
Dietary Transfer of Zinc Oxide Nanoparticles Induces Locomotive Defects Associated with GABAergic Motor Neuron Damage in Caenorhabditis elegans



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Characterization, and Strains Maintenance
2.2. Measurement of Zn Concentration in E. coli OP50
2.3. Dietary Transfer Assay
2.4. Locomotive Behaviors Tests
2.5. GABAergic Neuron Toxicity Assay
2.6. Statistical Analysis
3. Results and Discussion
3.1. Concentration of Zn Released from ZnO-NPs in LB Medium
3.2. Zn Accumulation in E. coli OP50
3.3. Distribution and Accumulation of ZnO-NPs in C. elegans via Dietary Transfer
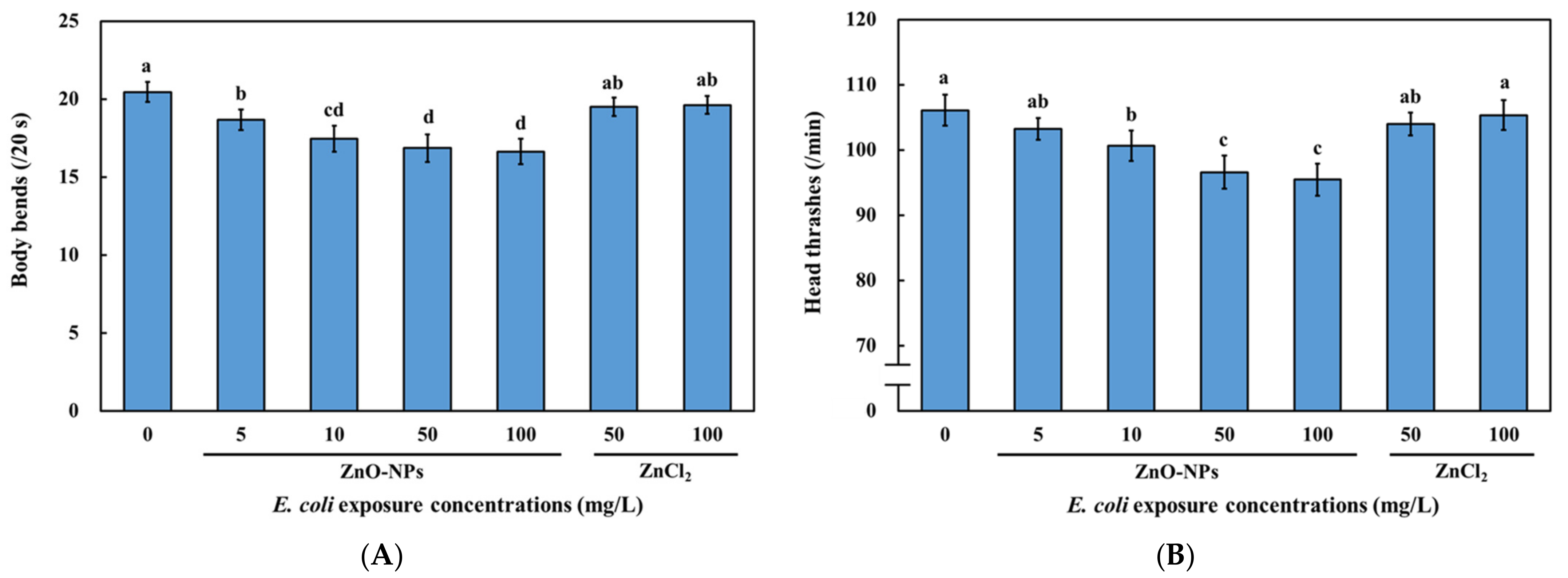
3.4. Effects of Dietary Transfer of ZnO-NPs on Locomotive Behaviors of C. elegans
3.5. Effects of Dietary Transfer of ZnO-NPs on D-Type GABAergic Motor Neurons of C. elegans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bundschuh, M.; Filser, J.; Luderwald, S.; Mckee, M.S.; Metreveli, G.; Schaumann, G.E.; Schulz, R.; Wagner, S. Nanoparticles in the environment: Where do we come from, where do we go to? Environ. Sci. Eur. 2018, 30, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangaa, S.R.; Selck, H.; Winther-Nielsen, M.; Khan, F.R. Trophic transfer of metal-based nanoparticles in aquatic environments: A review and recommendations for future research focus. Environ. Sci. Nano 2016, 3, 966–981. [Google Scholar] [CrossRef] [Green Version]
- Krol, A.; Pomastowski, P.; Rafinska, K.; Railean-Plugaru, V.; Buszewski, B. Zinc oxide nanoparticles: Synthesis, antiseptic activity and toxicity mechanism. Adv. Colloid Interface Sci. 2017, 249, 37–52. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Mandal, A.K.; Katuwal, S.; Tettey, F.; Gupta, A.; Bhattarai, S.; Jaisi, S.; Bhandari, D.P.; Shah, A.K.; Bhattarai, N.; Parajuli, N. Current research on zinc oxide nanoparticles: Synthesis, characterization and biomedical applications. Nanomaterials 2022, 12, 3066. [Google Scholar] [CrossRef]
- Moradpoor, H.; Safaei, M.; Mozaffari, H.R.; Sharifi, R.; Imani, M.M.; Golshah, A.; Bashardoust, N. An overview of recent progress in dental applications of zinc oxide nanoparticles. RSC Adv. 2021, 11, 21189–21206. [Google Scholar] [CrossRef]
- Jiang, J.; Pi, J.; Cai, J. The Advancing of zinc oxide nanoparticles for biomedical applications. Bioinorg. Chem. Appl. 2018, 2018, 1062562. [Google Scholar] [CrossRef]
- Rahman, H.S.; Othman, H.H.; Abdullah, R.; Edin, H.; Al-Haj, N.A. Beneficial and toxicological aspects of zinc oxide nanoparticles in animals. Vet. Med. Sci. 2022, 8, 1769–1779. [Google Scholar] [CrossRef]
- Goda, M.N.; Said, A.E.-A.A.; Abdelhamid, H.N. Highly selective dehydration of methanol over metal-organic frameworks (MOFs)-derived ZnO@Carbon. J. Environ. Chem. Eng. 2021, 9, 106336. [Google Scholar] [CrossRef]
