Advances in Aptamer-Based Biosensors and Cell-Internalizing SELEX Technology for Diagnostic and Therapeutic Application

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Aptamers against Pathogenic Microorganisms and Mammalian Cells
2.1. Aptamers against Pathogenic Bacteria
2.2. Aptamers against Pathogenic Viruses
2.3. Aptamers against Pathogenic Unicellular Parasites
2.4. Aptamers against Mammalian Cells
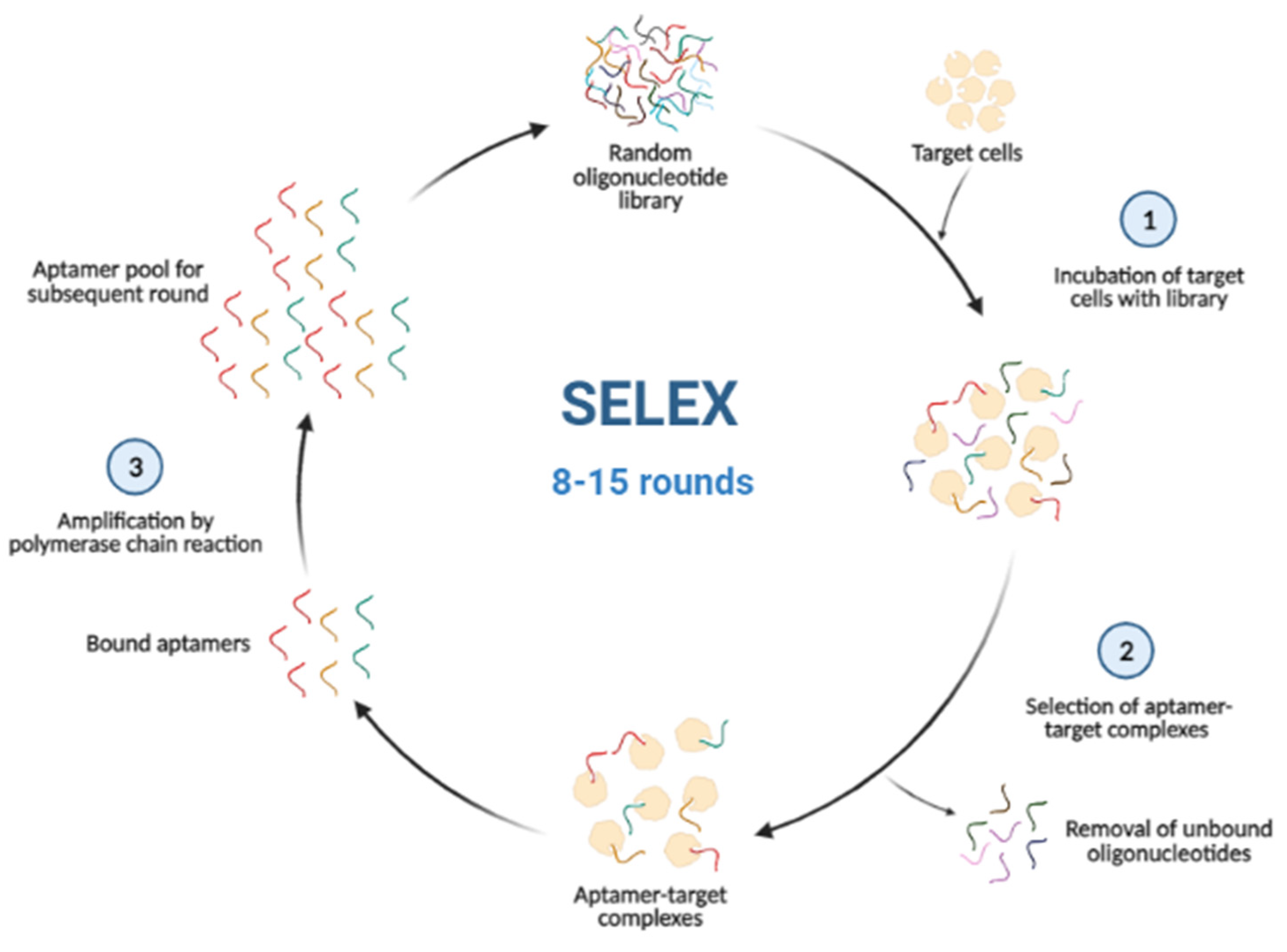
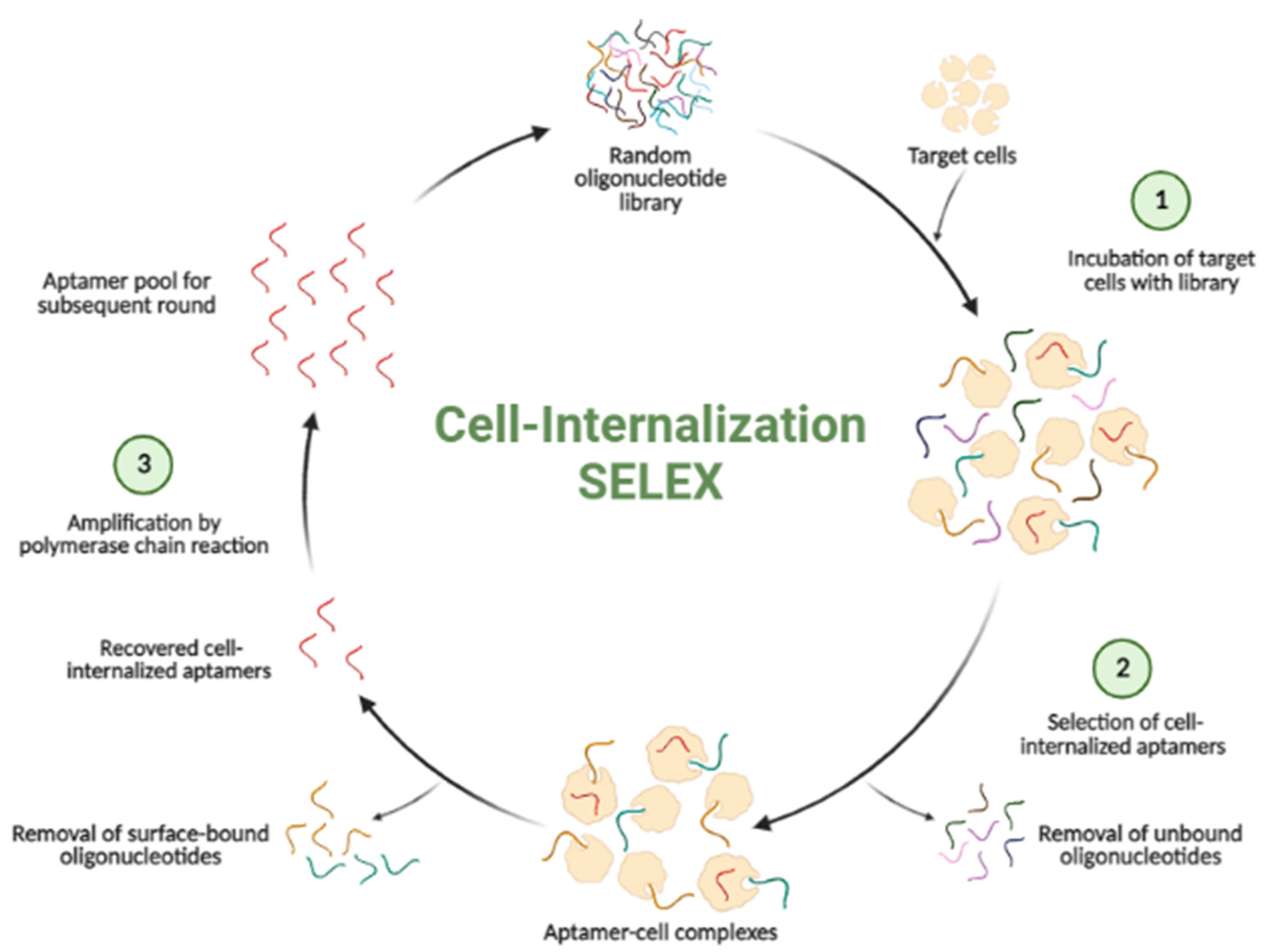
3. Cell-Internalization SELEX
3.1. Cell-Internalizing DNA Aptamers
3.2. Cell-Internalizing RNA Aptamers
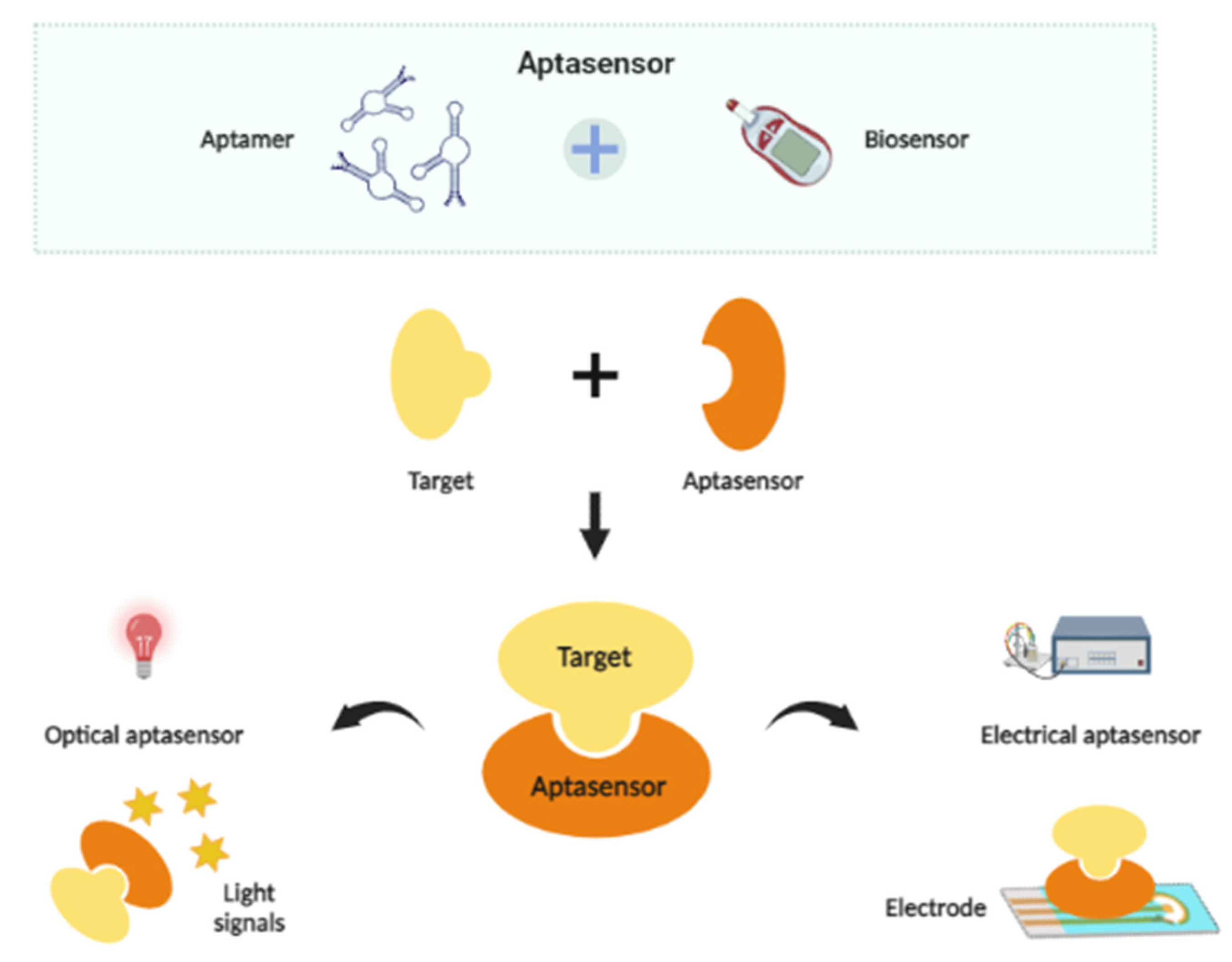
4. Aptamer-Based Biosensors
4.1. Optical Aptasensors for Microbial Detection
4.2. Electrical Aptasensors for Microbial Detection
5. Applications and Limitations of Aptamers
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ohuchi, S. Cell-Selex technology. BioRes. Open Access 2012, 1, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Xiong, H.; Cai, S.; Wen, N.; He, Q.; Liu, Y.; Peng, D.; Liu, Z. Advances in aptamer screening technologies. Talanta 2019, 200, 124–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Jiang, Y.; Hao, S.H.; Yan, X.Y.; Hong, D.F.; Naranmandura, H. Aptamers: An emerging navigation tool of therapeutic agents for targeted cancer therapy. J. Mater. Chem. B 2022, 10, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rossi, J. Aptamers as targeted therapeutics: Current potential and challenges. Nat. Rev. Drug Discov. 2017, 16, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Liu, M.; Tantai, W.; Xu, Q.; Zou, X.; Ma, F.; Zhang, C.Y. Aptamer-mediated rolling circle amplification for label-free and sensitive detection of histone acetyltransferase activity. Chem. Commun. 2021, 57, 2041–2044. [Google Scholar] [CrossRef]
- Yu, X.; Chen, F.; Wang, R.; Li, Y. Whole-bacterium SELEX of DNA aptamers for rapid detection of E. coli O157: H7 using a QCM sensor. J. Biotechnol. 2018, 266, 39–49. [Google Scholar] [CrossRef]
- Amraee, M.; Oloomi, M.; Yavari, A.; Bouzari, S. DNA aptamer identification and characterization for E. coli O157 detection using cell based SELEX method. Anal. Biochem. 2017, 536, 36–44. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Zhuo, Z.; Yu, Y.; Wang, M.; Li, J.; Zhang, Z.; Liu, J.; Wu, X.; Lu, A.; Zhang, G.; Zhang, B. Recent advances in SELEX technology and aptamer applications in biomedicine. Int. J. Mol. Sci. 2017, 18, 2142. [Google Scholar] [CrossRef]
- Davydova, A.; Vorobjeva, M.; Pyshnyi, D.; Altman, S.; Vlassov, V.; Venyaminova, A. Aptamers against pathogenic microorganisms. Crit. Rev. Microbiol. 2016, 42, 847–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Pandey, A. Whole cell SELEX technology and its applications. Aptamers Synth. Antibodies 2016, 2, 18–28. [Google Scholar]
- Zou, X.; Wu, J.; Gu, J.; Shen, L.; Mao, L. Application of aptamers in virus detection and antiviral therapy. Front. Microbiol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.Y.; Jia, W.N.; Li, X.L.; Zhang, L.; Liu, C.; Wu, J. Advances in detection of infectious agents by aptamer-based technologies. Emerg. Microbes Infect. 2020, 9, 1671–1681. [Google Scholar] [CrossRef]
