. Nat. Nanotechnol. 2012, 7, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular uptake of nanoparticles: Journey inside the cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef] [PubMed]
- British Standards Institution. Publicly Available Specification (PAS) 136: 2007; Terminology for Nanomaterials; British Standards Institution: London, UK, 2007. [Google Scholar]
- Kreuter, J. Encyclopaedia of Pharmaceutical Technology; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 165–190. [Google Scholar]
- Singh, R.; Lillard, J.W., Jr. Nanoparticle-based targeted drug delivery. Exp. Mol. Pathol. 2009, 86, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, H.F.; Wick, P. Nanotoxicology: An interdisciplinary challenge. Angew. Chem. Int. Ed. 2011, 50, 1260–1278. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Asgharian, B.; Price, O.T. Deposition of ultrafine (nano) particles in the human lung. Inhal. Toxicol. 2007, 19, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Geiser, M.; Kreyling, W.G. Deposition and biokinetics of inhaled nanoparticles. Part. Fibre Toxicol. 2010, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praphawatvet, T.; Peters, J.I.; Williams, R.O., III. Inhaled nanoparticles—An updated review. Int. J. Pharm. 2020, 587, 119671. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Navya, P.; Daima, H.K. Rational engineering of physicochemical properties of nanomaterials for biomedical applications with nanotoxicological perspectives. Nano Converg. 2016, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvizo, R.R.; Miranda, O.R.; Thompson, M.A.; Pabelick, C.M.; Bhattacharya, R.; Robertson, J.D.; Rotello, V.M.; Prakash, Y.; Mukherjee, P. Effect of nanoparticle surface charge at the plasma membrane and beyond. Nano Lett. 2010, 10, 2543–2548. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Jiang, C.; Wu, L.; Bai, X.; Zhai, S. Cytotoxicity-related bioeffects induced by nanoparticles: The role of surface chemistry. Front. Bioeng. Biotechnol. 2019, 7, 414. [Google Scholar] [CrossRef] [Green Version]
- Guerrini, L.; Alvarez-Puebla, R.A.; Pazos-Perez, N. Surface modifications of nanoparticles for stability in biological fluids. Materials 2018, 11, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Shen, C.; Feltis, B.N.; Martin, L.L.; Hughes, A.E.; Wright, P.F.; Turney, T.W. Reducing ZnO nanoparticle cytotoxicity by surface modification. Nanoscale 2014, 6, 5791–5798. [Google Scholar] [CrossRef]
- Kittler, S.; Greulich, C.; Diendorf, J.; Koller, M.; Epple, M. Toxicity of silver nanoparticles increases during storage because of slow dissolution under release of silver ions. Chem. Mater. 2010, 22, 4548–4554. [Google Scholar] [CrossRef]
- Jeong, J.; Kim, S.-H.; Lee, S.; Lee, D.-K.; Han, Y.; Jeon, S.; Cho, W.-S. Differential contribution of constituent metal ions to the cytotoxic effects of fast-dissolving metal-oxide nanoparticles. Front. Pharmacol. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Avramescu, M.-L.; Chénier, M.; Palaniyandi, S.; Rasmussen, P.E. Dissolution behavior of metal oxide nanomaterials in cell culture medium versus distilled water. J. Nanopart. Res. 2020, 22, 222. [Google Scholar] [CrossRef]
- Hansen, U.; Thünemann, A.F. Characterization of silver nanoparticles in cell culture medium containing fetal bovine serum. Langmuir 2015, 31, 6842–6852. [Google Scholar] [CrossRef] [PubMed]
- Huynh, K.A.; Chen, K.L. Aggregation kinetics of citrate and polyvinylpyrrolidone coated silver nanoparticles in monovalent and divalent electrolyte solutions. Environ. Sci. Technol. 2011, 45, 5564–5571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zook, J.M.; Long, S.E.; Cleveland, D.; Geronimo, C.L.A.; MacCuspie, R.I. Measuring silver nanoparticle dissolution in complex biological and environmental matrices using UV–visible absorbance. Anal. Bioanal. Chem. 2011, 401, 1993–2002. [Google Scholar] [CrossRef]
- Cho, W.-S.; Kang, B.-C.; Lee, J.K.; Jeong, J.; Che, J.-H.; Seok, S.H. Comparative absorption, distribution, and excretion of titanium dioxide and zinc oxide nanoparticles after repeated oral administration. Part. Fibre Toxicol. 2013, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yao, Y.; Sullivan, N.; Chen, Y. Modeling the primary size effects of citrate-coated silver nanoparticles on their ion release kinetics. Environ. Sci. Technol. 2011, 45, 4422–4428. [Google Scholar] [CrossRef]
- Ma, R.; Levard, C.; Marinakos, S.M.; Cheng, Y.; Liu, J.; Michel, F.M.; Brown, G.E., Jr.; Lowry, G.V. Size-controlled dissolution of organic-coated silver nanoparticles. Environ. Sci. Technol. 2012, 46, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, M.; Kasemets, K.; Kahru, A. Toxicity of ZnO and CuO nanoparticles to ciliated protozoa Tetrahymena thermophila. Toxicology 2010, 269, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, F.; Vogelsberger, W. Dissolution kinetics of synthetic amorphous silica in biological-like media and its theoretical description. J. Phys. Chem. B 2004, 108, 11308–11316. [Google Scholar] [CrossRef]
- Schmidt, J.; Vogelsberger, W. Dissolution kinetics of titanium dioxide nanoparticles: The observation of an unusual kinetic size effect. J. Phys. Chem. B 2006, 110, 3955–3963. [Google Scholar] [CrossRef] [PubMed]
- Franklin, N.M.; Rogers, N.J.; Apte, S.C.; Batley, G.E.; Gadd, G.E.; Casey, P.S. Comparative toxicity of nanoparticulate ZnO, bulk ZnO, and ZnCl2 to a freshwater microalga (Pseudokirchneriella subcapitata): The importance of particle solubility. Environ. Sci. Technol. 2007, 41, 8484–8490. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, L.; Lin, D. Toxicity of ZnO nanoparticles to Escherichia coli: Mechanism and the influence of medium components. Environ. Sci. Technol. 2011, 45, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.; Kramer, E.; Oomen, A.G.; Herrera Rivera, Z.E.; Oegema, G.; Tromp, P.C.; Fokkink, R.; Rietveld, A.; Marvin, H.J.; Weigel, S. Presence of nano-sized silica during in vitro digestion of foods containing silica as a food additive. ACS Nano 2012, 6, 2441–2451. [Google Scholar] [CrossRef]
- Walczak, A.P.; Fokkink, R.; Peters, R.; Tromp, P.; Herrera Rivera, Z.E.; Rietjens, I.M.; Hendriksen, P.J.; Bouwmeester, H. Behaviour of silver nanoparticles and silver ions in an in vitro human gastrointestinal digestion model. Nanotoxicology 2012, 7, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Guo, M.; Cui, X. Effect of food on orally-ingested titanium dioxide and zinc oxide nanoparticle behaviors in simulated digestive tract. Chemosphere 2021, 268, 128843. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, P.; Vauthier, C. Nanotechnology: Intelligent design to treat complex disease. Pharm. Res. 2006, 23, 1417–1450. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, I.-L.; Huang, Y.-J. Effects of serum on cytotoxicity of nano-and micro-sized ZnO particles. J. Nanopart. Res. 2013, 15, 1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, M.G.; Allegri, M.; Chiu, M.; Costa, A.L.; Blosi, M.; Ortelli, S.; Bussolati, O.; Bergamaschi, E. Lipopolysaccharide adsorbed to the bio-Corona of TiO2 nanoparticles powerfully activates selected pro-inflammatory transduction pathways. Front. Immunol. 2017, 8, 866. [Google Scholar] [CrossRef] [Green Version]
- Buchman, J.T.; Hudson-Smith, N.V.; Landy, K.M.; Haynes, C.L. Understanding nanoparticle toxicity mechanisms to inform redesign strategies to reduce environmental impact. Acc. Chem. Res. 2019, 52, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Applerot, G.; Lellouche, J.; Lipovsky, A.; Nitzan, Y.; Lubart, R.; Gedanken, A.; Banin, E. Understanding the antibacterial mechanism of CuO nanoparticles: Revealing the route of induced oxidative stress. Small 2012, 8, 3326–3337. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Fujita, K.; Kato, H.; Endoh, S.; Nishio, K.; Komaba, L.K.; Nakamura, A.; Miyauchi, A.; Kinugasa, S.; Hagihara, Y. Association of the physical and chemical properties and the cytotoxicity of metal oxidenanoparticles: Metal ion release, adsorption ability and specific surface area. Metallomics 2012, 4, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Sabella, S.; Carney, R.P.; Brunetti, V.; Malvindi, M.A.; Al-Juffali, N.; Vecchio, G.; Janes, S.M.; Bakr, O.M.; Cingolani, R.; Stellacci, F. A general mechanism for intracellular toxicity of metal-containing nanoparticles. Nanoscale 2014, 6, 7052–7061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Zhang, J.; Guo, J.; Zhang, J.; Ding, F.; Li, L.; Sun, Z. Role of the dissolved zinc ion and reactive oxygen species in cytotoxicity of ZnO nanoparticles. Toxicol. Lett. 2010, 199, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Rauch, J.; Kolch, W.; Laurent, S.; Mahmoudi, M. Big signals from small particles: Regulation of cell signaling pathways by nanoparticles. Chem. Rev. 2013, 113, 3391–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, L.Y.; Ming, K.; Chan, W.C. Strategies for the intracellular delivery of nanoparticles. Chem. Soc. Rev. 2011, 40, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wu, Y.; Jin, S.; Tian, Y.; Zhang, X.; Zhao, Y.; Yu, L.; Liang, X.-J. Gold nanoparticles induce autophagosome accumulation through size-dependent nanoparticle uptake and lysosome impairment. ACS Nano 2011, 5, 8629–8639. [Google Scholar] [CrossRef]
- National Nanotechnology Initiative. Strategy for Nanotechnology-Related Environmental; Health, and Safety Research, National Science and Technology Council: Washington, DC, USA, 2008. Available online: https://obamawhitehouse.archives.gov/galleries/NSTC/NNI_EHS_Research_Strategy.pdf (accessed on 1 March 2022).
