On the Electrochemical Detection of Alpha-Fetoprotein Using Aptamers: DNA Isothermal Amplification Strategies to Improve the Performance of Weak Aptamers

 ,
, 
 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Modification of Tosylactivated Magnetic Particles with AFP
2.2. Binding Assays on AFP-Modified Magnetic Particles
2.3. TdT Amplification Study by Gel Electrophoresis
3. Results
3.1. Comparison of Binding Affinity in the Absence of DNA Amplification
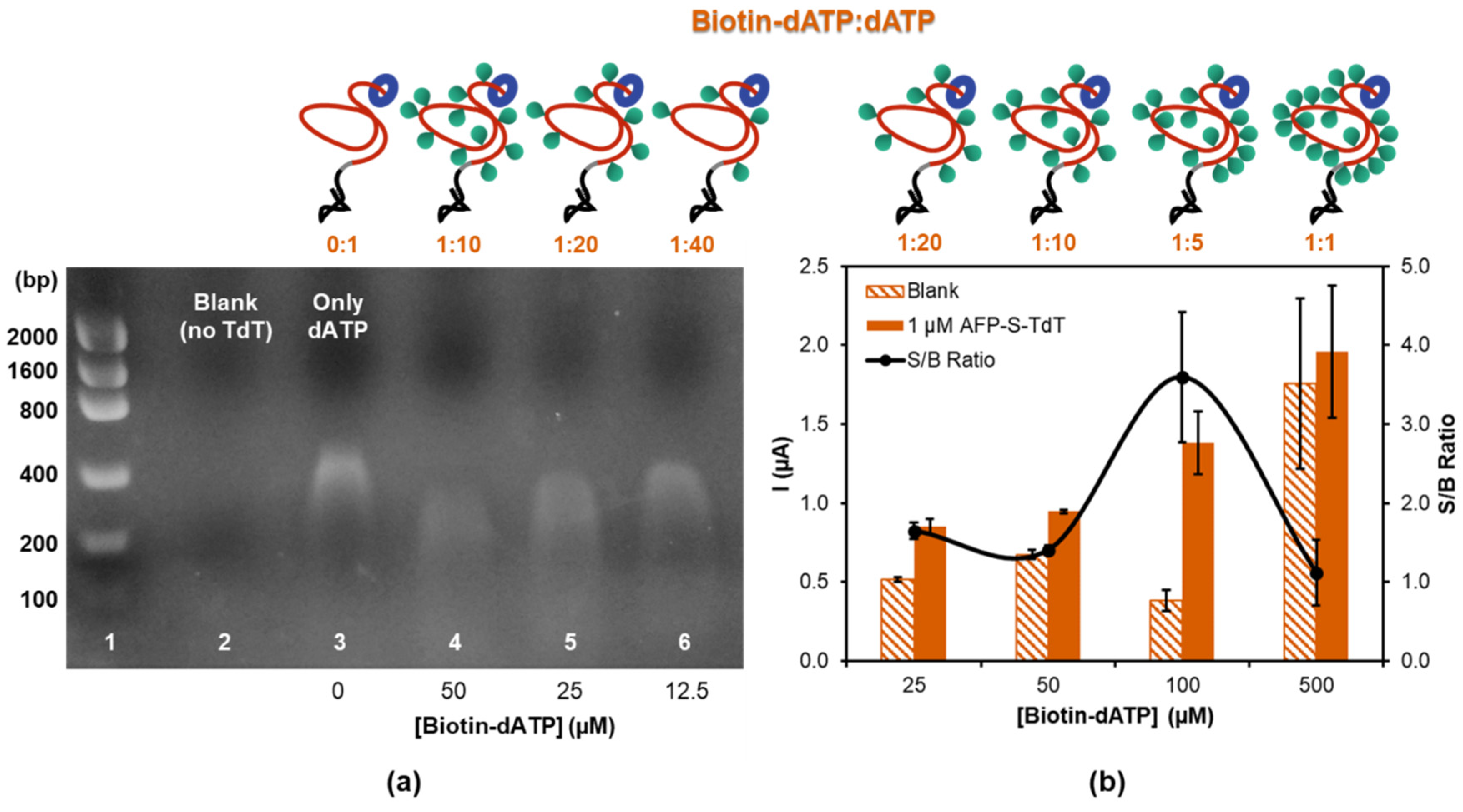
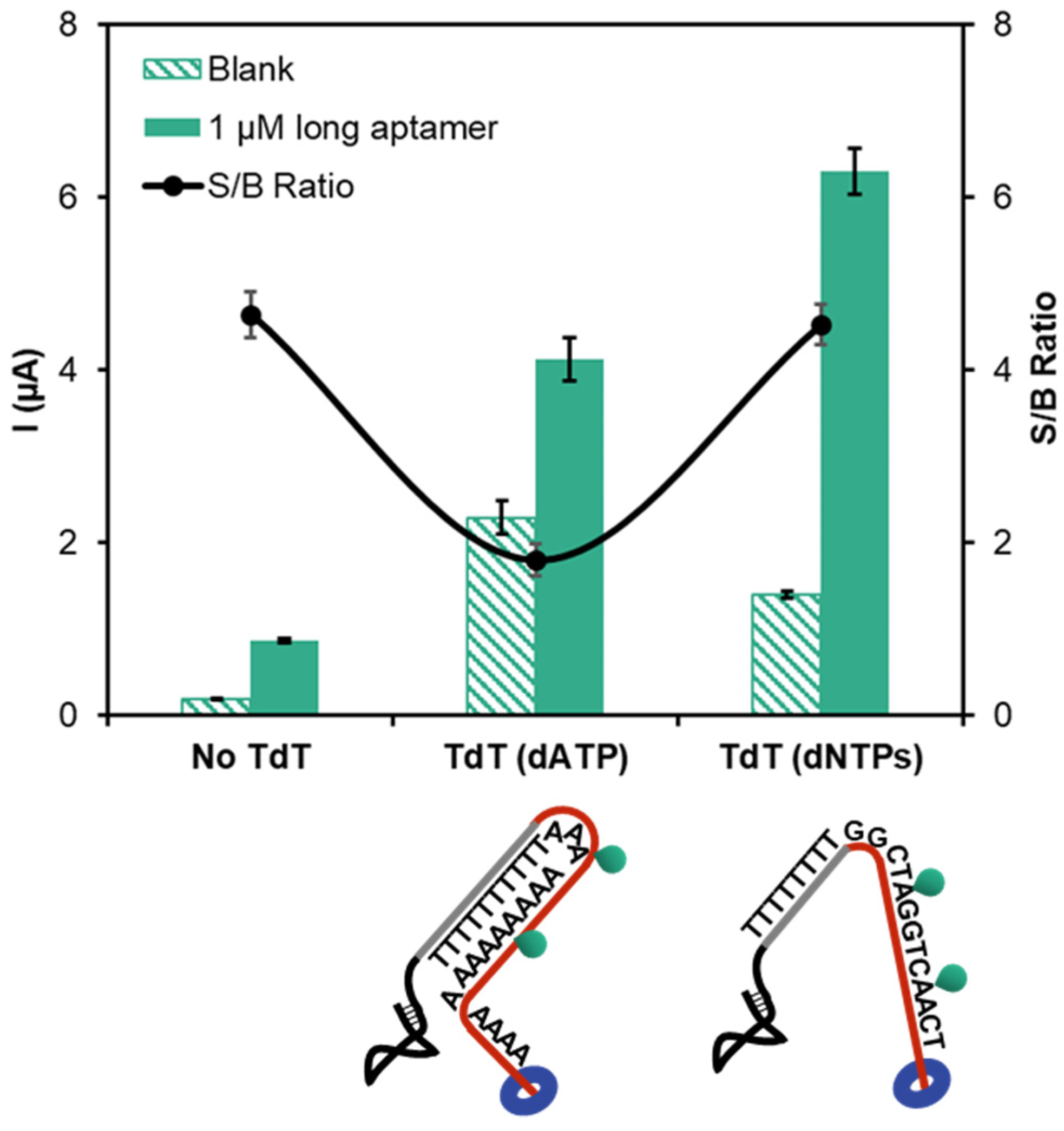
3.2. Amplification by Terminal Deoxynucleotidyl Transferase Elongation
3.3. Rolling Circle Amplification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diamandis, E.P. Cancer Biomarkers: Can We Turn Recent Failures into Success? J. Natl. Cancer Inst. 2010, 102, 1462–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisani, F.; Garuti, F.; Neri, A. Alpha-fetoprotein for Diagnosis, Prognosis, and Transplant Selection. Semin. Liver Dis. 2019, 39, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E.; Seppälä, M. α-Foetoprotein in Normal Human Serum. Nature 1972, 235, 161–162. [Google Scholar] [CrossRef] [PubMed]
- Galle, P.R.; Foerster, F.; Kudo, M.; Chan, S.L.; Llovet, J.M.; Qin, S.; Schelman, W.R.; Chintharlapalli, S.; Abada, P.B.; Sherman, M.; et al. Biology and Significance of alpha-Fetoprotein in Hepatocellular Carcinoma. Liver Int. 2019, 39, 2214–2229. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wei, C. Advances in the Early Diagnosis of Hepatocellular Carcinoma. Genes Dis. 2020. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Zhang, P.; Zhang, J.; Li, X.; Gan, D.; Cao, X.; Han, M.; Du, H.; Ye, Y. The Threshold of alpha-Fetoprotein (AFP) for the Diagnosis of Hepatocellular Carcinoma: A Systematic Review and Meta-analysis. PLoS ONE 2020, 15, e0228857. [Google Scholar] [CrossRef]