- Alamudi, S.H.; Kimoto, M.; Hirao, I. Uptake mechanisms of cell-internalizing nucleic acid aptamers for applications as pharmacological agents. RSC Med. Chem. 2021, 12, 1640–1649. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, M.; Wei, S.; Qin, X.; Qin, Q.; Li, P. Research progress and prospects for the use of aptamers in aquaculture biosecurity. Aquaculture 2021, 534, 736257. [Google Scholar] [CrossRef]
- Duan, N.; Wu, S.; Chen, X.; Huang, Y.; Xia, Y.; Ma, X.; Wang, Z. Selection and characterization of aptamers against Salmonella typhimurium using whole-bacterium systemic evolution of ligands by exponential enrichment (SELEX). J. Agric. Food Chem. 2013, 61, 3229–3234. [Google Scholar] [CrossRef]
- Park, H.C.; Baig, I.A.; Lee, S.C.; Moon, J.Y.; Yoon, M.Y. Development of ssDNA aptamers for the sensitive detection of Salmonella typhimurium and Salmonella enteritidis. Appl. Biochem. Biotechnol. 2014, 174, 793–802. [Google Scholar] [CrossRef]
- Smith, I. Mycobacterium tuberculosis pathogenesis and molecular determinants of virulence. Clin. Microbiol. Rev. 2003, 16, 463–496. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhou, J.; Huang, Y.H.; Huang, F.Y.; Liu, Q.; Fang, Z.; Yang, S.; Xiong, M.; Lin, Y.Y.; Tan, G.H. Function of ssDNA aptamer and aptamer pool against Mycobacterium tuberculosis in a mouse model. Mol. Med. Rep. 2013, 7, 669–673. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhou, J.; Luo, F.; Mohammed, A.B.; Zhang, X.L. Aptamer from whole-bacterium SELEX as new therapeutic reagent against virulent Mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2007, 357, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Z.; Jiang, F.; Fu, P.; Shen, J.; Wu, W.; Li, J. Two DNA aptamers against avian influenza H9N2 virus prevent viral infection in cells. PLoS ONE 2015, 10, e0123060. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.M.; Lee, K.H.; Han, B.W.; Han, M.R.; Kim, D.H.; Kim, D.E. An RNA aptamer that specifically binds to the glycosylated hemagglutinin of avian influenza virus and suppresses viral infection in cells. PLoS ONE 2014, 9, e97574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombelli, S.; Minunni, M.; Luzi, E.; Mascini, M. Aptamer-based biosensors for the detection of HIV-1 Tat protein. Bioelectrochemistry 2005, 67, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Mufhandu, H.T.; Gray, E.S.; Madiga, M.C.; Tumba, N.; Alexandre, K.B.; Khoza, T.; Wibmer, C.K.; Moore, P.L.; Morris, L.; Khati, M. UCLA1, a synthetic derivative of a gp120 RNA aptamer, inhibits entry of human immunodeficiency virus type 1 subtype C. J. Virol. 2012, 86, 4989–4999. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Shibata, T.; Kabashima, T.; Kai, M. Inhibition of HIV-1 protease expression in T cells owing to DNA aptamer-mediated specific delivery of siRNA. Eur. J. Med. Chem. 2012, 56, 396–399. [Google Scholar] [CrossRef]
- Valero, J.; Civit, L.; Dupont, D.M.; Selnihhin, D.; Reinert, L.S.; Idorn, M.; Israels, B.A.; Bednarz, A.M.; Bus, C.; Asbach, B.; et al. A serum-stable RNA aptamer specific for SARS-CoV-2 neutralizes viral entry. Proc. Natl. Acad. Sci. USA 2021, 118, e2112942118. [Google Scholar] [CrossRef]
