Detection of Exosomes Using Total Internal Reflected Imaging Ellipsometry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
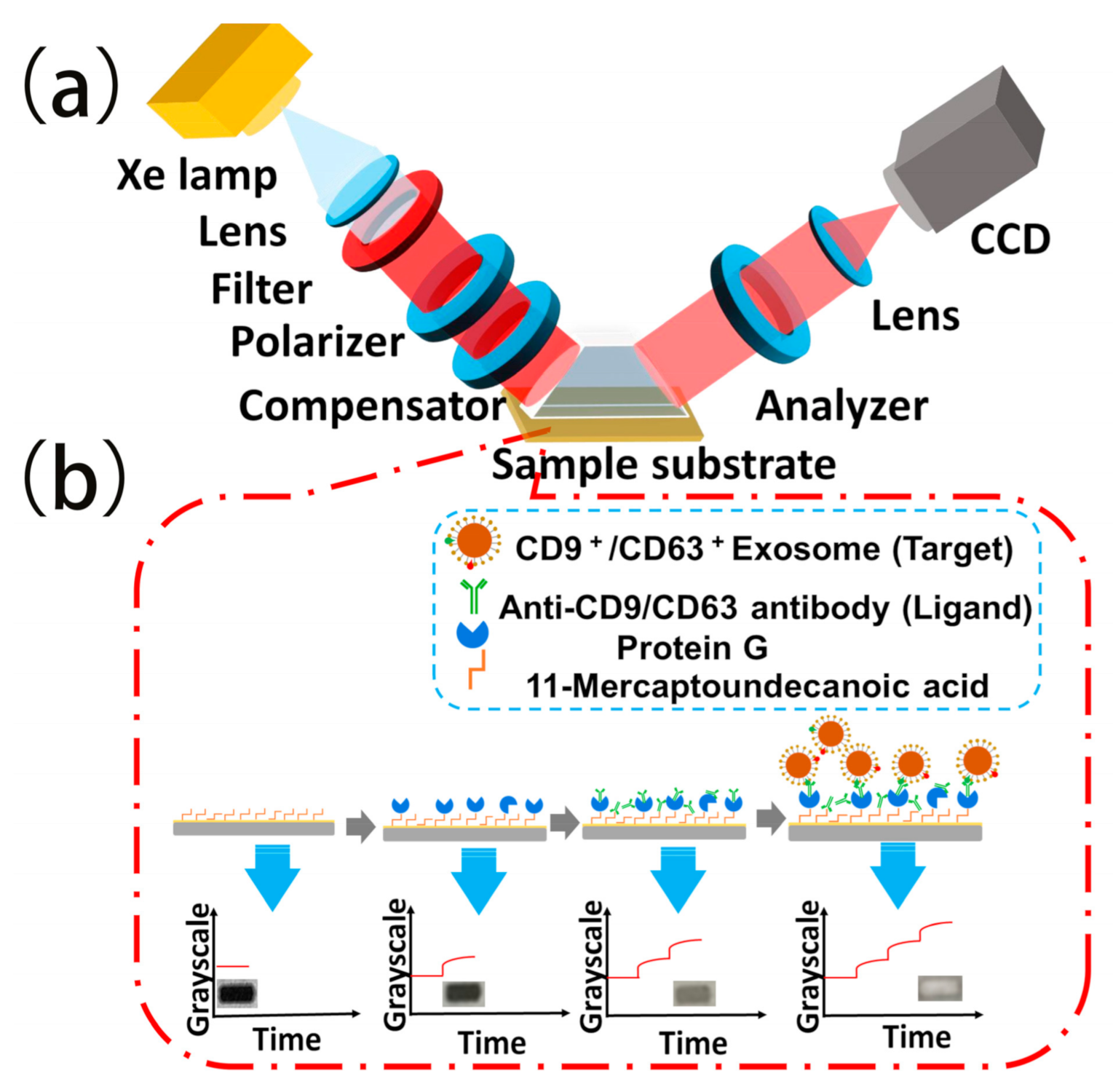
2.1. TIRIE
2.2. Chemicals and Biological Samples
2.3. Functionalization of Sensing Surface
2.4. The Surface Processing of Biomolecules
2.5. Dynamic Light Scattering Measurements
3. Results and Discussion
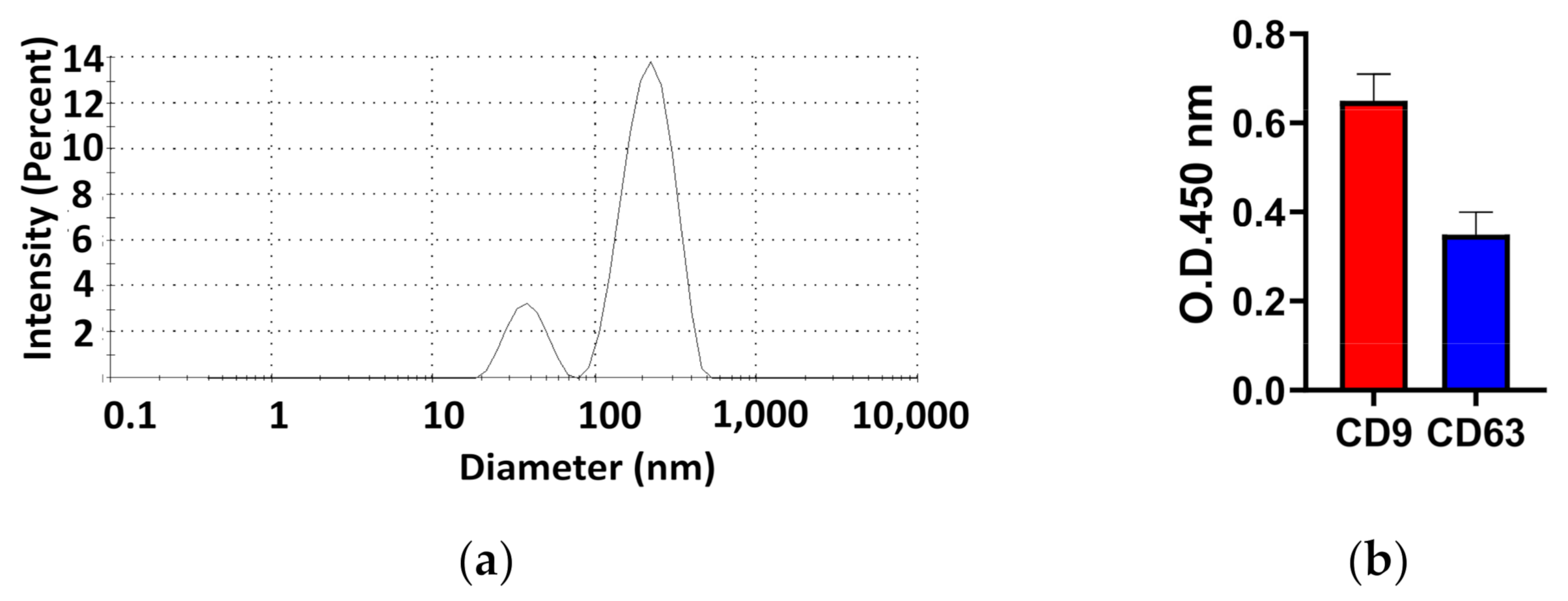
3.1. Characterization of the Diameter and Surface Protein of Exosomes
3.2. Design of Sensing Surface
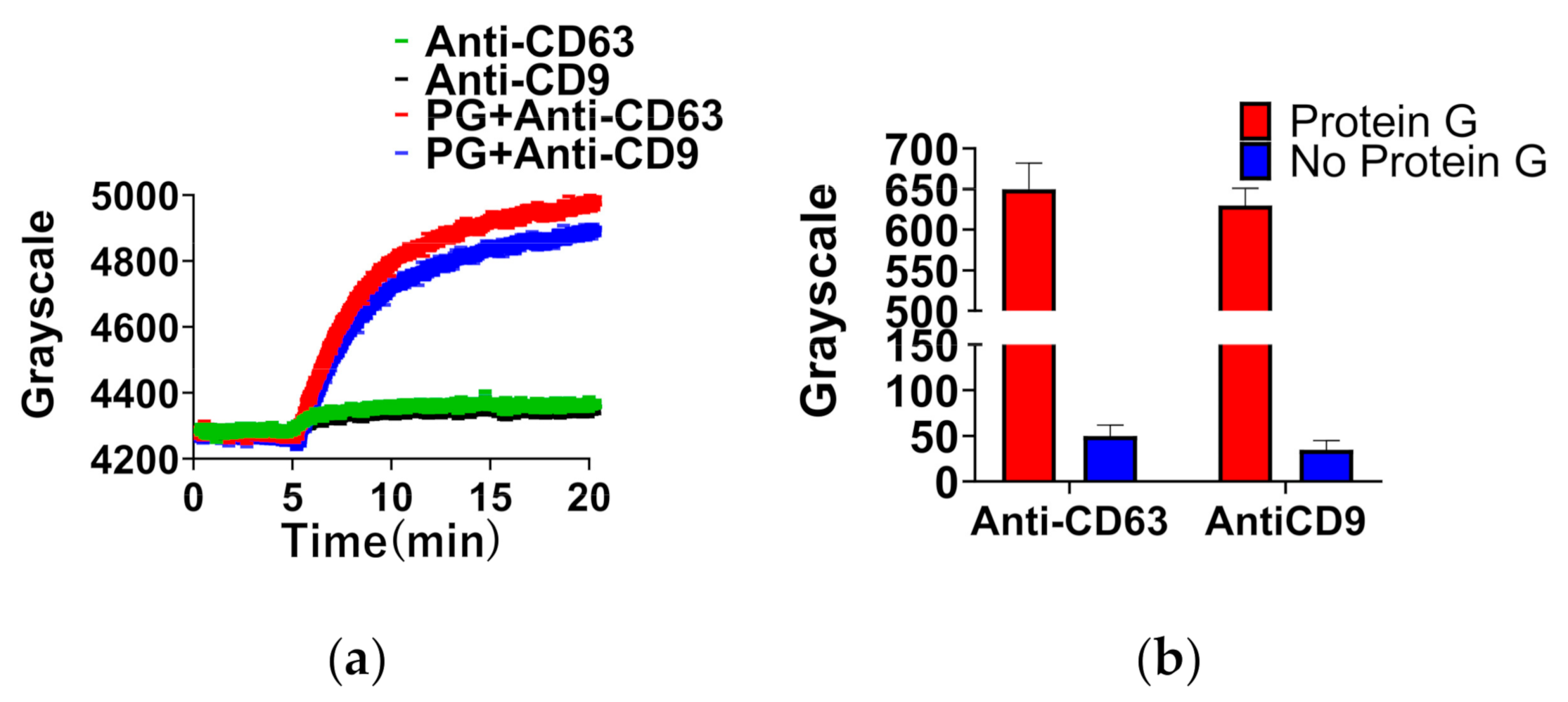
3.2.1. Immobilization of Antibody by Protein G
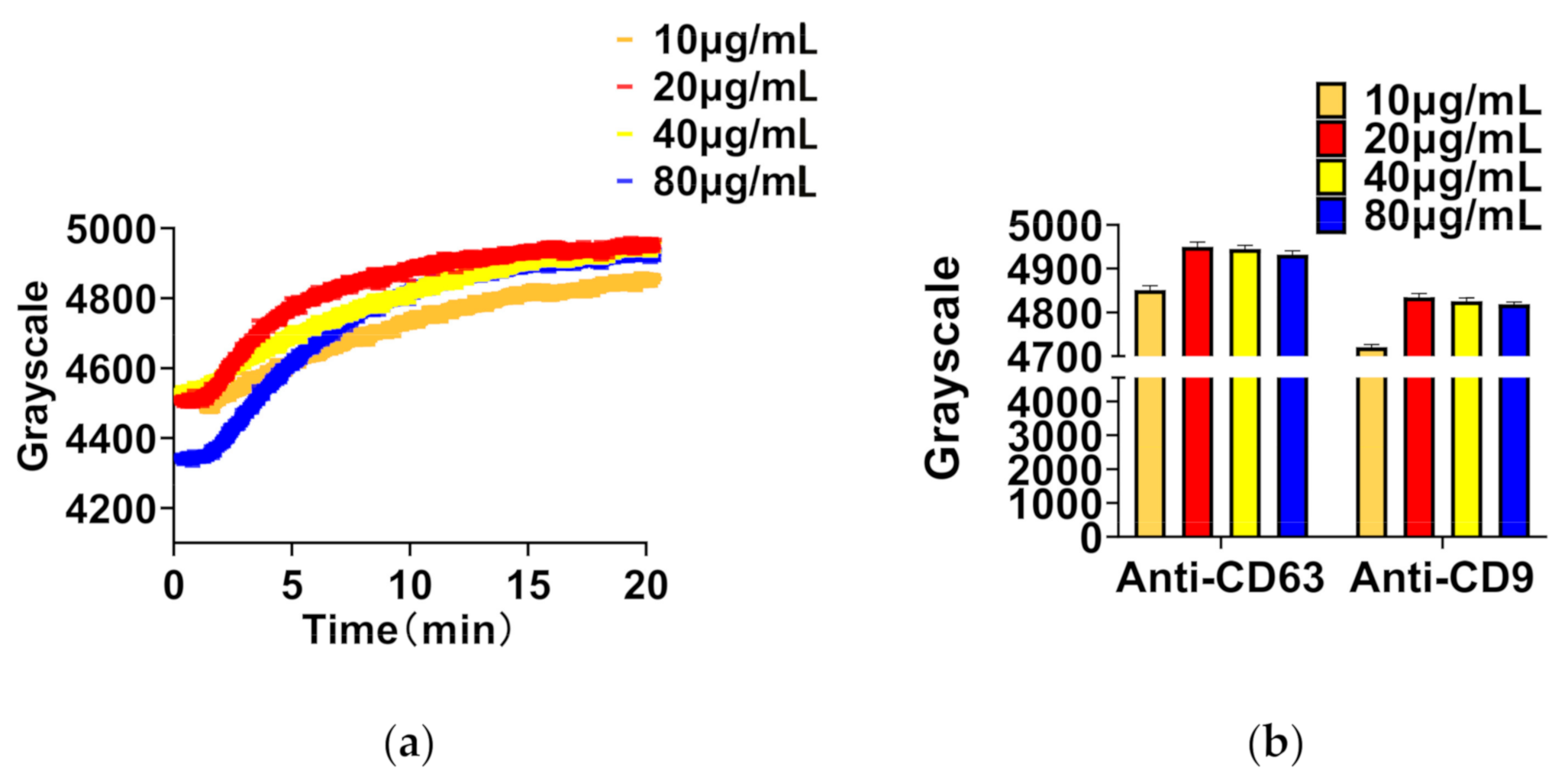
3.2.2. Optimum Concentration of Antibody
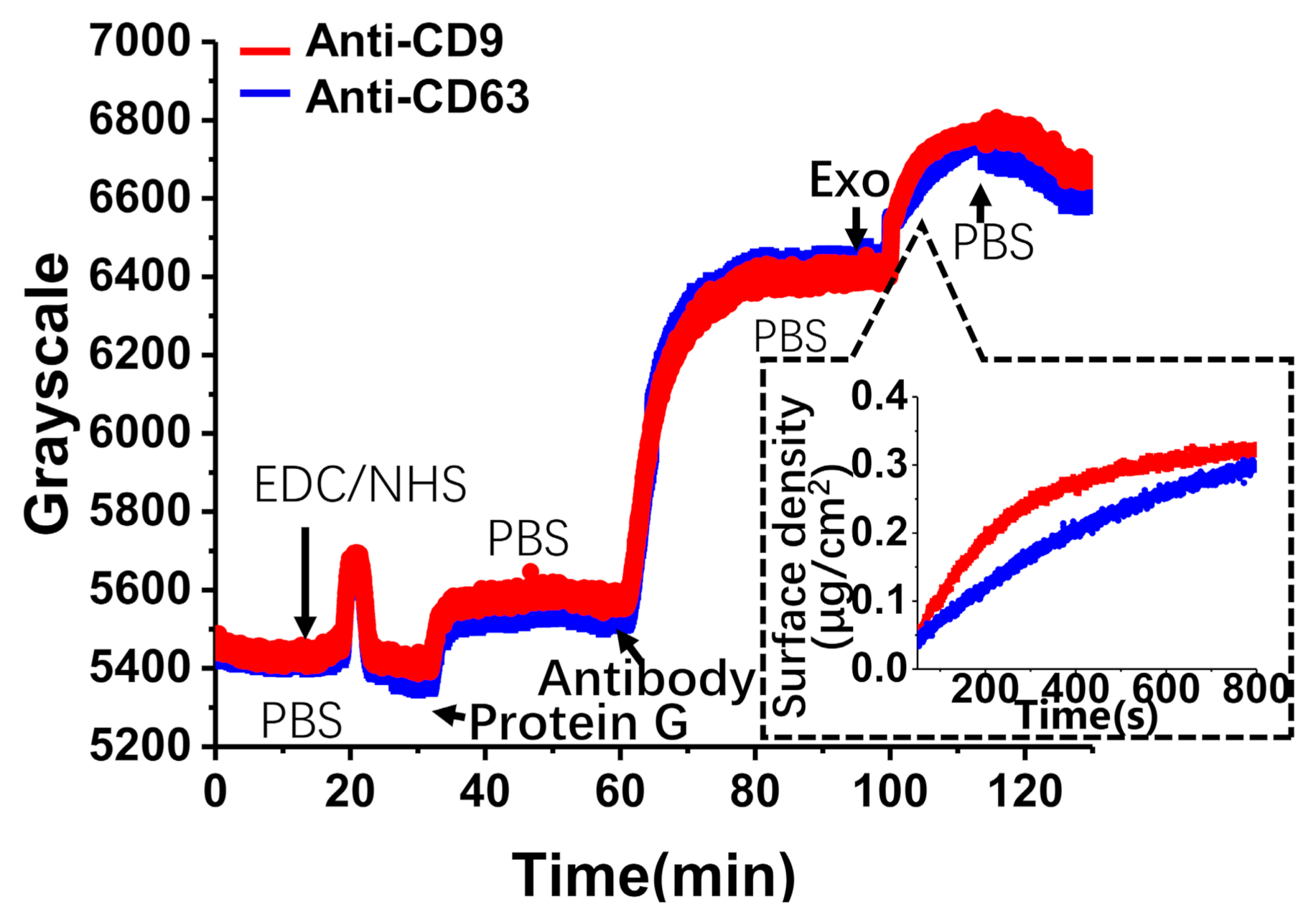
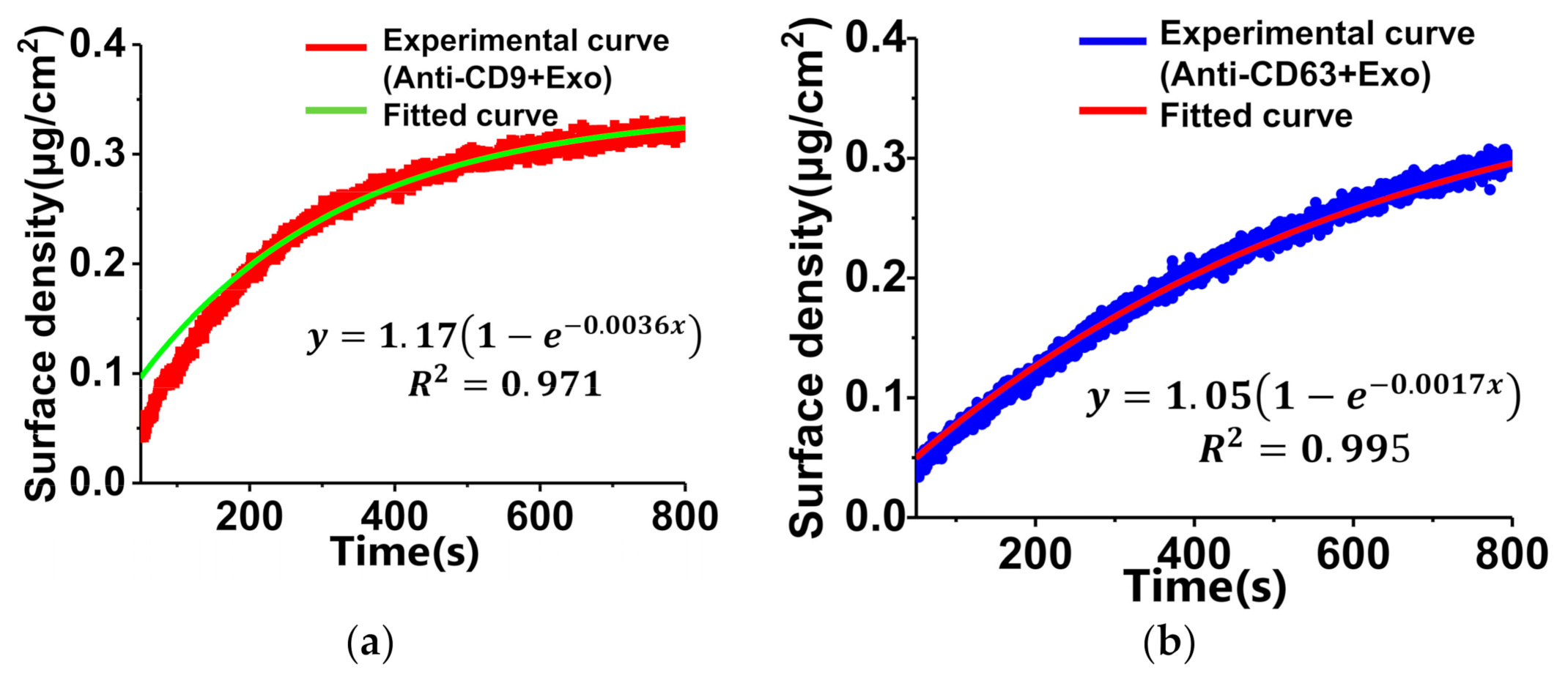
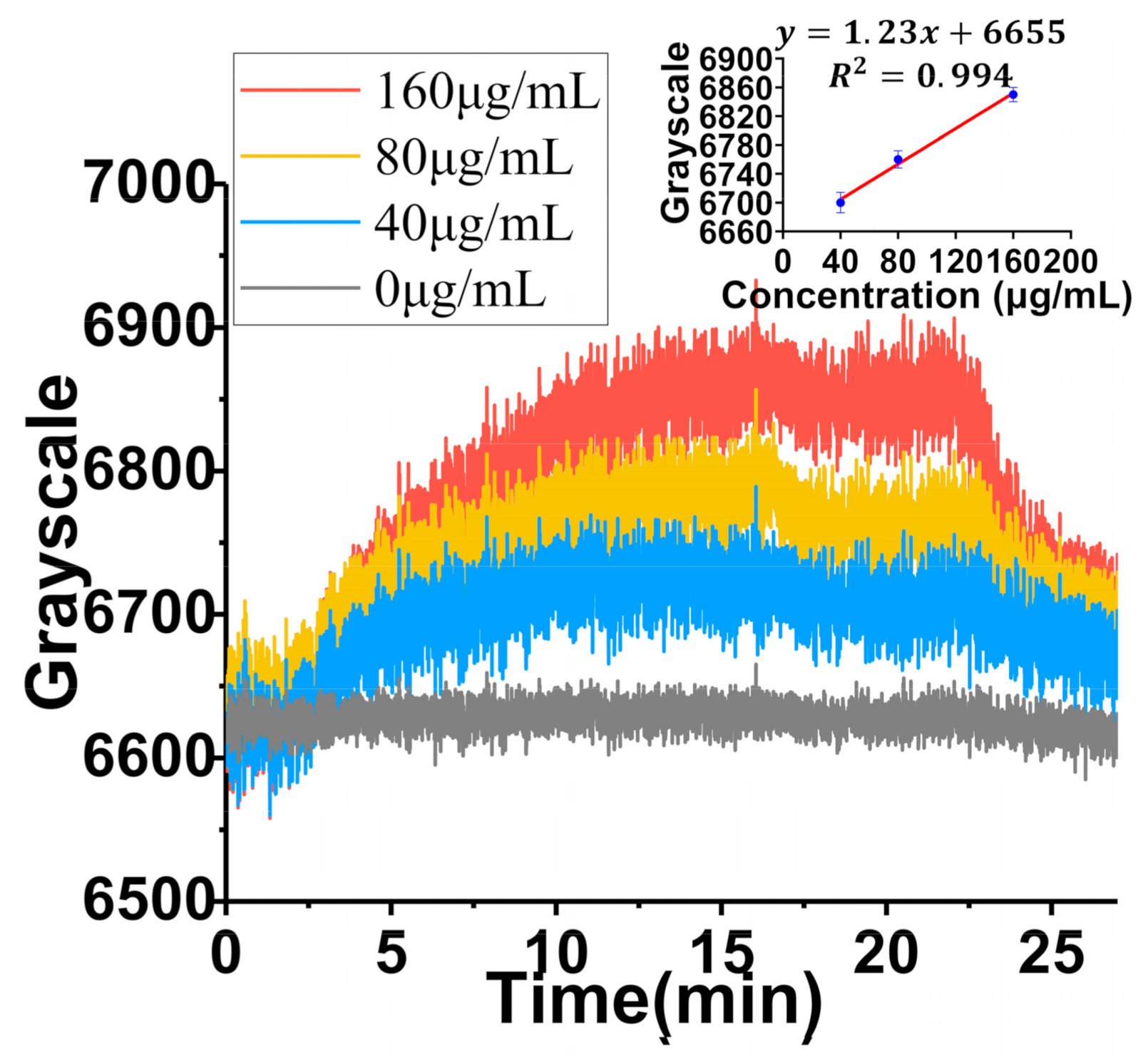
3.3. Detection and Analysis of Exosomes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Thery, C. Exosomes: Secreted vesicles and intercellular communications. F1000 Biol. Rep. 