- Soliman, A.I.A.; Abdel-Wahab, A.A.; Abdelhamid, H.N. Hierarchical porous zeolitic imidazolate frameworks (ZIF-8) and ZnO@N-doped carbon for selective adsorption and photocatalytic degradation of organic pollutants. RSC Adv. 2022, 12, 7075–7084. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, H.N.; Al Kiey, S.A.; Sharmoukh, W. A high-performance hybrid supercapacitor electrode based on ZnO/nitrogen-doped carbon nanohybrid. Appl. Organomet. Chem. 2022, 36, e6486. [Google Scholar] [CrossRef]
- Sun, T.Y.; Gottschalk, F.; Hungerbuhler, K.; Nowack, B. Comprehensive probabilistic modelling of environmental emissions of engineered nanomaterials. Environ. Pollut. 2014, 185, 69–76. [Google Scholar] [CrossRef]
- Dumont, E.; Johnson, A.C.; Keller, V.D.J.; Williams, R.J. Nano silver and nano zinc-oxide in surface waters—Exposure estimation for Europe at high spatial and temporal resolution. Environ. Pollut. 2015, 196, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.L.; Fang, C.M.; Pung, S.Y.; Ong, C.E.; Pung, Y.F.; Kong, C.; Pan, Y. Current updates on the in vivo assessment of zinc oxide nanoparticles toxicity using animal models. BioNanoScience 2021, 11, 590–620. [Google Scholar] [CrossRef]
- Suedel, B.C.; Boraczek, J.A.; Peddicord, R.K.; Clifford, P.A.; Dillon, T.M. Trophic transfer and biomagnification potential of contaminants in aquatic ecosystems. Rev. Environ. Contam. Toxicol. 1994, 136, 21–89. [Google Scholar]
- Fisher, N.S.; Hook, S.E. Toxicology tests with aquatic animals need to consider the trophic transfer of metals. Toxicology 2002, 181–182, 531–536. [Google Scholar] [CrossRef]
- Kalman, J.; Paul, K.B.; Khan, F.R.; Stone, V.; Fernandes, T.F. Characterisation of bioaccumulation dynamics of three differently coated silver nanoparticles and aqueous silver in a simple freshwater food chain. Environ. Chem. 2015, 12, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Y.; Li, H.R.; Han, X.Q.; Wei, X.Z. Transmission and accumulation of nano-TiO2 in a 2-step food chain (Scenedesmus obliquus to Daphnia magna). Bull. Environ. Contam. Toxicol. 2015, 95, 145–149. [Google Scholar] [CrossRef]
- Skjolding, L.M.; Winther-Nielsen, M.; Baun, A. Trophic transfer of differently functionalized zinc oxide nanoparticles from crustaceans (Daphnia magna) to zebrafish (Danio rerio). Aquat. Toxicol. 2014, 157, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Xu, S.M.; Yang, Y.N.; Li, L.Z.; Chen, S.P.; Xu, A.; Wu, L.J. Insights into the ecotoxicity of silver nanoparticles transferred from Escherichia coli to Caenorhabditis elegans. Sci. Rep. 2016, 6, 36465. [Google Scholar] [CrossRef]
- Kwak, J.I.; An, Y.J. Trophic transfer of silver nanoparticles from earthworms disrupts the locomotion of springtails (Collembola). J. Hazard. Mater. 2016, 315, 110–116. [Google Scholar] [CrossRef]