- Yang, G.; Li, Z.; Mohammed, I.; Zhao, L.; Wei, W.; Xiao, H.; Guo, W.; Zhao, Y.; Qu, F.; Huang, Y. Identification of SARS-CoV-2-against aptamer with high neutralization activity by blocking the RBD domain of spike protein 1. Signal Transduct. Target. Ther. 2021, 6, 227. [Google Scholar] [CrossRef]
- Hwang, S.D.; Midorikawa, N.; Punnarak, P.; Kikuchi, Y.; Kondo, H.; Hirono, I.; Aoki, T. Inhibition of hirame rhabdovirus growth by RNA aptamers. J. Fish Dis. 2012, 35, 927–934. [Google Scholar] [CrossRef]
- Li, P.; Yan, Y.; Wei, S.; Wei, J.; Gao, R.; Huang, X.; Huang, Y.; Jiang, G.; Qin, Q. Isolation and characterization of a new class of DNA aptamers specific binding to Singapore grouper iridovirus (SGIV) with antiviral activities. Virus Res. 2014, 188, 146–154. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, S.; Yu, Q.; Wei, S.; Liu, M.; Wei, J.; Huang, Y.; Huang, X.; Li, P.; Qin, Q. Characterization of novel aptamers specifically directed to red-spotted grouper nervous necrosis virus (RGNNV)-infected cells for mediating targeted siRNA delivery. Front. Microbiol. 2020, 11, 660. [Google Scholar] [CrossRef]
- Ulrich, H.; Magdesian, M.H.; Alves, M.J.M.; Colli, W. In vitro selection of RNA aptamers that bind to cell adhesion receptors of Trypanosoma cruzi and inhibit cell invasion. J. Biol. Chem. 2002, 277, 20756–20762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barfod, A.; Persson, T.; Lindh, J. In vitro selection of RNA aptamers against a conserved region of the Plasmodium falciparum erythrocyte membrane protein 1. Parasitol. Res. 2009, 105, 1557–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Duan, N.; Wu, S.; Shen, M.; Wang, Z. Selection, identification, and binding mechanism studies of an ssDNA aptamer targeted to different stages of E. coli O157: H7. J. Agric. Food Chem. 2018, 66, 5677–5682. [Google Scholar] [CrossRef] [PubMed]
- Rotherham, L.S.; Maserumule, C.; Dheda, K.; Theron, J.; Khati, M. Selection and application of ssDNA aptamers to detect active TB from sputum samples. PLoS ONE 2012, 7, e46862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Kim, G.; Park, S. Development of ssDNA aptamers for the capture and detection of Salmonella typhimurium. Anal. Methods 2014, 6, 7442–7448. [Google Scholar] [CrossRef]
- Kolovskaya, O.S.; Savitskaya, A.G.; Zamay, T.N.; Reshetneva, I.T.; Zamay, G.S.; Erkaev, E.N.; Wang, X.; Wehbe, M.; Salmina, A.B.; Perianova, O.V.; et al. Development of bacteriostatic DNA aptamers for Salmonella. J. Med. Chem. 2013, 56, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Yang, C.Y.; Sun, R.L.; Cheng, Y.F.; Kao, W.C.; Yang, P.C. Rapid single cell detection of Staphylococcus aureus by aptamer-conjugated gold nanoparticles. Sci. Rep. 2013, 3, 1863. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Liu, M.; Su, H.; Xiao, H.; Wu, S.; Qin, X.; Li, S.; Mi, H.; Lu, Z.; Shi, D.; et al. Selection and characterization of ssDNA aptamers specifically recognizing pathogenic Vibrio alginolyticus. J. Fish Dis. 2019, 42, 851–858. [Google Scholar] [CrossRef]
- Tang, X.; Zheng, J.; Yan, Q.; Li, Z.; Li, Y. Selection of aptamers against inactive Vibrio alginolyticus and application in a qualitative detection assay. Biotechnol. Lett. 2013, 35, 909–914. [Google Scholar] [CrossRef]