- Abbas, Q.; Yousaf, B.; Ali, M.U.; Munir, M.A.M.; El-Naggar, A.; Rinklebe, J.; Naushad, M. Transformation pathways and fate of engineered nanoparticles (ENPs) in distinct interactive environmental compartments: A review. Environ. Int. 2020, 138, 105646. [Google Scholar] [CrossRef] [PubMed]
- Amde, M.; Liu, J.-f.; Tan, Z.-Q.; Bekana, D. Transformation and bioavailability of metal oxide nanoparticles in aquatic and terrestrial environments. A review. Environ. Pollut. 2017, 230, 250–267. [Google Scholar] [CrossRef] [PubMed]
- Lombi, E.; Donner, E.; Tavakkoli, E.; Turney, T.W.; Naidu, R.; Miller, B.W.; Scheckel, K.G. Fate of zinc oxide nanoparticles during anaerobic digestion of wastewater and post-treatment processing of sewage sludge. Environ. Sci. Technol. 2012, 46, 9089–9096. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Wang, Y.C.; Wang, X.N.; Liu, Y.; Zhang, H.; Zhang, J.W.; Huang, Q.; Chen, S.P.; Hei, T.K.; Wu, L.J. Mutagenicity of ZnO nanoparticles in mammalian cells: Role of physicochemical transformations under the aging process. Nanotoxicology 2015, 9, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Lead, J.R.; Batley, G.E.; Alvarez, P.J.; Croteau, M.N.; Handy, R.D.; McLaughlin, M.J.; Judy, J.D.; Schirmer, K. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects—An updated review. Environ. Toxicol. Chem. 2018, 37, 2029–2063. [Google Scholar] [CrossRef]
- Liu, W.; Zeb, A.; Lian, J.; Wu, J.; Xiong, H.; Tang, J.; Zheng, S. Interactions of metal-based nanoparticles (MBNPs) and metal-oxide nanoparticles (MONPs) with crop plants: A critical review of research progress and prospects. Environ. Rev. 2020, 28, 294–310. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of metal and metal oxide nanoparticles on plant: A critical review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrega, J.; Fawcett, S.R.; Renshaw, J.C.; Lead, J.R. Silver nanoparticle impact on bacterial growth: Effect of pH, concentration, and organic matter. Environ. Sci. Technol. 2009, 43, 7285–7290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qiang, L.; Yuan, Y.; Wu, W.; Sun, B.; Zhu, L. Impacts of titanium dioxide nanoparticles on transformation of silver nanoparticles in aquatic environments. Environ. Sci.: Nano 2018, 5, 1191–1199. [Google Scholar] [CrossRef]
- Ding, Y.; Kuhlbusch, T.A.; Van Tongeren, M.; Jiménez, A.S.; Tuinman, I.; Chen, R.; Alvarez, I.L.; Mikolajczyk, U.; Nickel, C.; Meyer, J. Airborne engineered nanomaterials in the workplace—a review of release and worker exposure during nanomaterial production and handling processes. J. Hazard. Mater. 2017, 322, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Wang, P.; Dang, F.; Zhou, D.-M. The transformation and fate of silver nanoparticles in paddy soil: Effects of soil organic matter and redox conditions. Environ. Sci. Nano 2017, 4, 919–928. [Google Scholar] [CrossRef]
- Sun, W.; Dou, F.; Li, C.; Ma, X.; Ma, L.Q. Impacts of metallic nanoparticles and transformed products on soil health. Crit. Rev. Environ. Sci. Technol. 2021, 51, 973–1002. [Google Scholar] [CrossRef]
- Milani, N.; Hettiarachchi, G.M.; Kirby, J.K.; Beak, D.G.; Stacey, S.P.; McLaughlin, M.J. Fate of zinc oxide nanoparticles coated onto macronutrient fertilizers in an alkaline calcareous soil. PLoS ONE 2015, 10, e0126275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, N.; McLaughlin, M.J.; Stacey, S.P.; Kirby, J.K.; Hettiarachchi, G.M.; Beak, D.G.; Cornelis, G. Dissolution kinetics of macronutrient fertilizers coated with manufactured zinc oxide nanoparticles. J. Agric. Food Chem. 2012, 60, 3991–3998. [Google Scholar] [CrossRef]
- Lapresta-Fernández, A.; Fernández, A.; Blasco, J. Nanoecotoxicity effects of engineered silver and gold nanoparticles in aquatic organisms. TrAC Trends Anal. Chem. 2012, 32, 40–59. [Google Scholar] [CrossRef]
- Khoshnamvand, M.; Hao, Z.; Fadare, O.O.; Hanachi, P.; Chen, Y.; Liu, J. Toxicity of biosynthesized silver nanoparticles to aquatic organisms of different trophic levels. Chemosphere 2020, 258, 127346. [Google Scholar] [CrossRef]
- Matranga, V.; Corsi, I. Toxic effects of engineered nanoparticles in the marine environment: Model organisms and molecular approaches. Mar. Environ. Res. 2012, 76, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.Y.; Li, X.; Sun, M.Z.; Wang, Y.P.; Wu, M.; Zhang, C.Y.; Wang, Y.N.; Liu, B.; Zhang, Y.S.; Zhao, X. An assessment of the impact of SiO2 nanoparticles of different sizes on the rest/wake behavior and the developmental profile of zebrafish larvae. Small 2013, 9, 3161–3168. [Google Scholar] [CrossRef] [PubMed]
- Markus, A.A.; Parsons, J.R.; Roex, E.W.; de Voogt, P.; Laane, R.W. Modelling the Release, Transport and fate of engineered nanoparticles in the aquatic environment—A review. Rev. Environ. Contam. Toxicol. 2016, 243, 53–87. [Google Scholar]
- Gottschalk, F.; Sun, T.; Nowack, B. Environmental concentrations of engineered nanomaterials: Review of modeling and analytical studies. Environ. Pollut. 2013, 181, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Baysal, A.; Saygin, H.; Ustabasi, G.S. Interaction of PM2.5 airborne particulates with ZnO and TiO2 nanoparticles and their effect on bacteria. Environ. Monit. Assess. 2018, 190, 34. [Google Scholar] [CrossRef]
- Jiang, X.; Tong, M.; Kim, H. Influence of natural organic matter on the transport and deposition of zinc oxide nanoparticles in saturated porous media. J. Colloid Interface Sci. 2012, 386, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, H.; Keller, A.A.; Wang, T.; Li, F. The effect of humic acid on the aggregation of titanium dioxide nanoparticles under different pH and ionic strengths. Sci. Total Environ. 2014, 487, 375–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waalewijn-Kool, P.; Ortiz, M.D.; Lofts, S.; van Gestel, C.A. The effect of pH on the toxicity of zinc oxide nanoparticles to Folsomia candida in amended field soil. Environ. Toxicol. Chem. 2013, 32, 2349–2355. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, S.; Luo, L.; Han, W.; Zhang, J.; Yang, K.; Christie, P. Dissolution and microstructural transformation of ZnO nanoparticles under the influence of phosphate. Environ. Sci. Technol. 2012, 46, 7215–7221. [Google Scholar] [CrossRef]
- Kao, Y.-Y.; Chen, Y.-C.; Cheng, T.-J.; Chiung, Y.-M.; Liu, P.-S. Zinc oxide nanoparticles interfere with zinc ion homeostasis to cause cytotoxicity. Toxicol. Sci. 2012, 125, 462–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, S.; Williams, T.D.; Zhang, J.; Zhou, J.A.; Vandebriel, R.J.; De La Fonteyne, L.J.; Gremmer, E.R.; He, S.; Guggenheim, E.J.; Lynch, I. Multi-omics approaches confirm metal ions mediate the main toxicological pathways of metal-bearing nanoparticles in lung epithelial A549 cells. Environ. Sci. Nano 2018, 5, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.K.; Kumar, A.; Pandey, A.K.; Singh, S.S.; Dhawan, A. Titanium dioxide nanoparticles induce oxidative stress-mediated apoptosis in human keratinocyte cells. J. Biomed. Nanotechnol. 2011, 7, 100–101. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.K.; Sharma, V.; Pandey, A.K.; Singh, S.; Sultana, S.; Dhawan, A. ROS-mediated genotoxicity induced by titanium dioxide nanoparticles in human epidermal cells. Toxicol. Vitr. 2011, 25, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Saquib, Q.; Al-Khedhairy, A.A.; Siddiqui, M.A.; Abou-Tarboush, F.M.; Azam, A.; Musarrat, J. Titanium dioxide nanoparticles induced cytotoxicity, oxidative stress and DNA damage in human amnion epithelial (WISH) cells. Toxicol. Vitr. 2012, 26, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Singh, P.; Pandey, A.K.; Dhawan, A. Induction of oxidative stress, DNA damage and apoptosis in mouse liver after sub-acute oral exposure to zinc oxide nanoparticles. Mutat. Res. /Genet. Toxicol. Environ. Mutagenesis 2012, 745, 84–91. [Google Scholar] [CrossRef]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria mediated apoptosis in human liver cells (HepG2). Apoptosis 2012, 17, 852–870. [Google Scholar] [CrossRef] [PubMed]