2011, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkach, M.; Thery, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Lotvall, J.; Hill, A.F.; Hochberg, F.; Buzas, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Beckett, K.; Monier, S.; Palmer, L.; Alexandre, C.; Green, H.; Bonneil, E.; Raposo, G.; Thibault, P.; Le Borgne, R.; Vincent, J.-P. Drosophila S2 Cells Secrete Wingless on Exosome-Like Vesicles but the Wingless Gradient Forms Independently of Exosomes. Traffic 2013, 14, 82–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt proteins are secreted on exosomes. Nat. Cell Biol. 2012, 14, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Vyas, N.; Walvekar, A.; Tate, D.; Lakshmanan, V.; Bansal, D.; Lo Cicero, A.; Raposo, G.; Palakodeti, D.; Dhawan, J. Vertebrate Hedgehog is secreted on two types of extracellular vesicles with different signaling properties. Sci. Rep. 2014, 4, 7357. [Google Scholar] [CrossRef] [PubMed]
- Al-Nedawi, K.; Meehan, B.; Kerbel, R.S.; Allison, A.C.; Rak, J. Endothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFR. Proc. Natl. Acad. Sci. USA 2009, 106, 3794–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doldan, X.; Fagundez, P.; Cayota, A.; Laiz, J.; Tosar, J.P. Electrochemical