- Lai, H.C.; Wang, C.H.; Liou, T.M.; Lee, G.B. Influenza A virus-specific aptamers screened by using an integrated microfluidic system. Lab Chip 2014, 14, 2002–2013. [Google Scholar] [CrossRef] [PubMed]
- Ramos, E.; Moreno, M.; Martin, M.E.; Soto, M.; Gonzalez, V.M. In vitro selection of Leishmania infantum H3-binding ssDNA aptamers. Oligonucleotides 2010, 20, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Y.W.; Kwok, J.; Law, A.W.; Watt, R.M.; Kotaka, M.; Tanner, J.A. Structural basis for discriminatory recognition of Plasmodium lactate dehydrogenase by a DNA aptamer. Proc. Natl. Acad. Sci. USA 2013, 110, 15967–15972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homann, M.; Lorger, M.; Engstler, M.; Zacharias, M.; Ulrich Goringer, H. Serum-stable RNA aptamers to an invariant surface domain of live African trypanosomes. Comb. Chem. High Throughput Screen. 2006, 9, 491–499. [Google Scholar] [PubMed]
- Tan, W.; Wang, H.; Chen, Y.; Zhang, X.; Zhu, H.; Yang, C.; Yang, R.; Liu, C. Molecular aptamers for drug delivery. Trends Biotechnol. 2011, 29, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Xiang, J. Aptamers, the nucleic acid antibodies, in cancer therapy. Int. J. Mol. Sci. 2020, 21, 2793. [Google Scholar] [CrossRef]
- Daniels, D.A.; Chen, H.; Hicke, B.J.; Swiderek, K.M.; Gold, L. A tenascin-C aptamer identified by tumor cell SELEX: Systematic evolution of ligands by exponential enrichment. Proc. Natl. Acad. Sci. USA 2003, 100, 15416–15421. [Google Scholar] [CrossRef] [Green Version]
- Shangguan, D.; Li, Y.; Tang, Z.; Cao, Z.C.; Chen, H.W.; Mallikaratchy, P.; Sefah, K.; Yang, C.J.; Tan, W. Aptamers evolved from live cells as effective molecular probes for cancer study. Proc. Natl. Acad. Sci. USA 2006, 103, 11838–11843. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Kim, D.M.; Kim, K.S.; Jung, W.; Kim, D.E. Applications of cancer cell-specific aptamers in targeted delivery of anticancer therapeutic agents. Molecules 2018, 23, 830. [Google Scholar] [CrossRef] [Green Version]
- Mahlknecht, G.; Maron, R.; Mancini, M.; Schechter, B.; Sela, M.; Yarden, Y. Aptamer to ErbB-2/HER2 enhances degradation of the target and inhibits tumorigenic growth. Proc. Natl. Acad. Sci. USA 2013, 110, 8170–8175. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Parekh, P.; Turner, P.; Moyer, R.W.; Tan, W. Generating aptamers for recognition of virus-infected cells. Clin. Chem. 2009, 55, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Iwagawa, T.; Ohuchi, S.P.; Watanabe, S.; Nakamura, Y. Selection of RNA aptamers against mouse embryonic stem cells. Biochimie 2012, 94, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Han, D.; Ocsoy, I.; Tan, W. Aptamers selected by cell-SELEX for application in cancer studies. Bioanalysis 2010, 2, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liang, J.; Ma, Y.; Sun, B.; Li, X.; Wei, Y.; Fu, G.; Li, M.; Xia, W. Identification of a novel molecular probe for recognition of human osteosarcoma cell using the cell-SELEX method. Int. J. Clin. Exp. Med. 2015, 8, 18151. [Google Scholar] [PubMed]