- Safar, R.; Doumandji, Z.; Saidou, T.; Ferrari, L.; Nahle, S.; Rihn, B.H.; Joubert, O. Cytotoxicity and global transcriptional responses induced by zinc oxide nanoparticles NM 110 in PMA-differentiated THP-1 cells. Toxicol. Lett. 2019, 308, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-J.; Jeong, M.-S.; Choi, D.-M.; Kim, K.-N.; Wie, M.-B. Zinc oxide nanoparticles induce autophagy and apoptosis via oxidative injury and pro-inflammatory cytokines in primary astrocyte cultures. Nanomaterials 2019, 9, 1043. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.K.; Kumar, A.; Gurbani, D.; Pandey, A.K.; Singh, S.; Dhawan, A. TiO2 nanoparticles induce oxidative DNA damage and apoptosis in human liver cells. Nanotoxicology 2013, 7, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Tan, D.; Ze, Y.; Sang, X.; Liu, X.; Gui, S.; Cheng, Z.; Cheng, J.; Hu, R.; Gao, G. Pulmotoxicological effects caused by long-term titanium dioxide nanoparticles exposure in mice. J. Hazard. Mater. 2012, 235, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Han, Y.; Gu, M.; Du, H.; Song, M.; Zhu, X.; Ma, G.; Pan, C.; Wang, W.; Zhao, E. Food Additives: Foodborne Titanium Dioxide Nanoparticles Induce Stronger Adverse Effects in Obese Mice than Non-Obese Mice: Gut Microbiota Dysbiosis, Colonic Inflammation, and Proteome Alterations (Small 36/2020). Small 2020, 16, 2070199. [Google Scholar] [CrossRef]
- Hadrup, N.; Rahmani, F.; Jacobsen, N.R.; Saber, A.T.; Jackson, P.; Bengtson, S.; Williams, A.; Wallin, H.; Halappanavar, S.; Vogel, U. Acute phase response and inflammation following pulmonary exposure to low doses of zinc oxide nanoparticles in mice. Nanotoxicology 2019, 13, 1275–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, S.L.; Tay, C.Y.; Setyawati, M.I.; Leong, D.T. Biomimicry 3D gastrointestinal spheroid platform for the assessment of toxicity and inflammatory effects of zinc oxide nanoparticles. Small 2015, 11, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Doumandji, Z.; Safar, R.; Lovera-Leroux, M.; Nahle, S.; Cassidy, H.; Matallanas, D.; Rihn, B.; Ferrari, L.; Joubert, O. Protein and lipid homeostasis altered in rat macrophages after exposure to metallic oxide nanoparticles. Cell Biol. Toxicol. 2020, 36, 65–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsagaby, S.A.; Vijayakumar, R.; Premanathan, M.; Mickymaray, S.; Alturaiki, W.; Al-Baradie, R.S.; AlGhamdi, S.; Aziz, M.A.; Alhumaydhi, F.A.; Alzahrani, F.A. Transcriptomics-Based Characterization of the Toxicity of ZnO Nanoparticles Against Chronic Myeloid Leukemia Cells. Int. J. Nanomed. 2020, 15, 7901–7921. [Google Scholar] [CrossRef] [PubMed]
- Jevapatarakul, D.; Jiraroj, T.; Payungporn, S.; Chavalit, T.; Khamwut, A.; T-Thienprasert, N.P. Utilization of Cratoxylum formosum crude extract for synthesis of ZnO nanosheets: Characterization, biological activities and effects on gene expression of nonmelanoma skin cancer cell. Biomed. Pharmacother. 2020, 130, 110552. [Google Scholar] [CrossRef]
- Tuomela, S.; Autio, R.; Buerki-Thurnherr, T.; Arslan, O.; Kunzmann, A.; Andersson-Willman, B.; Wick, P.; Mathur, S.; Scheynius, A.; Krug, H.F. Gene expression profiling of immune-competent human cells exposed to engineered zinc oxide or titanium dioxide nanoparticles. PLoS ONE 2013, 8, e68415. [Google Scholar] [CrossRef]
- Hu, H.; Guo, Q.; Fan, X.; Wei, X.; Yang, D.; Zhang, B.; Liu, J.; Wu, Q.; Oh, Y.; Feng, Y. Molecular mechanisms underlying zinc oxide nanoparticle induced insulin resistance in mice. Nanotoxicology 2020, 14, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Choi, S.-J. Particle Size and Biological Fate of ZnO Do Not Cause Acute Toxicity, but Affect Toxicokinetics and Gene Expression Profiles in the Rat Livers after Oral Administration. Int. J. Mol. Sci. 2021, 22, 1698. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Zhao, Y.; Lai, F.-N.; Liu, J.-C.; Sun, Y.-C.; Wang, J.-J.; Cheng, S.-F.; Zhang, X.-F.; Sun, L.-L.; Li, L. Cutaneous applied nano-ZnO reduce the ability of hair follicle stem cells to differentiate. Nanotoxicology 2017, 11, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Osmond-McLeod, M.J.; Oytam, Y.; Rowe, A.; Sobhanmanesh, F.; Greenoak, G.; Kirby, J.; McInnes, E.F.; McCall, M.J. Long-term exposure to commercially available sunscreens containing nanoparticles of TiO2 and ZnO revealed no biological impact in a hairless mouse model. Part. Fibre Toxicol. 2015, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Proquin, H.; Jonkhout, M.C.; Jetten, M.J.; van Loveren, H.; de Kok, T.M.; Briedé, J.J. Transcriptome changes in undifferentiated Caco-2 cells exposed to food-grade titanium dioxide (E171): Contribution of the nano-and micro-sized particles. Sci. Rep. 2019, 9, 18287. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, D.T.; Kumar, A.; Kippner, L.E.; Ho, P.-Y.; Kemp, M.L.; Fan, Y.; Payne, C.K. TiO2 nanoparticles generate superoxide and alter gene expression in human lung cells. RSC Adv. 2019, 9, 25039–25047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuster, E.; Candela, H.; Estévez, J.; Vilanova, E.; Sogorb, M.A. Titanium Dioxide, but Not Zinc Oxide, Nanoparticles Cause SE-Vere Transcriptomic Alterations in t98g Human Glioblastoma Cells. Int. J. Mol. Sci. 2021, 22, 2084. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Jetten, M.J.; Jonkhout, M.C.; Garduño-Balderas, L.G.; Briedé, J.J.; de Kok, T.M.; van Loveren, H.; Chirino, Y.I. Transcriptomics analysis reveals new insights in E171-induced molecular alterations in a mouse model of colon cancer. Sci. Rep. 2018, 8, 9738. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, L.; Guo, Q.; Zong, H.; Yan, Y.; Yin, Y.; Wang, Y.; Oh, Y.; Feng, Y.; Wu, Q. RNA sequencing analysis shows that titanium dioxide nanoparticles induce endoplasmic reticulum stress, which has a central role in mediating plasma glucose in mice. Nanotoxicology 2018, 12, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, H.; Ze, Y.; Zengli, Z.; Hu, Y.; Cheng, Z.; Cheng, J.; Hu, R.; Gao, G.; Wang, L. Gene expression in liver injury caused by long-term exposure to titanium dioxide nanoparticles in mice. Toxicol. Sci. 2012, 128, 171–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, L.; Wang, L.; Sang, X.; Zhao, X.; Hong, J.; Cheng, S.; Yu, X.; Liu, D.; Xu, B.; Hu, R. Nano-sized titanium dioxide-induced splenic toxicity: A biological pathway explored using microarray technology. J. Hazard. Mater. 2014, 278, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Ze, Y.; Li, B.; Zhao, X.; Zhang, T.; Sheng, L.; Hu, R.; Gui, S.; Sang, X.; Sun, Q. Ovarian dysfunction and gene-expressed characteristics of female mice caused by long-term exposure to titanium dioxide nanoparticles. J. Hazard. Mater. 2012, 243, 19–27. [Google Scholar] [CrossRef]
- Park, E.-J.; Yoon, J.; Choi, K.; Yi, J.; Park, K. Induction of chronic inflammation in mice treated with titanium dioxide nanoparticles by intratracheal instillation. Toxicology 2009, 260, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Su, S.F.; Chien, C.T.; Lin, W.H.; Yu, S.L.; Chou, C.C.; Chen, J.J.; Yang, P.C.; Chen, H.W.; Su, S.F. Titanium dioxide nanoparticles induce emphysema-like lung injury in mice. FASEB J. 2006, 20, 2393–2395. [Google Scholar] [CrossRef]
- Halappanavar, S.; Jackson, P.; Williams, A.; Jensen, K.A.; Hougaard, K.S.; Vogel, U.; Yauk, C.L.; Wallin, H. Pulmonary response to surface-coated nanotitanium dioxide particles includes induction of acute phase response genes, inflammatory cascades, and changes in microRNAs: A toxicogenomic study. Environ. Mol. Mutagenesis 2011, 52, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Ze, Y.; Sun, Q.; Zhang, T.; Sang, X.; Cui, Y.; Wang, X.; Gui, S.; Tan, D.; Zhu, M. Molecular mechanisms of nanosized titanium dioxide–Induced pulmonary injury in mice. PLoS ONE 2013, 8, e55563. [Google Scholar] [CrossRef] [Green Version]
- Husain, M.; Saber, A.T.; Guo, C.; Jacobsen, N.R.; Jensen, K.A.; Yauk, C.L.; Williams, A.; Vogel, U.; Wallin, H.; Halappanavar, S. Pulmonary instillation of low doses of titanium dioxide nanoparticles in mice leads to particle retention and gene expression changes in the absence of inflammation. Toxicol. Appl. Pharmacol. 2013, 269, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Rahman, L.; Wu, D.; Johnston, M.; Williams, A.; Halappanavar, S. Toxicogenomics analysis of mouse lung responses following exposure to titanium dioxide nanomaterials reveal their disease potential at high doses. Mutagenesis 2017, 32, 59–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, M.; Wu, D.; Saber, A.T.; Decan, N.; Jacobsen, N.R.; Williams, A.; Yauk, C.L.; Wallin, H.; Vogel, U.; Halappanavar, S. Intratracheally instilled titanium dioxide nanoparticles translocate to heart and liver and activate complement cascade in the heart of C57BL/6 mice. Nanotoxicology 2015, 9, 1013–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, P.; Halappanavar, S.; Hougaard, K.S.; Williams, A.; Madsen, A.M.; Lamson, J.S.; Andersen, O.; Yauk, C.; Wallin, H.; Vogel, U. Maternal inhalation of surface-coated nanosized titanium dioxide (UV-Titan) in C57BL/6 mice: Effects in prenatally exposed offspring on hepatic DNA damage and gene expression. Nanotoxicology 2013, 7, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, P.; Hathaway, Q.; Nichols, C.; Abukabda, A.; Pinti, M.; Shepherd, D.; McBride, C.; Yi, J.; Castranova, V.; Hollander, J. Maternal engineered nanomaterial inhalation during gestation alters the fetal transcriptome. Part. Fibre Toxicol. 2018, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, Q.; Long, J.; Ding, Y.; Zou, X.; Liao, G.; Cao, Y. A comparative study of toxicity of TiO2, ZnO, and Ag nanoparticles to human aortic smooth-muscle cells. Int. J. Nanomed. 2018, 13, 8037. [Google Scholar] [CrossRef] [Green Version]