- Gambardella, C.; Gallus, L.; Gatti, A.M.; Faimali, M.; Carbone, S.; Antisari, L.V.; Falugi, C.; Ferrando, S. Toxicity and transfer of metal oxide nanoparticles from microalgae to sea urchin larvae. Chem. Ecol. 2014, 30, 308–316. [Google Scholar] [CrossRef]
- Sengul, A.B.; Asmatulu, E. Toxicity of metal and metal oxide nanoparticles: A review. Environ. Chem. Lett. 2020, 18, 1659–1683. [Google Scholar] [CrossRef]
- Von Mikecz, A. Lifetime eco-nanotoxicology in an adult organism: Where and when is the invertebrate C. elegans vulnerable? Environ. Sci. Nano 2018, 5, 616–622. [Google Scholar] [CrossRef]
- Piechulek, A.; von Mikecz, A. Life span-resolved nanotoxicology enables identification of age-associated neuromuscular vulnerabilities in the nematode Caenorhabditis elegans. Environ. Pollut. 2018, 233, 1095–1103. [Google Scholar] [CrossRef]
- Huang, C.W.; Li, S.W.; Liao, V.H.C. Chronic ZnO-NPs exposure at environmentally relevant concentrations results in metabolic and locomotive toxicities in Caenorhabditis elegans. Environ. Pollut. 2017, 220, 1456–1464. [Google Scholar] [CrossRef]
- Chen, T.H.; Lin, C.C.; Meng, P.J. Zinc oxide nanoparticles alter hatching and larval locomotor activity in zebrafish (Danio rerio). J. Hazard. Mater. 2014, 277, 134–140. [Google Scholar] [CrossRef]
- Du, M.; Wang, D. The neurotoxic effects of heavy metal exposure on GABAergic nervous system in nematode Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2009, 27, 314–320. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Wang, X.; Wu, Q.L.; Li, Y.P.; Tang, M.; Wang, D.Y. Quantum dots exposure alters both development and function of D-type GABAergic motor neurons in nematode Caenorhabditis elegans. Toxicol. Res. 2015, 4, 399–408. [Google Scholar] [CrossRef]
- Legradi, J.B.; Di Paolo, C.; Kraak, M.H.S.; van der Geest, H.G.; Schymanski, E.L.; Williams, A.J.; Dingemans, M.M.L.; Massei, R.; Brack, W.; Cousin, X.; et al. An ecotoxicological view on neurotoxicity assessment. Environ. Sci. Eur. 2018, 30, 46. [Google Scholar] [CrossRef]
- Leung, M.C.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Comber, S.D.; Conrad, A.U.; Hoss, S.; Webb, S.; Marshall, S. Chronic toxicity of sediment-associated linear alkylbenzene sulphonates (LAS) to freshwater benthic organisms. Environ. Pollut. 2006, 144, 661–668. [Google Scholar] [CrossRef]
- Amin, N.; Zulkifli, S.Z.; Azmai, M.N.A.; Ismail, A. Toxicity of zinc oxide nanoparticles on the embryo of Javanese medaka (Oryzias javanicus Bleeker, 1854): A comparative study. Animals 2021, 11, 2170. [Google Scholar] [CrossRef]
- Zhang, L.M.; Xia, J.G.; Zhao, Q.H.; Liu, L.W.; Zhang, Z.J. Functional graphene oxide as a nanocarrier for controlled loading and targeted delivery of mixed anticancer drugs. Small 2010, 6, 537–544. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tsalik, E.L.; Hobert, O. Functional mapping of neurons that control locomotory behavior in Caenorhabditis elegans. J. Neurobiol. 2003, 56, 178–197. [Google Scholar] [CrossRef]