- Yoon, S.; Rossi, J.J. Aptamers: Uptake mechanisms and intracellular applications. Adv. Drug Deliv. Rev. 2018, 134, 22–35. [Google Scholar] [CrossRef]
- Wu, C.C.; Castro, J.E.; Motta, M.; Cottam, H.B.; Kyburz, D.; Kipps, T.J.; Corr, M.; Carson, D.A. Selection of oligonucleotide aptamers with enhanced uptake and activation of human leukemia B cells. Hum. Gene Ther. 2003, 14, 849–860. [Google Scholar] [CrossRef]
- Ranches, G.; Lukasser, M.; Schramek, H.; Ploner, A.; Stasyk, T.; Mayer, G.; Mayer, G.; Hüttenhofer, A. In vitro selection of cell-internalizing DNA aptamers in a model system of inflammatory kidney disease. Mol. Ther. -Nucleic Acids 2017, 8, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Okuda, T.; Kasahara, Y.; Obika, S. Base-modified aptamers obtained by cell-internalization SELEX facilitate cellular uptake of an antisense oligonucleotide. Mol. Ther. -Nucleic Acids 2021, 23, 440–449. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Hernandez, F.J.; Gu, B.; Stockdale, K.R.; Nanapaneni, K.; Scheetz, T.E.; Behlke, M.A.; Peek, A.S.; Bair, T.; Giangrande, P.H.; et al. RNA aptamer-based functional ligands of the neurotrophin receptor, TrkB. Mol. Pharmacol. 2012, 82, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Thiel, W.H.; Bair, T.; Peek, A.S.; Liu, X.; Dassie, J.; Stockdale, K.R.; Behlke, M.A.; Miller Jr, F.J.; Giangrande, P.H. Rapid identification of cell-specific, internalizing RNA aptamers with bioinformatics analyses of a cell-based aptamer selection. PLoS ONE 2012, 7, e43836. [Google Scholar] [CrossRef] [Green Version]
- Thiel, W.H.; Thiel, K.W.; Flenker, K.S.; Bair, T.; Dupuy, A.J.; McNamara, J.O.; Miller, F.J.; Giangrande, P.H. Cell-internalization SELEX: Method for identifying cell-internalizing RNA aptamers for delivering siRNAs to target cells. In RNA Interference; Humana Press: New York, NY, USA, 2015; pp. 187–199. [Google Scholar]
- Thiel, K.W.; Hernandez, L.I.; Dassie, J.P.; Thiel, W.H.; Liu, X.; Stockdale, K.R.; Rothman, A.M.; Hernandez, F.J.; McNamara, J.O.; Giangrande, P.H. Delivery of chemo-sensitizing siRNAs to HER2+-breast cancer cells using RNA aptamers. Nucleic Acids Res. 2012, 40, 6319–6337. [Google Scholar] [CrossRef] [PubMed]
- Paniel, N.; Baudart, J.; Hayat, A.; Barthelmebs, L. Aptasensor and genosensor methods for detection of microbes in real world samples. Methods 2013, 64, 229–240. [Google Scholar] [PubMed]
- Hong, P.; Li, W.; Li, J. Applications of aptasensors in clinical diagnostics. Sensors 2012, 12, 1181–1193. [Google Scholar] [PubMed]
- Han, K.; Liang, Z.; Zhou, N. Design strategies for aptamer-based biosensors. Sensors 2010, 10, 4541–4557. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, A.S.; Ansari, N.; Ramezani, M.; Abnous, K.; Mohsenzadeh, M.; Taghdisi, S.M.; Alibolandi, M. Optical and electrochemical aptasensors for the detection of amphenicols. Biosens. Bioelectron. 2018, 118, 137–152. [Google Scholar] [CrossRef]
- Pashazadeh, P.; Mokhtarzadeh, A.; Hasanzadeh, M.; Hejazi, M.; Hashemi, M.; de la Guardia, M. Nano-materials for use in sensing of salmonella infections: Recent advances. Biosens. Bioelectron. 2017, 87, 1050–1064. [Google Scholar] [CrossRef]