- Poon, W.-L.; Alenius, H.; Ndika, J.; Fortino, V.; Kolhinen, V.; Meščeriakovas, A.; Wang, M.; Greco, D.; Lähde, A.; Jokiniemi, J. Nano-sized zinc oxide and silver, but not titanium dioxide, induce innate and adaptive immunity and antiviral response in differentiated THP-1 cells. Nanotoxicology 2017, 11, 936–951. [Google Scholar] [CrossRef] [PubMed]
- Monteiro-Riviere, N.A.; Tran, C.L. Nanotoxicology: Characterization, Dosing and Health Effects; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Ikehata, H.; Yamamoto, M. Roles of the KEAP1-NRF2 system in mammalian skin exposed to UV radiation. Toxicol. Appl. Pharmacol. 2018, 360, 69–77. [Google Scholar] [CrossRef]
- López-Armada, M.J.; Riveiro-Naveira, R.R.; Vaamonde-García, C.; Valcárcel-Ares, M.N. Mitochondrial dysfunction and the inflammatory response. Mitochondrion 2013, 13, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.J.; Tait, S.W. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Magdolenova, Z.; Collins, A.; Kumar, A.; Dhawan, A.; Stone, V.; Dusinska, M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 2014, 8, 233–278. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Manshian, B.; Jenkins, G.J.; Griffiths, S.M.; Williams, P.M.; Maffeis, T.G.; Wright, C.J.; Doak, S.H. NanoGenotoxicology: The DNA damaging potential of engineered nanomaterials. Biomaterials 2009, 30, 3891–3914. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.-J.; Chen, C.; Zhao, Y.; Jia, L.; Wang, P.C. Biopharmaceutics and therapeutic potential of engineered nanomaterials. Curr. Drug Metab. 2008, 9, 697–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackenberg, S.; Scherzed, A.; Technau, A.; Kessler, M.; Froelich, K.; Ginzkey, C.; Koehler, C.; Burghartz, M.; Hagen, R.; Kleinsasser, N. Cytotoxic, genotoxic and pro-inflammatory effects of zinc oxide nanoparticles in human nasal mucosa cells in vitro. Toxicol. Vitr. 2011, 25, 657–663. [Google Scholar] [CrossRef] [PubMed]
- EU Commission. Amending Annex VI to Regulation (EC) No 1223/2009 of the european parliament and of the council on cosmetic products. Off. J. Eur. Union 2016, 106, 4. [Google Scholar]
- European Commission. Re-Evaluation. Available online: https://ec.europa.eu/food/safety/food-improvement-agents/additives/re-evaluation_en#ecl-inpage-779 (accessed on 9 November 2021).
- Farrera, C.; Fadeel, B. It takes two to tango: Understanding the interactions between engineered nanomaterials and the immune system. Eur. J. Pharm. Biopharm. 2015, 95, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B. Clear and present danger? Engineered nanoparticles and the immune system. Swiss Med. Wkly. 2012, 142, w13609. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Bondarenko, O.; Kohonen, P.; Andón, F.T.; Brzicová, T.; Gessner, I.; Mathur, S.; Bottini, M.; Calligari, P.; Stella, L. Macrophage sensing of single-walled carbon nanotubes via Toll-like receptors. Sci. Rep. 2018, 8, 1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turabekova, M.; Rasulev, B.; Theodore, M.; Jackman, J.; Leszczynska, D.; Leszczynski, J. Immunotoxicity of nanoparticles: A computational study suggests that CNTs and C 60 fullerenes might be recognized as pathogens by Toll-like receptors. Nanoscale 2014, 6, 3488–3495. [Google Scholar] [CrossRef]
- Kagan, V.E.; Kapralov, A.A.; St. Croix, C.M.; Watkins, S.C.; Kisin, E.R.; Kotchey, G.P.; Balasubramanian, K.; Vlasova, I.I.; Yu, J.; Kim, K. Lung macrophages “digest” carbon nanotubes using a superoxide/peroxynitrite oxidative pathway. ACS Nano 2014, 8, 5610–5621. [Google Scholar] [CrossRef]
- Kagan, V.E.; Konduru, N.V.; Feng, W.; Allen, B.L.; Conroy, J.; Volkov, Y.; Vlasova, I.I.; Belikova, N.A.; Yanamala, N.; Kapralov, A. Carbon nanotubes degraded by neutrophil myeloperoxidase induce less pulmonary inflammation. Nat. Nanotechnol. 2010, 5, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Kotchey, G.P.; Zhao, Y.; Kagan, V.E.; Star, A. Peroxidase-mediated biodegradation of carbon nanotubes in vitro and in vivo. Adv. Drug Deliv. Rev. 2013, 65, 1921–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witasp, E.; Shvedova, A.A.; Kagan, V.E.; Fadeel, B. Single-walled carbon nanotubes impair human macrophage engulfment of apoptotic cell corpses. Inhal. Toxicol. 2009, 21, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Lundborg, M.; Dahlén, S.-E.; Johard, U.; Gerde, P.; Jarstrand, C.; Camner, P.; Låstbom, L. Aggregates of ultrafine particles impair phagocytosis of microorganisms by human alveolar macrophages. Environ. Res. 2006, 100, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Val, S.; Hussain, S.; Boland, S.; Hamel, R.; Baeza-Squiban, A.; Marano, F. Carbon black and titanium dioxide nanoparticles induce pro-inflammatory responses in bronchial epithelial cells: Need for multiparametric evaluation due to adsorption artifacts. Inhal. Toxicol. 2009, 21, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, D.; Liu, W.; Yan, Y.; Zhou, F.; Wu, W.; Yan, Z. Reactive oxygen species trigger NF-κB-mediated NLRP3 inflammasome activation induced by zinc oxide nanoparticles in A549 cells. Toxicol. Ind. Health 2017, 33, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, A.S.; Guarda, G.; Riteau, N.; Drexler, S.K.; Tardivel, A.; Couillin, I.; Tschopp, J. Nanoparticles activate the NLR pyrin domain containing 3 (Nlrp3) inflammasome and cause pulmonary inflammation through release of IL-1α and IL-1β. Proc. Natl. Acad. Sci. USA 2010, 107, 19449–19454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Flavin, K.; Kopf, I.; Radics, G.; Hearnden, C.H.; McManus, G.J.; Moran, B.; Villalta-Cerdas, A.; Echegoyen, L.A.; Giordani, S. Functionalization of carbon nanoparticles modulates inflammatory cell recruitment and NLRP3 inflammasome activation. Small 2013, 9, 4194–4206. [Google Scholar] [CrossRef] [PubMed]
- Peeters, P.M.; Perkins, T.N.; Wouters, E.F.; Mossman, B.T.; Reynaert, N.L. Silica induces NLRP3 inflammasome activation in human lung epithelial cells. Part. Fibre Toxicol. 2013, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bonner, J.C.; Brown, J.M. Interaction of Nanomaterials with the Immune System; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Shvedova, A.; Kisin, E.; Murray, A.; Kommineni, C.; Castranova, V.; Fadeel, B.; Kagan, V. Increased accumulation of neutrophils and decreased fibrosis in the lung of NADPH oxidase-deficient C57BL/6 mice exposed to carbon nanotubes. Toxicol. Appl. Pharmacol. 2008, 231, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Schanen, B.C.; Karakoti, A.S.; Seal, S.; Drake , D.R., III; Warren, W.L.; Self, W.T. Exposure to titanium dioxide nanomaterials provokes inflammation of an in vitro human immune construct. ACS Nano 2009, 3, 2523–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraschi, D.; Costantino, L.; Italiani, P. Interaction of nanoparticles with immunocompetent cells: Nanosafety considerations. Nanomedicine 2012, 7, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Zolnik, B.S.; González-Fernández, Á.; Sadrieh, N.; Dobrovolskaia, M.A. Minireview: Nanoparticles and the immune system. Endocrinology 2010, 151, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Laverny, G.; Casset, A.; Purohit, A.; Schaeffer, E.; Spiegelhalter, C.; de Blay, F.; Pons, F. Immunomodulatory properties of multi-walled carbon nanotubes in peripheral blood mononuclear cells from healthy subjects and allergic patients. Toxicol. Lett. 2013, 217, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Tkach, A.V.; Shurin, G.V.; Shurin, M.R.; Kisin, E.R.; Murray, A.R.; Young, S.-H.; Star, A.; Fadeel, B.; Kagan, V.E.; Shvedova, A.A. Direct effects of carbon nanotubes on dendritic cells induce immune suppression upon pulmonary exposure. ACS Nano 2011, 5, 5755–5762. [Google Scholar] [CrossRef]