- Gupta, G.S.; Dhawan, A. Chapter 17—Fate and potential hazards of nanoparticles in the environment. In Nanoparticle Therapeutics; Kesharwani, P., Singh, K.K., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 581–602. [Google Scholar]
- Matula, K.; Richter, L.; Adamkiewicz, W.; Akerstrom, B.; Paczesny, J.; Holyst, R. Influence of nanomechanical stress induced by ZnO nanoparticles of different shapes on the viability of cells. Soft Matter 2016, 12, 4162–4169. [Google Scholar] [CrossRef]
- Gelabert, A.; Sivry, Y.; Gobbi, P.; Mansouri-Guilani, N.; Menguy, N.; Brayner, R.; Siron, V.; Benedetti, M.F.; Ferrari, R. Testing nanoeffect onto model bacteria: Impact of speciation and genotypes. Nanotoxicology 2016, 10, 216–225. [Google Scholar] [CrossRef]
- Li, M.; Lin, D.H.; Zhu, L.Z. Effects of water chemistry on the dissolution of ZnO nanoparticles and their toxicity to Escherichia coli. Environ. Pollut. 2013, 173, 97–102. [Google Scholar] [CrossRef]
- Miao, A.J.; Zhang, X.Y.; Luo, Z.; Chen, C.S.; Chin, W.C.; Santschi, P.H.; Quigg, A. Zinc oxide-engineered nanoparticles: Dissolution and toxicity to marine phytoplankton. Environ. Toxicol. Chem. 2010, 29, 2814–2822. [Google Scholar] [CrossRef]
- Jiang, C.; Aiken, G.R.; Hsu-Kim, H. Effects of natural organic matter properties on the dissolution kinetics of zinc oxide nanoparticles. Environ. Sci. Technol. 2015, 49, 11476–11484. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Mielke, R.E.; Priester, J.H.; Werlin, R.A.; Gelb, J.; Horst, A.M.; Orias, E.; Holden, P.A. Differential growth of and nanoscale TiO2 accumulation in Tetrahymena thermophila by direct feeding versus trophic transfer from Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2013, 79, 5616–5624. [Google Scholar] [CrossRef] [Green Version]
- Siddiqi, K.S.; Husen, A. Properties of zinc oxide nanoparticles and their activity against microbes. Nanoscale Res. Lett. 2018, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano-Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brayner, R.; Ferrari-Iliou, R.; Brivois, N.; Djediat, S.; Benedetti, M.F.; Fievet, F. Toxicological impact studies based on Escherichia coli bacteria in ultrafine ZnO nanoparticles colloidal medium. Nano Lett. 2006, 6, 866–870. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Peralta-Videa, J.R.; Plascencia-Villa, G.; Jose-Yacaman, M.; Gardea-Torresdey, J.L. Comparative toxicity assessment of CeO2 and ZnO nanoparticles towards Sinorhizobium meliloti, a symbiotic alfalfa associated bacterium: Use of advanced microscopic and spectroscopic techniques. J. Hazard. Mater. 2012, 241, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; Calder, A.; Britt, D.W.; McLean, J.E.; Anderson, A.J. Responses of a soil bacterium, Pseudomonas chlororaphis O6 to commercial metal oxide nanoparticles compared with responses to metal ions. Environ. Pollut. 2011, 159, 1749–1756. [Google Scholar] [CrossRef]