- Zahra, Q.; Khan, Q.A.; Luo, Z. Advances in optical aptasensors for early detection and diagnosis of various cancer types. Front. Oncol. 2021, 11, 632165. [Google Scholar] [CrossRef]
- Musumeci, D.; Platella, C.; Riccardi, C.; Moccia, F.; Montesarchio, D. Fluorescence sensing using DNA aptamers in cancer research and clinical diagnostics. Cancers 2017, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, H.P.; Smiley, R.D.; Jaykus, L.A. Selection and characterization of DNA aptamers with binding selectivity to Campylobacter jejuni using whole-cell SELEX. Appl. Microbiol. Biotechnol. 2010, 87, 2323–2334. [Google Scholar] [CrossRef]
- Ohk, S.H.; Koo, O.K.; Sen, T.; Yamamoto, C.M.; Bhunia, A.K. Antibody–aptamer functionalized fibre-optic biosensor for specific detection of Listeria monocytogenes from food. J. Appl. Microbiol. 2010, 109, 808–817. [Google Scholar] [CrossRef]
- Chung, J.; Kang, J.S.; Jurng, J.S.; Jung, J.H.; Kim, B.C. Fast and continuous microorganism detection using aptamer-conjugated fluorescent nanoparticles on an optofluidic platform. Biosens. Bioelectron. 2015, 67, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Zhong, Z.; Gao, X.; Jia, L. Graphene oxide quantum dots assisted construction of fluorescent aptasensor for rapid detection of Pseudomonas aeruginosa in food samples. J. Agric. Food Chem. 2018, 66, 10898–10905. [Google Scholar] [CrossRef]
- Kou, X.; Zhang, X.; Shao, X.; Jiang, C.; Ning, L. Recent advances in optical aptasensor technology for amplification strategies in cancer diagnostics. Anal. Bioanal. Chem. 2020, 412, 6691–6705. [Google Scholar] [CrossRef]
- Khang, J.; Kim, D.; Chung, K.W.; Lee, J.H. Chemiluminescent aptasensor capable of rapidly quantifying Escherichia coli O157: H7. Talanta 2016, 147, 177–183. [Google Scholar] [CrossRef]
- Akshaya, K.; Arthi, C.; Pavithra, A.J.; Poovizhi, P.; Antinate, S.S.; Hikku, G.S.; Jeyasubramanian, K.; Murugesan, R. Bioconjugated gold nanoparticles as an efficient colorimetric sensor for cancer diagnostics. Photodiagnosis Photodyn. Ther. 2020, 30, 101699. [Google Scholar] [CrossRef] [PubMed]
- Duan, N.; Xu, B.; Wu, S.; Wang, Z. Magnetic nanoparticles-based aptasensor using gold nanoparticles as colorimetric probes for the detection of Salmonella typhimurium. Anal. Sci. 2016, 32, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Deng, W.; Yang, L.; Tan, Y.; Xie, Q.; Yao, S. Copper-based metal–organic framework nanoparticles with peroxidase-like activity for sensitive colorimetric detection of Staphylococcus aureus. ACS Appl. Mater. Interfaces 2017, 9, 24440–24445. [Google Scholar] [CrossRef]
- Feng, J.; Shen, Q.; Wu, J.; Dai, Z.; Wang, Y. Naked-eyes detection of Shigella flexneri in food samples based on a novel gold nanoparticle-based colorimetric aptasensor. Food Control. 2019, 98, 333–341. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Park, J.; Kang, S.; Kim, M. Surface plasmon resonance: A versatile technique for biosensor applications. Sensors 2015, 15, 10481–10510. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Wang, R.; Hargis, B.; Lu, H.; Li, Y. A SPR aptasensor for detection of avian influenza virus H5N1. Sensors 2012, 12, 12506–12518. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ye, Z.; Si, C.; Ying, Y. Application of aptamer based biosensors for detection of pathogenic microorganisms. Chin. J. Anal. Chem. 2012, 40, 634–642. [Google Scholar] [CrossRef]