- Wijnhoven, S.W.; Peijnenburg, W.J.; Herberts, C.A.; Hagens, W.I.; Oomen, A.G.; Heugens, E.H.; Roszek, B.; Bisschops, J.; Gosens, I.; Van De Meent, D. Nano-silver–a review of available data and knowledge gaps in human and environmental risk assessment. Nanotoxicology 2009, 3, 109–138. [Google Scholar] [CrossRef]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; Von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, J.P.M.; Chen, A.L.; Foster, A.; Drezek, R. In vivo biodistribution of nanoparticles. Nanomedicine 2011, 6, 815–835. [Google Scholar] [CrossRef] [PubMed]
- Ghamsari, M.S.; Alamdari, S.; Han, W.; Park, H.-H. Impact of nanostructured thin ZnO film in ultraviolet protection. Int. J. Nanomed. 2017, 12, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M. Antibacterial, deodorizing, and UV absorbing materials obtained with zinc oxide (ZnO) coated fabrics. J. Coat. Fabr. 1993, 23, 150–164. [Google Scholar] [CrossRef]
- Schneider, S.L.; Lim, H.W. A review of inorganic UV filters zinc oxide and titanium dioxide. Photodermatol. Photoimmunol. Photomed. 2019, 35, 442–446. [Google Scholar] [CrossRef] [Green Version]
- Landsiedel, R.; Ma-Hock, L.; Kroll, A.; Hahn, D.; Schnekenburger, J.; Wiench, K.; Wohlleben, W. Testing metal-oxide nanomaterials for human safety. Adv. Mater. 2010, 22, 2601–2627. [Google Scholar] [CrossRef] [PubMed]
- Maynard, A.D.; Baron, P.A.; Foley, M.; Shvedova, A.A.; Kisin, E.R.; Castranova, V. Exposure to carbon nanotube material: Aerosol release during the handling of unrefined single-walled carbon nanotube material. J. Toxicol. Environ. Health Part A 2004, 67, 87–107. [Google Scholar] [CrossRef]
- Adamcakova-Dodd, A.; Stebounova, L.V.; Kim, J.S.; Vorrink, S.U.; Ault, A.P.; T O’Shaughnessy, P.; Grassian, V.H.; Thorne, P.S. Toxicity assessment of zinc oxide nanoparticles using sub-acute and sub-chronic murine inhalation models. Part. Fibre Toxicol. 2014, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, Y.; Izumi, H.; Yoshiura, Y.; Tomonaga, T.; Oyabu, T.; Myojo, T.; Kawai, K.; Yatera, K.; Shimada, M.; Kubo, M. Evaluation of pulmonary toxicity of zinc oxide nanoparticles following inhalation and intratracheal instillation. Int. J. Mol. Sci. 2016, 17, 1241. [Google Scholar] [CrossRef]
- Chen, J.-K.; Ho, C.-C.; Chang, H.; Lin, J.-F.; Yang, C.S.; Tsai, M.-H.; Tsai, H.-T.; Lin, P. Particulate nature of inhaled zinc oxide nanoparticles determines systemic effects and mechanisms of pulmonary inflammation in mice. Nanotoxicology 2015, 9, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-L.; Lee, Y.-H.; Chen, H.-I.; Liao, H.-S.; Chiang, B.-L.; Cheng, T.-J. Zinc oxide nanoparticles induce eosinophilic airway inflammation in mice. J. Hazard. Mater. 2015, 297, 304–312. [Google Scholar] [CrossRef]
- Xia, T.; Zhao, Y.; Sager, T.; George, S.; Pokhrel, S.; Li, N.; Schoenfeld, D.; Meng, H.; Lin, S.; Wang, X. Decreased dissolution of ZnO by iron doping yields nanoparticles with reduced toxicity in the rodent lung and zebrafish embryos. ACS Nano 2011, 5, 1223–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Tang, Q.; Zhang, J.; Wang, H.; Bai, L.; Meng, P.; Qin, X.; Xu, G.; Bose, D.D.; Wang, B. Autophagy-dependent release of zinc ions is critical for acute lung injury triggered by zinc oxide nanoparticles. Nanotoxicology 2018, 12, 1068–1091. [Google Scholar] [CrossRef]
- Cho, W.-S.; Duffin, R.; Howie, S.E.; Scotton, C.J.; Wallace, W.A.; MacNee, W.; Bradley, M.; Megson, I.L.; Donaldson, K. Progressive severe lung injury by zinc oxide nanoparticles; the role of Zn2+ dissolution inside lysosomes. Part. Fibre Toxicol. 2011, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Underhill, D.M.; Goodridge, H.S. Information processing during phagocytosis. Nat. Rev. Immunol. 2012, 12, 492–502. [Google Scholar] [CrossRef] [Green Version]
- Seong, S.-Y.; Matzinger, P. Hydrophobicity: An ancient damage-associated molecular pattern that initiates innate immune responses. Nat. Rev. Immunol. 2004, 4, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Kinaret, P.A.; Ndika, J.; Ilves, M.; Wolff, H.; Vales, G.; Norppa, H.; Savolainen, K.; Skoog, T.; Kere, J.; Moya, S. Toxicogenomic Profiling of 28 Nanomaterials in Mouse Airways. Adv. Sci. 2021, 8, 2004588. [Google Scholar] [CrossRef]
- Song, B.; Liu, J.; Feng, X.; Wei, L.; Shao, L. A review on potential neurotoxicity of titanium dioxide nanoparticles. Nanoscale Res. Lett. 2015, 10, 342. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.; Weaver, C. Janeway’s Immunobiology; Garland science: New York, NY, USA, 2016. [Google Scholar]
- Hou, J.; Liu, H.; Wang, L.; Duan, L.; Li, S.; Wang, X. Molecular toxicity of metal oxide nanoparticles in Danio rerio. Environ. Sci. Technol. 2018, 52, 7996–8004. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Kim, R.-O.; Yoon, S.; Kim, W.-K. Developmental toxicity of zinc oxide nanoparticles to zebrafish (Danio rerio): A transcriptomic analysis. PLoS ONE 2016, 11, e0160763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, R.-O.; Choi, J.S.; Kim, B.-C.; Kim, W.-K. Comparative analysis of transcriptional profile changes in larval zebrafish exposed to zinc oxide nanoparticles and zinc sulfate. Bull. Environ. Contam. Toxicol. 2017, 98, 183–189. [Google Scholar] [CrossRef]
- Starnes, D.; Unrine, J.; Chen, C.; Lichtenberg, S.; Starnes, C.; Svendsen, C.; Kille, P.; Morgan, J.; Baddar, Z.E.; Spear, A. Toxicogenomic responses of Caenorhabditis elegans to pristine and transformed zinc oxide nanoparticles. Environ. Pollut. 2019, 247, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Landa, P.; Vankova, R.; Andrlova, J.; Hodek, J.; Marsik, P.; Storchova, H.; White, J.C.; Vanek, T. Nanoparticle-specific changes in Arabidopsis thaliana gene expression after exposure to ZnO, TiO2, and fullerene soot. J. Hazard. Mater. 2012, 241, 55–62. [Google Scholar] [CrossRef]
- Landa, P.; Prerostova, S.; Petrova, S.; Knirsch, V.; Vankova, R.; Vanek, T. The transcriptomic response of Arabidopsis thaliana to zinc oxide: A comparison of the impact of nanoparticle, bulk, and ionic zinc. Environ. Sci. Technol. 2015, 49, 14537–14545. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, B.; Ji, T.; Palić, D. Gene expression of zebrafish embryos exposed to titanium dioxide nanoparticles and hydroxylated fullerenes. Ecotoxicol. Environ. Saf. 2011, 74, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, X.; Zhang, X.; Zhao, Z.; Liu, H.; George, R.; Wilson-Rawls, J.; Chang, Y.; Chen, Y. Disruption of zebrafish (Danio rerio) reproduction upon chronic exposure to TiO2 nanoparticles. Chemosphere 2011, 83, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-C.; Wu, G.-H.; Hua, T.-E.; Wagner, O.I.; Yen, T.-J. Uptake of TiO2 nanoparticles into C. elegans neurons negatively affects axonal growth and worm locomotion behavior. ACS Appl. Mater. Interfaces 2018, 10, 8485–8495. [Google Scholar] [CrossRef]
- Rocheleau, S.; Arbour, M.; Elias, M.; Sunahara, G.I.; Masson, L. Toxicogenomic effects of nano-and bulk-TiO2 particles in the soil nematode Caenorhabditis elegans. Nanotoxicology 2015, 9, 502–512. [Google Scholar] [CrossRef]
- Tumburu, L.; Andersen, C.P.; Rygiewicz, P.T.; Reichman, J.R. Phenotypic and genomic responses to titanium dioxide and cerium oxide nanoparticles in Arabidopsis germinants. Environ. Toxicol. Chem. 2015, 34, 70–83. [Google Scholar] [CrossRef]
- Felix, L.C.; Ortega, V.A.; Ede, J.D.; Goss, G.G. Physicochemical characteristics of polymer-coated metal-oxide nanoparticles and their toxicological effects on zebrafish (Danio rerio) development. Environ. Sci. Technol. 2013, 47, 6589–6596. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, X.; Chen, Y.; Chang, Y. Application of embryonic and adult zebrafish for nanotoxicity assessment. In Nanotoxicity; Springer: Berlin/Heidelberg, Germany, 2012; pp. 317–329. [Google Scholar]
- Parng, C. In vivo zebrafish assays for toxicity testing. Curr. Opin. Drug Discov. Dev. 2005, 8, 100–106. [Google Scholar]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef]