Sandwich Immunosensor for Determination of Exosomes Based on Surface Marker-Mediated Signal Amplification. Anal. Chem. 2016, 88, 10466–10473. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, V.; Ludwig, A.-K.; Hornung, S.; Rotan, O.; Horn, P.A.; Epple, M.; Glebel, B. Characterisation of exosomes derived from human cells by nanoparticle tracking analysis and scanning electron microscopy. Colloids Surf. B Biointerfaces 2011, 87, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Rupert, D.L.M.; Lasser, C.; Eldh, M.; Block, S.; Zhdanov, V.P.; Lotvall, J.O.; Bally, M.; Hook, F. Determination of Exosome Concentration in Solution Using Surface Plasmon Resonance Spectroscopy. Anal. Chem. 2014, 86, 5929–5936. [Google Scholar] [CrossRef] [PubMed]
- Marti, G.E.; Stetler-Stevenson, M.; Bleesing, J.J.; Fleisher, T.A. Introduction to flow cytometry. Semin. Hematol. 2001, 38, 93–99. [Google Scholar] [CrossRef]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanchetta, G.; Lanfranco, R.; Giavazzi, F.; Bellini, T.; Buscaglia, M. Emerging applications of label-free optical biosensors. Nanophotonics 2017, 6, 627–645. [Google Scholar] [CrossRef]