- Yue, Y.; Chen, X.; Zhang, S.; Xu, Z.; Wang, Q. Commutability of Reference Materials for α-Fetoprotein in Human Serum. Arch. Pathol. Lab. Med. 2017, 141, 1421–1427. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Lee, S.-W. Regression of Hepatocarcinoma Cells using RNA Aptamer Specific to alpha-Fetoprotein. Biochem. Biophys. Res. Commun. 2012, 417, 521–527. [Google Scholar] [CrossRef]
- Huang, C.-J.; Lin, H.-I.; Shiesh, S.-C.; Lee, G.-B. An Integrated Microfluidic System for Rapid Screening of alpha-Fetoprotein-specific Aptamers. Biosens. Bioelectron. 2012, 35, 50–55. [Google Scholar] [CrossRef]
- Dong, L.; Tan, Q.; Ye, W.; Liu, D.; Chen, H.; Hu, H.; Wen, D.; Liu, Y.; Cao, Y.; Kang, J.; et al. Screening and Identifying a Novel ssDNA Aptamer against Alpha-fetoprotein Using CE-SELEX. Sci. Rep. 2015, 5, 15552. [Google Scholar] [CrossRef]
- Wu, X.; Fu, P.; Ma, W.; Xu, L.; Kuang, H.; Xu, C. SERS-active Silver Nanoparticle Trimers for sub-attomolar Detection of alpha-Fetoprotein. RSC Adv. 2015, 5, 73395–73398. [Google Scholar] [CrossRef]
- Gui, R.; He, W.; Jin, H.; Sun, J.; Wang, Y. DNA Assembly of Carbon Dots and 5-Fluorouracil used for Room-Remperature Phosphorescence turn-on Sensing of AFP and AFP-triggered Simultaneous Release of Dual-drug. Sens. Actuators B Chem. 2018, 255, 1623–1630. [Google Scholar] [CrossRef]
- Heiat, M.; Negahdary, M. Sensitive Diagnosis of alpha-Fetoprotein by a Label-free Nanoaptasensor designed by modified Au Electrode with Spindle-shaped Gold Nanostructure. Microchem. J. 2019, 148, 456–466. [Google Scholar] [CrossRef]
- Li, G.; Li, S.; Wang, Z.; Xue, Y.; Dong, C.; Zeng, J.; Huang, Y.; Liang, J.; Zhou, Z. Label-free Electrochemical Aptasensor for Detection of alpha-Fetoprotein based on AFP-aptamer and Thionin/Reduced Graphene Oxide/Gold Nanoparticles. Anal. Biochem. 2018, 547, 37–44. [Google Scholar] [CrossRef]
- De Ávila, B.E.F.; Watkins, H.M.; Pingarrón, J.M.; Plaxco, K.W.; Palleschi, G.; Ricci, F. Determinants of the Detection Limit and Specificity of Surface-Based Biosensors. Anal. Chem. 2013, 85, 6593–6597. [Google Scholar] [CrossRef]