- Rubinstein, A.L. Zebrafish assays for drug toxicity screening. Expert Opin. Drug Metab. Toxicol. 2006, 2, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, S.; Wu, Y.; You, H.; Lv, L. Acute ZnO nanoparticles exposure induces developmental toxicity, oxidative stress and DNA damage in embryo-larval zebrafish. Aquat. Toxicol. 2013, 136, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Qiu, Z.; Chen, Z.; Du Laing, G.; Liu, A.; Yan, C. Impact of TiO2 and ZnO nanoparticles at predicted environmentally relevant concentrations on ammonia-oxidizing bacteria cultures under ammonia oxidation. Environ. Sci. Pollut. Res. 2015, 22, 2891–2899. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.C.; Nowack, B. Exposure modeling of engineered nanoparticles in the environment. Environ. Sci. Technol. 2008, 42, 4447–4453. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, L.; Duan, Z.; Qi, R.; Li, Y.; Lang, Y. Comparative toxicity of several metal oxide nanoparticle aqueous suspensions to Zebrafish (Danio rerio) early developmental stage. J. Environ. Sci. Health Part A 2008, 43, 278–284. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, J.; Zhang, X.; Chang, Y.; Chen, Y. The impact of ZnO nanoparticle aggregates on the embryonic development of zebrafish (Danio rerio). Nanotechnology 2009, 20, 195103. [Google Scholar] [CrossRef]
- Hao, L.; Chen, L.; Hao, J.; Zhong, N. Bioaccumulation and sub-acute toxicity of zinc oxide nanoparticles in juvenile carp (Cyprinus carpio): A comparative study with its bulk counterparts. Ecotoxicol. Environ. Saf. 2013, 91, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Mehinto, A.C.; Prucha, M.S.; Colli-Dula, R.C.; Kroll, K.J.; Lavelle, C.M.; Barber, D.S.; Vulpe, C.D.; Denslow, N.D. Gene networks and toxicity pathways induced by acute cadmium exposure in adult largemouth bass (Micropterus salmoides). Aquat. Toxicol. 2014, 152, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, D.; Jimmy, C.Y.; Chan, K.M. Effects of Cu2O nanoparticle and CuCl2 on zebrafish larvae and a liver cell-line. Aquat. Toxicol. 2011, 105, 344–354. [Google Scholar] [CrossRef]
- Zhou, Z.; Son, J.; Harper, B.; Zhou, Z.; Harper, S. Influence of surface chemical properties on the toxicity of engineered zinc oxide nanoparticles to embryonic zebrafish. Beilstein J. Nanotechnol. 2015, 6, 1568–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.; Fang, T.; Yu, L.; Sima, X.; Zhu, W. Effects of nano-scale TiO2, ZnO and their bulk counterparts on zebrafish: Acute toxicity, oxidative stress and oxidative damage. Sci. Total Environ. 2011, 409, 1444–1452. [Google Scholar] [CrossRef]
- Wu, Q.; Qu, Y.; Li, X.; Wang, D. Chromium exhibits adverse effects at environmental relevant concentrations in chronic toxicity assay system of nematode Caenorhabditis elegans. Chemosphere 2012, 87, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; He, X.; Zhang, Z.; Zhang, P.; Li, Y.; Ma, Y.; Kuang, Y.; Zhao, Y.; Chai, Z. Nano-CeO2 exhibits adverse effects at environmental relevant concentrations. Environ. Sci. Technol. 2011, 45, 3725–3730. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Pradhan, A.; Khan, F.A.; Lindström, P.; Ragnvaldsson, D.; Ivarsson, P.; Olsson, P.-E.; Jass, J. Comparative analysis of stress induced gene expression in Caenorhabditis elegans following exposure to environmental and lab reconstituted complex metal mixture. PLoS ONE 2015, 10, e0132896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejeda-Benitez, L.; Flegal, R.; Odigie, K.; Olivero-Verbel, J. Pollution by metals and toxicity assessment using Caenorhabditis elegans in sediments from the Magdalena River, Colombia. Environ. Pollut. 2016, 212, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Rui, Q.; Cai, T.; Wu, Q.; Li, Y.; Wang, D. Close association of intestinal autofluorescence with the formation of severe oxidative damage in intestine of nematodes chronically exposed to Al2O3-nanoparticle. Environ. Toxicol. Pharmacol. 2011, 32, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yang, Y.; Wang, D. Induction of reproductive deficits in nematode Caenorhabditis elegans exposed to metals at different developmental stages. Reprod. Toxicol. 2009, 28, 90–95. [Google Scholar] [CrossRef]
- Wang, H.; Wick, R.L.; Xing, B. Toxicity of nanoparticulate and bulk ZnO, Al2O3 and TiO2 to the nematode Caenorhabditis elegans. Environ. Pollut. 2009, 157, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Bertsch, P.M.; Glenn, T.C.; Kabengi, N.J.; Williams, P.L. Toxicity of manufactured zinc oxide nanoparticles in the nematode Caenorhabditis elegans. Environ. Toxicol. Chem. Int. J. 2009, 28, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-Y.; Park, Y.-K.; Park, K.; Choi, J. Ecotoxicological investigation of CeO2 and TiO2 nanoparticles on the soil nematode Caenorhabditis elegans using gene expression, growth, fertility, and survival as endpoints. Environ. Toxicol. Pharmacol. 2010, 29, 167–172. [Google Scholar] [CrossRef]
- Khare, P.; Sonane, M.; Pandey, R.; Ali, S.; Gupta, K.C.; Satish, A. Adverse effects of TiO2 and ZnO nanoparticles in soil nematode, Caenorhabditis elegans. J. Biomed. Nanotechnol. 2011, 7, 116–117. [Google Scholar] [CrossRef]
- Ma, H.; Kabengi, N.; Bertsch, P.; Unrine, J.; Glenn, T.; Williams, P. Comparative phototoxicity of nanoparticulate and bulk ZnO to a free-living nematode Caenorhabditis elegans: The importance of illumination mode and primary particle size. Environ. Pollut. 2011, 159, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-W.; Li, S.-W.; Liao, V.H.-C. Chronic ZnO-NPs exposure at environmentally relevant concentrations results in metabolic and locomotive toxicities in Caenorhabditis elegans. Environ. Pollut. 2017, 220, 1456–1464. [Google Scholar] [CrossRef]
- Huang, C.-W.; Li, S.-W.; Liao, V.H.-C. Long-term sediment exposure to ZnO nanoparticles induces oxidative stress in Caenorhabditis elegans. Environ. Sci. Nano 2019, 6, 2602–2614. [Google Scholar] [CrossRef]
- Wu, Q.; Nouara, A.; Li, Y.; Zhang, M.; Wang, W.; Tang, M.; Ye, B.; Ding, J.; Wang, D. Comparison of toxicities from three metal oxide nanoparticles at environmental relevant concentrations in nematode Caenorhabditis elegans. Chemosphere 2013, 90, 1123–1131. [Google Scholar] [CrossRef]
- McLaggan, D.; Amezaga, M.R.; Petra, E.; Frost, A.; Duff, E.I.; Rhind, S.M.; Fowler, P.A.; Glover, L.A.; Lagido, C. Impact of sublethal levels of environmental pollutants found in sewage sludge on a novel Caenorhabditis elegans model biosensor. PLoS ONE 2012, 7, e46503. [Google Scholar] [CrossRef] [Green Version]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nanoparticle Res. 2012, 14, 1125. [Google Scholar] [CrossRef]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Farzaneh, M.; Ghassempour, A. Effect of silver nanoparticles on Oryza sativa L. and its rhizosphere bacteria. Ecotoxicol. Environ. Saf. 2013, 88, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Peng, B.; Hernandez-Viezcas, J.A.; Rico, C.; Sun, Y.; Peralta-Videa, J.R.; Tang, X.; Niu, G.; Jin, L.; Varela-Ramirez, A. Stress response and tolerance of Zea mays to CeO2 nanoparticles: Cross talk among H2O2, heat shock protein, and lipid peroxidation. ACS Nano 2012, 6, 9615–9622. [Google Scholar] [CrossRef] [Green Version]
- Sunahara, G.I.; Renoux, A.Y.; Thellen, C.; Gaudet, C.L.; Pilon, A. Environmental Analysis of Contaminated Sites; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Pennacchio, M.; Jefferson, L.V.; Havens, K. Arabidopsis thaliana: A new test species for phytotoxic bioassays. J. Chem. Ecol. 2005, 31, 1877–1885. [Google Scholar] [CrossRef]
- Asli, S.; Neumann, P.M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, S.; Lee, I. Alteration of phytotoxicity and oxidant stress potential by metal oxide nanoparticles in Cucumis sativus. Water Air Soil Pollut. 2012, 223, 2799–2806. [Google Scholar] [CrossRef]
- Feng, Y.; Cui, X.; He, S.; Dong, G.; Chen, M.; Wang, J.; Lin, X. The role of metal nanoparticles in influencing arbuscular mycorrhizal fungi effects on plant growth. Environ. Sci. Technol. 2013, 47, 9496–9504. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-J.; Kwak, J.I.; Lee, W.-M.; Holden, P.A.; An, Y.-J. Zinc oxide nanoparticles delay soybean development: A standard soil microcosm study. Ecotoxicol. Environ. Saf. 2014, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.D.; Morales, M.I.; Castillo-Michel, H.; Hernandez-Viezcas, J.A.; Munoz, B.; Zhao, L.; Nunez, J.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Synchrotron verification of TiO2 accumulation in cucumber fruit: A possible pathway of TiO2 nanoparticle transfer from soil into the food chain. Environ. Sci. Technol. 2013, 47, 11592–11598. [Google Scholar] [CrossRef]