- Avella-Oliver, M.; Puchades, R.; Wachsmann-Hogiu, S.; Maquieira, A. Label-free SERS analysis of proteins and exosomes with large-scale substrates from recordable compact disks. Sens. Actuators B Chem. 2017, 252, 657–662. [Google Scholar] [CrossRef]
- Su, J. Label-Free Single Exosome Detection Using Frequency-Locked Microtoroid Optical Resonators. Acs Photonics 2015, 2, 1241–1245. [Google Scholar] [CrossRef]
- Aygun, U.; Ozkumur, A.Y.; Durmus, N.G.; Demirci, U.; Urey, H. Label-free imaging of exosomes using depth scanning correlation (DSC) interferometric microscopy. In Label-Free Biomedical Imaging and Sensing; Shaked, N.T., Hayden, O., Eds.; International Society for Optics and Photonics: Bellingham, WA, USA, 2020; Volume 11251. [Google Scholar] [CrossRef]
- Amano, R.; Takada, K.; Tanaka, Y.; Nakamura, Y.; Kawai, G.; Kozu, T.; Sakamoto, T. Kinetic and Thermodynamic Analyses of Interaction between a High Affinity RNA Aptamer and Its Target Protein. Biochemistry 2016, 55, 6221–6229. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Hsu, H.-Y.; Huang, C.-J.C. A protein detection technique by using surface plasmon resonance (SPR) with rolling circle amplification (RCA) and nanogold-modified tags. Biosens. Bioelectron. 2007, 22, 980–985. [Google Scholar] [CrossRef]
- Soler, M.; Carmen Estevez, M.; de Lourdes Moreno, M.; Cebolla, A.; Lechuga, L.M. Label-free SPR detection of gluten peptides in urine for non-invasive celiac disease follow-up. Biosens. Bioelectron. 2016, 79, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Tamada, K.; Nakamura, F.; Ito, M.; Li, X.; Baba, A. SPR-based DNA detection with metal nanoparticles. Plasmonics 2007, 2, 185–191. [Google Scholar] [CrossRef]
- Thakur, A.; Qiu, G.; Siu-Pang, N.G.; Guan, J.; Yue, J.; Lee, Y.; Wu, C.-M.L. Direct detection of two different tumor-derived extracellular vesicles by SAM-AuNIs LSPR biosensor. Biosens. Bioelectron. 2017, 94, 400–407. [Google Scholar] [CrossRef]