- Ma, X.; Xu, X.; Xia, Y.; Wang, Z. SERS aptasensor for Salmonella typhimurium detection based on spiny gold nanoparticles. Food Control. 2018, 84, 232–237. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, X.; Liu, Y.; Duan, N.; Wu, S.; Wang, Z.; Xu, B. Gold nanoparticles enhanced SERS aptasensor for the simultaneous detection of Salmonella typhimurium and Staphylococcus aureus. Biosens. Bioelectron. 2015, 74, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Xiao, W.; Li, G.; Wu, P.; He, Y.; Chen, C.; He, Y.; Ding, P.; Kai, T. The research of aptamer biosensor technologies for detection of microorganism. Appl. Microbiol. Biotechnol. 2020, 104, 9877–9890. [Google Scholar] [CrossRef]
- Abbaspour, A.; Norouz-Sarvestani, F.; Noori, A.; Soltani, N. Aptamer conjugated silver nanoparticles for electrochemical dual-aptamer-based sandwich detection of Staphylococcus aureus. Biosens. Bioelectron. 2015, 68, 149–155. [Google Scholar] [CrossRef]
- Diba, F.S.; Kim, S.; Lee, H.J. Amperometric bioaffinity sensing platform for avian influenza virus proteins with aptamer modified gold nanoparticles on carbon chips. Biosens. Bioelectron. 2015, 72, 355–361. [Google Scholar] [CrossRef]
- Zelada-Guillén, G.A.; Sebastián-Avila, J.L.; Blondeau, P.; Riu, J.; Rius, F.X. Label-free detection of Staphylococcus aureus in skin using real-time potentiometric biosensors based on carbon nanotubes and aptamers. Biosens. Bioelectron. 2012, 31, 226–232. [Google Scholar] [CrossRef]
- Zelada-Guillén, G.A.; Bhosale, S.V.; Riu, J.; Rius, F.X. Real-time potentiometric detection of bacteria in complex samples. Anal. Chem. 2010, 82, 9254–9260. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, Z.; Ying, Y. New trends in impedimetric biosensors for the detection of foodborne pathogenic bacteria. Sensors 2012, 12, 3449–3471. [Google Scholar] [CrossRef] [Green Version]
- Bagheryan, Z.; Raoof, J.B.; Golabi, M.; Turner, A.P.F.; Beni, V. Diazonium-based impedimetric aptasensor for the rapid label-free detection of Salmonella typhimurium in food sample. Biosens. Bioelectron. 2016, 80, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Shorie, M.; Sabherwal, P. Electrochemical aptasensor using boron-carbon nanorods decorated by nickel nanoparticles for detection of E. coli O157: H7. Microchim. Acta 2020, 187, 461. [Google Scholar] [CrossRef] [PubMed]
- Minunni, M.; Tombelli, S.; Gullotto, A.; Luzi, E.; Mascini, M. Development of biosensors with aptamers as bio-recognition element: The case of HIV-1 Tat protein. Biosens. Bioelectron. 2004, 20, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, Y. Hydrogel based QCM aptasensor for detection of avian influenzavirus. Biosens. Bioelectron. 2013, 42, 148–155. [Google Scholar] [CrossRef]
- Douglas, S.M.; Bachelet, I.; Church, G.M. A logic-gated nanorobot for targeted transport of molecular payloads. Science 2012, 335, 831–834. [Google Scholar] [CrossRef]