- Wu, Y.; Tilley, R.D.; Gooding, J.J. Challenges and Solutions in Developing Ultrasensitive Biosensors. J. Am. Chem. Soc. 2018, 141, 1162–1170. [Google Scholar] [CrossRef]
- Lorenzo-Gómez, R.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Lobo-Castañón, M.J. Electrochemical Aptamer-based Assays Coupled to Isothermal Nucleic Acid Amplification Techniques: New Tools for Cancer Diagnosis. Curr. Opin. Electrochem. 2019, 14, 32–43. [Google Scholar] [CrossRef]
- Li, F.; Zhang, H.; Wang, Z.; Newbigging, A.M.; Reid, M.S.; Li, X.-F.; Le, X.C. Aptamers Facilitating Amplified Detection of Biomolecules. Anal. Chem. 2014, 87, 274–292. [Google Scholar] [CrossRef]
- Barreda-García, S.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Miranda-Ordieres, A.J.; Lobo-Castañón, M.J. Solid-Phase Helicase Dependent Amplification and Electrochemical Detection of Salmonella on highly Stable Oligonucleotide-modified ITO Electrodes. Chem. Commun. 2017, 53, 9721–9724. [Google Scholar] [CrossRef]
- Barreda-Garcia, S.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Miranda-Ordieres, A.J.; Lobo-Castañón, M.J. Helicase-Dependent Isothermal Amplification: A Novel Tool in the Development of Molecular-based Analytical Systems for Rapid Pathogen Detection. Anal. Bioanal. Chem. 2017, 410, 679–693. [Google Scholar] [CrossRef]
- Sánchez-Salcedo, R.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Lobo-Castañón, M.J. On-Gold Recombinase Polymerase Primer Elongation for Electrochemical Detection of Bacterial Genome: Mechanism Insights and Influencing Factors. ChemElectroChem 2018, 6, 793–800. [Google Scholar] [CrossRef]
- Deng, H.; Gao, Z. Bioanalytical Applications of Isothermal Nucleic Acid Amplification Techniques. Anal. Chim. Acta 2015, 853, 30–45. [Google Scholar] [CrossRef]