- Kang, T.F.; Niu, Y.; Jin, G. Visualization of the interaction between tris and lysozyme with a biosensor based on total internal reflection imaging ellipsometry. Thin Solid Film. 2014, 571, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Jin, G. Development of biosensor based on imaging ellipsometry. Phys. Status Solidi A 2008, 205, 810–816. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Meng, Y.H.; Liu, L.; Niu, Y.; Chen, S.; Cai, Q.; Jiang, T.J. Development of biosensor based on imaging ellipsometry and biomedical applications. Thin Solid Film. 2011, 519, 2750–2757. [Google Scholar] [CrossRef] [Green Version]
- Nabok, A.V.; Tsargorodskaya, A.; Hassan, A.K.; Starodub, N.F. Total internal reflection ellipsometry and SPR detection of low molecular weight environmental toxins. Appl. Surf. Sci. 2005, 246, 381–386. [Google Scholar] [CrossRef]
- Yuan, W.; Ho, H.P.; Wu, S.Y.; Suen, Y.K.; Kong, S.K. Polarization-sensitive surface plasmon enhanced ellipsometry biosensor using the photoelastic modulation technique. Sens. Actuators A Phys. 2009, 151, 23–28. [Google Scholar] [CrossRef]
- Niu, Y.; Jin, G. Protein microarray biosensors based on imaging ellipsometry techniques and their applications. Protein Cell 2011, 2, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, M.; Lv, B.; Chen, Y.Y.; Ma, H.W.; Viana, A.S.; Correia, J.P.; Jin, G. An Imaging Ellipsometry Approach to Dissolved Oxygen Measurement on Surface Tethered Weak Polyelectrolyte Modified Electrode. J. Electrochem. Soc. 2016, 163, H286–H291. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Shen, J.; Liu, W.; Niu, Y.; Jin, G. Imaging ellipsometry biosensor: Basic theory, principles of operation, and applications. J. Vac. Sci. Technol. B 2020, 38. [Google Scholar] [CrossRef]
- Derrick, J.P.; Wigley, D.B. The 3rd igG-binding Domain From Streptococcal Protein G: An analysis by X-ray crystallography of the structure alone and in a complex with fab. J. Mol. Biol. 1994, 243, 906–918. [Google Scholar] [CrossRef]
- Mahshid, S.S.; Camire, S.; Ricci, F.; Vallee-Belisle, A. A Highly Selective Electrochemical DNA-Based Sensor That Employs Steric Hindrance Effects to Detect Proteins Directly in Whole Blood. J. Am. Chem. Soc. 2015, 137, 15596–15599. [Google Scholar] [CrossRef]