- Lee, C.W.; Mahendra, S.; Zodrow, K.; Li, D.; Tsai, Y.C.; Braam, J.; Alvarez, P.J. Developmental phytotoxicity of metal oxide nanoparticles to Arabidopsis thaliana. Environ. Toxicol. Chem. Int. J. 2010, 29, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Khan, S.S.; Pakrashi, S.; Mukherjee, A.; Chandrasekaran, N. Cytogenetic and genotoxic effects of zinc oxide nanoparticles on root cells of Allium cepa. J. Hazard. Mater. 2011, 190, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Bandyopadhyay, M.; Mukherjee, A. Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: Plant and human lymphocytes. Chemosphere 2010, 81, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Chhikara, S.; Xing, B.; Musante, C.; White, J.C.; Dhankher, O.P. Physiological and molecular response of Arabidopsis thaliana (L.) to nanoparticle cerium and indium oxide exposure. ACS Sustain. Chem. Eng. 2013, 1, 768–778. [Google Scholar] [CrossRef]
- Khan, A.R.; Wakeel, A.; Muhammad, N.; Liu, B.; Wu, M.; Liu, Y.; Ali, I.; Zaidi, S.H.R.; Azhar, W.; Song, G. Involvement of ethylene signaling in zinc oxide nanoparticle-mediated biochemical changes in Arabidopsis thaliana leaves. Environ. Sci. Nano 2019, 6, 341–355. [Google Scholar] [CrossRef]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.-I.; Lin, X. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Feizi, H.; Moghaddam, P.R.; Shahtahmassebi, N.; Fotovat, A. Impact of bulk and nanosized titanium dioxide (TiO2) on wheat seed germination and seedling growth. Biol. Trace Elem. Res. 2012, 146, 101–106. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| ZnO In Vitro | |||||
|---|---|---|---|---|---|
| Study | Method | Cell Model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [94] | RNAseq | Human lung epithelial carcinoma cells (A549) | Uncoated 42 nm | 15 μg/mL for 1, 6 or 24 h | Enriched terms “response to metal ions”, “metallothioneins bind metals”, “apoptosis” and “immune system” ( at 6 &24 h); ↓ molecules related to DNA repair; Nrf2 pathway was predicted to be activated at 6 h but repressed at 24 h |
| [100] | Microarray | Phorbol 12-myristate 13-acetate (PMA)-differentiated THP-1 macrophages | Uncoated, <50 nm | 2 or 8 μg/mL for 4 h | Affected genes involved in metal metabolism, transcription regulation, DNA binding, protein synthesis and structure; at higher dose, altered gene expression involved in inflammation, apoptosis and mitochondrial dysfunction |
| [107] | Microarray | Rat alveolar macrophages (NR8383) | Uncoatad, 158 nm | 4 and 17 μg/mL for 4 h | Disturbed protein synthesis/homeostasis with the eIF2 and VEGF signaling pathways, stress response with mitochondrial dysfunction, and sirtuin signaling; ↑ metallothioneins, genes related to membrane damage sensor, lung fibrosis, and protein synthesis regulator; ↓ stress response mediator, cell-cycle regulator, and transcription factor |
| [108] | Microarray | Human chronic myeloid leukemia cells (K562 cell line) | Uncoated, ≤40 nm | 40 μg/mL for 15 h | ↑ Genes involved in “response to zinc ions”, “detoxification of inorganic compound”, and “negative regulation of growth”; ↓ genes that regulated “immune responses”, “cell proliferation/migration”, “receptor signaling pathway via JAK-STAT” and “phosphatidylinositol 3-kinase signaling”; ↑ anti-oxidant defense system, mitochondrial-dependent apoptosis, and ↓ NF-κB pathway |
| [109] | RNAseq | Human skin cancer cells (A431) | Uncoated, around 500 nm | 150 μg/mL for 6 h | Altered gene expression for pathways in cancer, alcoholism, environmental information processing including MAPK, cytokine, TNF signaling pathways; ↑ genes related to injured or inflamed skin, and ↓ genes of apoptosis/cell cycle/cell survival |
| [110] | Microarray | Human monocyte-derived macrophages; Jurkat T cell leukemia derived cell | Uncoated, 15 nm | 1 or 10 μg/mL for 6 or 24 h | Affected cell death, cell growth, immune system development processes |
| ZnO In Vitro | |||||
| Study | Method | Animal model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [105] | Microarray (lung) | C57BL/6J BomTac female mice | Uncoated, 100 nm | Intratracheal instillation at 11, 33 or 100 mg/kg once | Enriched pathways for cell cycle G2 to M phase DNA damage checkpoint regulation, circadian rhythm signaling, protein ubiquitination pathway, unfolded protein response, and AMPK signaling |
| [111] | RNAseq (liver) | CD-1 male mice | Around 35 nm | Oral administration at 25 mg/kg for 8 or 12 weeks | Most significantly enriched Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways involved membranes, endoplasmic reticulum stress and ROS generation |
| [112] | RNAseq (liver) | Sprague Dawley female rats | Uncoated, 86.3 nm | Oral administration at 100 mg/kg for 14 consecutive days | ↑ Metabolic process and metal binding in liver |
| [113] | RNAseq (cultured skin cell) | CD-1 mice | Around 30 nm | Mouse hair follicle stem cells were exposed at 20 μg/mL for 12 h | Perturbed genes associated with hair follicle stem cell apoptosis and differentiation; altered pathways involved in cellular communication and RNA biosynthetic processes |
| [114] | RNAseq (liver) | Hairless SKH:QS mice | Uncoated, 18.2 ± 0.4 nm | Dermal application at 2 mg/cm2 to the head, ears, back, sides and tail, for 30 treatments | No statistically significant DEGs |
| TiO2 In Vitro | |||||
|---|---|---|---|---|---|
| Study | Method | Cell Model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [115] | Microarray | Undifferentiated Caco-2 cells | E171; Antase, 15–25 nm | 1.4 μg/cm2 for 2, 4, and 24 h | E171 and TiO2 NPs ↑ genes for inflammation, immune system, transport and cancer; E171 ↑ metabolism of proteins with the insulin processing pathway; TiO2 NPs affected pathways involved in metabolism of amino acids, prostaglandin, urea cycle, oxidative stress; two common biological processes: transport of molecules and neuronal system |
| [116] | RNAseq | Human lung epithelial carcinoma cells (A549) | Anatase (80%) and rutile (20%), 21 nm | 800 μg/mL for 24 h | ↑ Genes related to inflammatory response, cell surface signaling, oxidative stress, extracellular organization, electron transport, respiratory chain complex, and metabolic processes; ↓ genes that control cell cycle, secretion and cell–cell communication |
| [117] | RNAseq | Human glioblastoma cells (T98G) | 18 nm | 20 μg/mL for 72 h | Altered biological processes and functions were “granulocyte chemotaxis”, “response to lipopolysaccharide”, “response to cytokine”; enriched pathways “interleukin signaling”, “chemokine and cytokine signaling”, “B-cell activation” and “T-cell activation”, “cadherin signaling” and “integrin signaling” |
| TiO2 In Vitro | |||||
| Study | Method | Animal model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [118] | RNAseq (colon) | BALB/c male and female mice | E171 | Oral administration at 5 mg/kg for 2, 7, 14 or 21 days | ↓ Genes involved in innate and adaptive immune system; modulated signalling genes involved in colorectal cancer and biotransformation of xenobiotics |
| [119] | RNAseq (liver) | CD-1 mice | Anatase (80%) and rutile (20%), 21 nm | Oral administration at 50 mg/kg for 26 weeks | Most significantly enriched GO terms and KEGG pathways included plasma glucose homeostasis, metabolic mechanisms, generation of ROS, endoplasmic reticulum stress, and unfolded protein |
| [120] | Microarray (liver) | CD-1 female mice | Anatase, 5–6 nm | Oral administration at 10 mg/kg for 90 days | Altered gene expression for inflammatory response, apoptosis, oxidative stress, metabolic process, signal transduction, cytoskeleton, ion transport, cell proliferation, and cell differentiation |
| [121] | Microarray (spleen) | CD-1 female mice | Anatase, 7 nm | Oral administration at 10 mg/kg for 90 days | Perturbed gene expression involved in immune responses, apoptosis, stress responses, metabolic processes, signal transduction, cytoskeleton, oxidative stress, ion transport, cell division, and translation |
| [122] | Microarray (ovary) | CD-1 female mice | Anatase, 6 nm | Oral administration at 10 mg/kg for 90 days | Significantly upregulated DEGs involving hormone levels and reproduction, immune and inflammatory responses, transcription, ion transport, regulation of cell proliferation, and oxidoreductase activity |
| [114] | RNAseq (liver) | hairless SKH:QS mice | Anatase (80%) and rutile (20%), 21 nm | Dermal application at 2 mg/cm2 to the head, ears, back, sides and tail, for 30 treatments | No significant changes |