- Anne, A.; Bonnaudat, C.; Demaille, C.; Wang, K. Enzymatic Redox 3‘-End-Labeling of DNA Oligonucleotide Monolayers on Gold Surfaces Using Terminal Deoxynucleotidyl Transferase (TdT)-Mediated Single Base Extension. J. Am. Chem. Soc. 2007, 129, 2734–2735. [Google Scholar] [CrossRef]
- Motea, E.A.; Berdis, A.J. Terminal Deoxynucleotidyl Transferase: The Story of a Misguided DNA Polymerase. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2009, 1804, 1151–1166. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, B.; Wiltshire, S.; Lambert, J.; O’Malley, S.; Kukanskis, K.; Zhu, Z.; Kingsmore, S.F.; Lizardi, P.M.; Ward, D.C. Immunoassays with Rolling Circle DNA Amplification: A versatile Platform for Ultrasensitive Antigen Detection. Proc. Natl. Acad. Sci. USA 2000, 97, 10113–10119. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Bian, S.; Yang, Y.; Wu, X. Chiroplasmonic Assemblies of Gold Nanoparticles as a Novel Method for Sensitive Detection of alpha-Fetoprotein. Microchim. Acta 2017, 10, 871–1862. [Google Scholar] [CrossRef]
- Gao, T.; Zhi, J.; Mu, C.; Gu, S.; Xiao, J.; Yang, J.; Wang, Z.; Xiang, Y. One-Step Detection for Two Serological Biomarker Species to improve the Diagnostic Accuracy of Hepatocellular Carcinoma. Talanta 2018, 178, 89–93. [Google Scholar] [CrossRef]
- Fowler, J.D.; Suo, Z. Biochemical, Structural, and Physiological Characterization of Terminal Deoxynucleotidyl Transferase. Chem. Rev. 2006, 106, 2092–2110. [Google Scholar] [CrossRef]
- Jensen, M.; Davis, R.W. Template-Independent Enzymatic Oligonucleotide Synthesis (TiEOS): Its History, Prospects, and Challenges. Biochemistry 2018, 57, 1821–1832. [Google Scholar] [CrossRef]