- Li, Y.K.; Liu, W.; Jin, G.; Niu, Y.; Chen, Y.P.; Xie, M.X. Label-Free Sandwich Imaging Ellipsometry Immunosensor for Serological Detection of Procalcitonin. Anal. Chem. 2018, 90, 8002–8010. [Google Scholar] [CrossRef] [Green Version]
- Vijayendran, R.A.; Ligler, F.S.; Leckband, D.E. A computational reaction-diffusion model for the analysis of transport-limited kinetics. Anal. Chem. 1999, 71, 5405–5412. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Liu, W.; Jin, G. Detection of Exosomes Using Total Internal Reflected Imaging Ellipsometry. Biosensors 2021, 11, 164. https://doi.org/10.3390/bios11050164
Liu H, Liu W, Jin G. Detection of Exosomes Using Total Internal Reflected Imaging Ellipsometry. Biosensors. 2021; 11(5):164. https://doi.org/10.3390/bios11050164
Chicago/Turabian StyleLiu, Haoyu, Wei Liu, and Gang Jin. 2021. "Detection of Exosomes Using Total Internal Reflected Imaging Ellipsometry" Biosensors 11, no. 5: 164. https://doi.org/10.3390/bios11050164
APA StyleLiu, H., Liu, W., & Jin, G. (2021). Detection of Exosomes Using Total Internal Reflected Imaging Ellipsometry. Biosensors, 11(5), 164. https://doi.org/10.3390/bios11050164