| [123] | Microarray (lung) | CD-1 mice | Anatase (80%) and rutile (20%), 21 nm | Intratracheal instillation at 5, 20 or 50 mg/kg once | ↑ Enriched genes related with antigen presentation and induction of chemotaxis of immune cells; probably caused chronic inflammatory diseases through Th2-mediated pathway |
| [124] | Microarray (lung) | CD-1 male mice | Rutile, 21 nm | Intratracheal instillation of 0.1 or 0.5 mg once | ↑ Pathways including cell cycle, apoptosis, chemokines, and complement cascades; ↑ genes in placenta growth factor and other chemokines expressions that may cause pulmonary emphysema and alveolar epithelial cell apoptosis |
| [125] | Microarray (lung and liver) | C57BL/6BomTac female mice | Rutile, 20 nm, coated with polyalcohols | Whole-body inhalation at 42 mg/m3 for 11 days (1 h/day) | ↑ Genes associated with acute phase, inflammation and immune response; associated pathways included cytokine–cytokine receptor interaction, metabolism, complement and coagulation cascade, hematopoeitic cell lineage, and biosynthesis of steroids |
| [126] | Microarray (lung) | CD-1 female mice | Anatase, 6 nm | Nasal instillation at 2.5, 5 or 10 mg/kg for 90 days | ↑ Genes involved in immune/inflammatory responses, apoptosis, oxidative stress, cell cycle, metabolic processes, stress responses, signal transduction, and cell differentiation |
| [127] | Microarray (lung) | C57BL/6 female mice | Rutile, 21 nm, coated with polyalcohols | Intratracheal instillation at 18, 54 or 162 μg/mouse once | ↑ Inflammatory gene expression; ↓ genes involved in ion homeostasis and muscle regulation |
| [128] | Microarray (lung) | C57BL/6J female mice | Anatase, rutile or anatase/rutile; 8, 20 and 300 nm; and hydrophobic or hydrophilic surface modifications | Intratracheal instillation at 18, 54, 162 or 486 μg/mouse once | Rutile type induced higher number of DEGs relate to inflammataion and acute phase signaling; hydrophilic surface induced higher DEGs; among the anatase, the smallest type showed the maximum response; anatase types enriched inflammatory response, response to wounding, defense response, chemotaxis; high dose of anatase TiO2 affected cytokine–cytokine receptor interaction, chemokine signalling, NOD-like receptor signalling, p53 signalling, ataxia telangiectasia mutated signalling, and steroid metabolic process |
| [129] | Microarray (liver and heart) | C57BL/6 female mice | Rutile, 21 nm, coated with polyalcohols | Intratracheal instillation at 162 μg/mouse once | ↑ Complement cascade and inflammatory processes in heart for particle opsonisation and clearance; mild changes in gene associated with acute phase responses in liver |
| [130] | Microarray (liver) | C57BL/6BomTac female mice | Rutile, 21 nm, coated with polyalcohols | Whole-body inhalation at 42 mg/m3 for 10 days (1 h/day) during gestation | Altered gene expression related to the retinoic acid signalling pathway in the female newborn livers; associated pathways related tissue development and vitamin, mineral and lipid metabolism |
| [131] | RNAseq (heart) | Sprague Dawley female rats | Anatase (80%) and rutile (20%), 21 nm | Whole-body inhalation at 10 mg/m3 for 7–8 days (4–6 h/day) during gestation | Altered pathways involved in inflammatory signaling and organismal development; ↓ protein kinase B (AKT) signaling; ↑ IL-8 signaling |
| ZnO | |||||
| Study | Method | Ecotox Model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [185] | Microarray | Zebrafish | <50 nm | 4.8 mg/L for 96 h | Mainly affected nucleic acid metabolism via altering nucleic acid binding; enriched KEGG pathways included “cell cycle”, “DNA replication”, and “homologous recombination” |
| [186] | Microarray | Zebrafish | Uncoated, 20–30 nm | 0.01, 0.1, 1 or 10 mg/L for 96 h post-fertilization | ↑ Genes for inflammation and the immune system; toxicological pathways included cytokine-cytokine receptor interactions and the intestinal immune network for IgA production |
| [187] | Microarray | Zebrafish larva | Uncoated, 10–30 nm | 1 or 4 μmol/L for 72 h post-fertilization | ↑ Cell differentiation and pathways associated with the immune system; ↑ several key genes involved in cancer cell signaling |
| [188] | Microarray | Caenorhabditis elegans | Pristine, phosphatized or sulfidized, 30 nm | 0.7 mg/L (ZnO), 7.5 mg/L (pZnO) and 7.5 mg/L (sZnO) for 48 h | Induced DEGs related to metal responsive genes; enriched pathways for protein biosynthesis (Aminoacyl-tRNA biosynthesis) and associated with detoxification (ABC transporters) were shared among pristine and one or both transformed ZnO NPs |
| [189] | Microarray (roots) | Arabidopsis thaliana | Uncoated, <100 nm | 100 mg/L for 7 days | Mainly perturbed genes involved in stress responses to abiotic (oxidative, salt, water deprivation) and biotic (wounding and defense to pathogens) stimuli; ↑ genes involved in cellular metal ion homeostasis and transport, and enzymes against oxidative stress; ↓ genes related to cell organization and biogenesis, translation, nucleosome assembly and microtubule-based process |
| [190] | Microarray | Arabidopsis thaliana | Uncoated, 20 nm | 4 mg/L for 7 days | ↑ Genes for stress responses (e.g., to salt, osmotic stress or water deprivation), responses to pathogens, oxidative stress, transcription factors, and transporters; ↓ genes involved in cell organization and biogenesis, nucleic acid metabolism, ribosomal proteins, cell wall modification and cell growth |
| TiO2 | |||||
| Study | Method | Ecotox Model | Material Properties | Exposure Conditions | Main Transcriptomic Findings (↑ Means Upregulate, Increase, Stimulate; ↓ Means Downregulate, Decrease or Suppress) |
| [191] | Microarray | Zebrafish embryos | Anatase, 25 nm | Microinjections of 8.5 ng/g | Interfered pathways related to circadian rhythm, cell signaling through kinase-related activities, trafficking of Golgi vesicles, immune function, and exocytosis |
| [192] | Microarray (ovary) | Zebrafish | Anatase, <25 nm | 0.1 and 1 mg/L for 13 weeks | Perturbed expresssion of genes involved in proteolysis, oxidative stress regulation, metabolism, insulin signaling, apoptosis and oocyte maturation; ↑ genes associated with protein degradation or ROS production |
| [193] | Microarray | Caenorhabditis elegans | Anatase, 32 nm | 200 μg/mL for 72 h | Affected genes involved in metal binding/detoxification, fertility and reproduction, worm growth, body morphogenesis, and neuronal function |
| [194] | Microarray | Caenorhabditis elegans | Anatase (83%) and rutile (17%), 34.1 nm; anatase: 5.9–16.2 nm; rutile: 12.6–68.9 nm | 0.01, 0.1, 1 and 10 mg/L for 24 h | Altered regulation of anti-oxidant system, stress resistance regulator and embryonic development; anatase type greatly influenced metabolic pathways whereas rutile particles significantly affected developmental processes |
| [189] | Microarray (root) | Arabidopsis thaliana | Anatase (80%) and rutile (20%), 21 nm | 100 mg/L for 7 days | Mild changes, primarily responses to biotic and abiotic stimuli |
| [195] | Microarray | Arabidopsis thaliana germinants | Anatase (80%) and rutile (20%), 21 nm | 500 mg/L for 12 days | ↑ Genes related to metabolic processes (DNA metabolism, hormone metabolism, triterpenoid biosynthesis and photosynthesis, indole glucosinolate metabolism, tryptophan catabolism), root development and cell differentiation, ion transport, and redox reaction; ↓ genes related to respiratory burst, responses to stress, hypoxia, and immune responses |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Alenius, H.; El-Nezami, H.; Karisola, P. A New Look at the Effects of Engineered ZnO and TiO2 Nanoparticles: Evidence from Transcriptomics Studies. Nanomaterials 2022, 12, 1247. https://doi.org/10.3390/nano12081247
Wang S, Alenius H, El-Nezami H, Karisola P. A New Look at the Effects of Engineered ZnO and TiO2 Nanoparticles: Evidence from Transcriptomics Studies. Nanomaterials. 2022; 12(8):1247. https://doi.org/10.3390/nano12081247
Chicago/Turabian StyleWang, Shuyuan, Harri Alenius, Hani El-Nezami, and Piia Karisola. 2022. "A New Look at the Effects of Engineered ZnO and TiO2 Nanoparticles: Evidence from Transcriptomics Studies" Nanomaterials 12, no. 8: 1247. https://doi.org/10.3390/nano12081247
APA StyleWang, S., Alenius, H., El-Nezami, H., & Karisola, P. (2022). A New Look at the Effects of Engineered ZnO and TiO2 Nanoparticles: Evidence from Transcriptomics Studies. Nanomaterials, 12(8), 1247. https://doi.org/10.3390/nano12081247