- Hermanová, M.; Havranová-Vidláková, P.; Ondráčková, A.; Kumar, S.S.; Bowater, R.; Fojta, M. Label-free Voltammetric Detection of Products of Terminal Deoxynucleotidyl Transferase Tailing Reaction. Electroanalysis 2018, 31, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Gómez, R.; Fernández-Alonso, N.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Lobo-Castañón, M.J. Unravelling the Lipocalin 2 Interaction with Aptamers: May Rolling Circle Amplification improve Their Functional Affinity? Talanta 2019, 197, 406–412. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, E.; de-los-Santos-Álvarez, N.; Miranda-Ordieres, A.J.; Lobo-Castañón, M.J. SPR Evaluation of Binding Kinetics and Affinity study of Modified RNA Aptamers Towards Small Molecules. Talanta 2012, 99, 767–773. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenzo-Gómez, R.; González-Robles, D.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Lobo-Castañón, M.J. On the Electrochemical Detection of Alpha-Fetoprotein Using Aptamers: DNA Isothermal Amplification Strategies to Improve the Performance of Weak Aptamers. Biosensors 2020, 10, 46. https://doi.org/10.3390/bios10050046
Lorenzo-Gómez R, González-Robles D, Miranda-Castro R, de-los-Santos-Álvarez N, Lobo-Castañón MJ. On the Electrochemical Detection of Alpha-Fetoprotein Using Aptamers: DNA Isothermal Amplification Strategies to Improve the Performance of Weak Aptamers. Biosensors. 2020; 10(5):46. https://doi.org/10.3390/bios10050046
Chicago/Turabian StyleLorenzo-Gómez, Ramón, Daniel González-Robles, Rebeca Miranda-Castro, Noemí de-los-Santos-Álvarez, and María Jesús Lobo-Castañón. 2020. "On the Electrochemical Detection of Alpha-Fetoprotein Using Aptamers: DNA Isothermal Amplification Strategies to Improve the Performance of Weak Aptamers" Biosensors 10, no. 5: 46. https://doi.org/10.3390/bios10050046
APA StyleLorenzo-Gómez, R., González-Robles, D., Miranda-Castro, R., de-los-Santos-Álvarez, N., & Lobo-Castañón, M. J. (2020). On the Electrochemical Detection of Alpha-Fetoprotein Using Aptamers: DNA Isothermal Amplification Strategies to Improve the Performance of Weak Aptamers. Biosensors, 10(5), 46. https://doi.org/10.3390/bios10050046