- Jarvis, T.A.; Miller, R.J.; Lenihan, H.S.; Bielmyer, G.K. Toxicity of ZnO nanoparticles to the copepod Acartia tonsa, exposed through a phytoplankton diet. Environ. Toxicol. Chem. 2013, 32, 1264–1269. [Google Scholar] [CrossRef]
- Bhuvaneshwari, M.; Iswarya, V.; Vishnu, S.; Chandrasekaran, N.; Mukherjee, A. Dietary transfer of zinc oxide particles from algae (Scenedesmus obliquus) to daphnia (Ceriodaphnia dubia). Environ. Res. 2018, 164, 395–404. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, F.; Li, W.; Luan, T.; Lin, L. Comparison on the effects of water-borne and dietary-borne accumulated ZnO nanoparticles on Daphnia magna. Chemosphere 2017, 189, 94–103. [Google Scholar] [CrossRef]
- Huang, C.W.; Chang, C.H.; Li, S.W.; Yen, P.L.; Liao, V.H.C. Co-exposure to foodborne and waterborne ZnO nanoparticles in aquatic sediment environments enhances DNA damage and stress gene expression in freshwater Asian clam Corbicula fluminea. Environ. Sci. Nano 2020, 7, 1252–1265. [Google Scholar] [CrossRef]
- Ates, M.; Arslan, Z.; Demir, V.; Daniels, J.; Farah, I.O. Accumulation and toxicity of CuO and ZnO nanoparticles through waterborne and dietary exposure of goldfish (Carassius auratus). Environ. Toxicol. 2015, 30, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Xu, T.; Wu, S.; Lei, L.; He, D. Chronic exposure to graphene-based nanomaterials induces behavioral deficits and neural damage in Caenorhabditis elegans. J. Appl. Toxicol. 2017, 37, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Pluskota, A.; Horzowski, E.; Bossinger, O.; von Mikecz, A. In Caenorhabditis elegans nanoparticle-bio-interactions become transparent: Silica-nanoparticles induce reproductive senescence. PLoS ONE 2009, 4, e6622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.L.; Li, Y.X.; Li, Y.P.; Zhao, Y.L.; Ge, L.; Wang, H.F.; Wang, D.Y. Crucial role of the biological barrier at the primary targeted organs in controlling the translocation and toxicity of multi-walled carbon nanotubes in the nematode Caenorhabditis elegans. Nanoscale 2013, 5, 11166–11178. [Google Scholar] [CrossRef] [PubMed]
- Tseng, I.L.; Yang, Y.F.; Yu, C.W.; Li, W.H.; Liao, V.H.C. Phthalates induce neurotoxicity affecting locomotor and thermotactic behaviors and AFD neurons through oxidative stress in Caenorhabditis elegans. PLoS ONE 2014, 9, e99945. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yu, S.; Wu, Q.; Tang, M.; Pu, Y.; Wang, D. Chronic Al2O3-nanoparticle exposure causes neurotoxic effects on locomotion behaviors by inducing severe ROS production and disruption of ROS defense mechanisms in nematode Caenorhabditis elegans. J. Hazard. Mater. 2012, 219–220, 221–230. [Google Scholar] [CrossRef]
- Boxall, A.; Chaudhry, Q.; Sinclair, C.; Jones, A.; Aitken, R.; Jefferson, B.; Watts, C. Current and Future Predicted Environmental Exposure to Engineered Nanoparticles; Central Science Laboratory: York, UK, 2007. [Google Scholar]
- Yu, Q.; Zhang, Z.; Monikh, F.A.; Wu, J.; Wang, Z.; Vijver, M.G.; Bosker, T.; Peijnenburg, W.J.G.M. Trophic transfer of Cu nanoparticles in a simulated aquatic food chain. Ecotoxicol. Environ. Saf. 2022, 242, 113920. [Google Scholar] [CrossRef]
- Chen, P.; Martinez-Finley, E.J.; Bornhorst, J.; Chakraborty, S.; Aschner, M. Metal-induced neurodegeneration in C. elegans. Front. Aging Neurosci. 2013, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Li, N.; Sheng, W.; Ji, X.; Liang, X.; Kong, B.; Yin, P.; Li, Y.; Zhang, X.; Liu, K. Toxicity of different zinc oxide nanomaterials and dose-dependent onset and development of Parkinson’s disease-like symptoms induced by zinc oxide nanorods. Environ. Int. 2021, 146, 106179. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, H.; Fang, Y.; Li, K.; Tian, L.; Liu, X.; Zhang, W.; Tan, Y.; Lai, W.; Bian, L.; et al. Neurotoxicity and biomarkers of zinc oxide nanoparticles in main functional brain regions and dopaminergic neurons. Sci. Total Environ. 2020, 705, 135809. [Google Scholar] [CrossRef] [PubMed]
- Babayevska, N.; Przysiecka, Ł.; Iatsunskyi, I.; Nowaczyk, G.; Jarek, M.; Janiszewska, E.; Jurga, S. ZnO size and shape effect on antibacterial activity and cytotoxicity profile. Sci. Rep. 2022, 12, 8148. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Mishra, H.; Ekielski, A.; Talegaonkar, S.; Vaidya, B. Zinc oxide nanoparticles: A promising nanomaterial for biomedical applications. Drug Discov. 2017, 22, 1825–1834. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
How, C.M.; Huang, C.-W. Dietary Transfer of Zinc Oxide Nanoparticles Induces Locomotive Defects Associated with GABAergic Motor Neuron Damage in Caenorhabditis elegans. Nanomaterials 2023, 13, 289. https://doi.org/10.3390/nano13020289
How CM, Huang C-W. Dietary Transfer of Zinc Oxide Nanoparticles Induces Locomotive Defects Associated with GABAergic Motor Neuron Damage in Caenorhabditis elegans. Nanomaterials. 2023; 13(2):289. https://doi.org/10.3390/nano13020289
Chicago/Turabian StyleHow, Chun Ming, and Chi-Wei Huang. 2023. "Dietary Transfer of Zinc Oxide Nanoparticles Induces Locomotive Defects Associated with GABAergic Motor Neuron Damage in Caenorhabditis elegans" Nanomaterials 13, no. 2: 289. https://doi.org/10.3390/nano13020289
APA StyleHow, C. M., & Huang, C. -W. (2023). Dietary Transfer of Zinc Oxide Nanoparticles Induces Locomotive Defects Associated with GABAergic Motor Neuron Damage in Caenorhabditis elegans. Nanomaterials, 13(2), 289. https://doi.org/10.3390/nano13020289