- Ayodele, O.O.; Adesina, A.O.; Pourianejad, S.; Averritt, J.; Ignatova, T. Recent advances in nanomaterial-based aptasensors in medical diagnosis and therapy. Nanomaterials 2021, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, S.H.; Kim, J.H.; Noh, Y.H.; Noh, G.J.; Lee, S.W. Pharmacokinetics of a cholesterol-conjugated aptamer against the hepatitis C virus (HCV) NS5B protein. Mol. Ther. -Nucleic Acids 2015, 4, e254. [Google Scholar] [CrossRef]
- Heo, K.; Min, S.W.; Sung, H.J.; Kim, H.G.; Kim, H.J.; Kim, Y.H.; Choi, B.K.; Han, S.; Chung, S.; Lee, E.S.; et al. An aptamer-antibody complex (oligobody) as a novel delivery platform for targeted cancer therapies. J. Control. Release 2016, 229, 1–9. [Google Scholar] [CrossRef]
- Blind, M.; Blank, M. Aptamer Selection Technology and Recent Advances. Mol. Ther. Nucleic Acids 2015, 4, e223. [Google Scholar] [CrossRef]
- Wang, R.E.; Wu, H.; Niu, Y.; Cai, J. Improving the stability of aptamers by chemical modification. Curr. Med. Chem. 2011, 18, 4126–4138. [Google Scholar] [CrossRef]
- Smirnov, I.; Shafer, R.H. Effect of loop sequence and size on DNA aptamer stability. Biochemistry 2000, 39, 1462–1468. [Google Scholar] [CrossRef]
- Darfeuille, F.; Hansen, J.B.; Orum, H.; Primo, C.D.; Toulmé, J. LNA/DNA chimeric oligomers mimic RNA aptamers targeted to the TAR RNA element of HIV-1. Nucleic Acids Res. 2004, 32, 3101–3107. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.D.; Cload, S.T. SELEX with modified nucleotides. Curr. Opin. Chem. Biol. 2008, 12, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Goringer, H.U.; Adler, A.; Forster, N.; Homann, M. Post-SELEX chemical optimization of a trypanosome-specific RNA aptamer. Comb. Chem. High Throughput Screen. 2008, 11, 16–23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Microorganisms | Nucleic Acid Pool | Dissociation Constant (Kd) | References |
|---|---|---|---|
| Bacteria | |||
| Escherichia coli O157:H7 | DNA | 10.30 nM | [6] |
| DNA | 107.6 ± 67.8 pM | [7] | |
| DNA | 9.04 ± 2.80 nM | [34] | |
| Mycobacterium tuberculosis H37Rv | DNA | - | [20,21] |
| Mycobacterium tuberculosis | DNA | Nanomolar (nM) range | [35] |
| Salmonella typhimurium | DNA | 6.33 ± 0.58 nM | [17] |
| DNA | 0.360 ± 0.103 nM | [36] | |
| Salmonella typhimurium, Salmonella enteritidis | DNA | Nanomolar (nM) to micromolar (µM) range | [18] |
| DNA | 25 nM, 7 nM | [37] | |
| Staphylococcus aureus | DNA | 35 nM, 129 nM | [38] |
| Vibrio alginolyticus | DNA | 14.31 ± 4.26 nM, 90.00 ± 13.51 nM | [39] |
| DNA | 27.5 ± 9.2 nM | [40] | |
| Viruses | |||
| Avian influenza H9N2 virus | DNA | Nanomolar (nM) range | [22] |
| Avian influenza H5N1 virus | RNA | - | [23] |
| Avian influenza H1N1 virus | DNA | 55.14 ± 22.40 nM | [41] |
| Hirame rhabdovirus (HIRRV) | RNA | - | [29] |
| Human immunodeficiency virus (HIV) | RNA | - | [24] |
| RNA | 0.15 nM | [25] | |
| DNA | - | [26] | |
| Red-spotted grouper nervous necrosis virus (RGNNV) | DNA | Nanomolar (nM) range | [31] |
| Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) | 2′-fPy-RNA | Picomolar (pM) range | [27] |
| DNA | 0.118 ± 0.033–85.610 ± 14.219 nM | [28] | |
| Singapore grouper iridovirus (SGIV) | DNA | - | [30] |
| Unicellular parasites | |||
| Leishmania infantum | DNA | 0.94 ± 0.19 nM | [42] |
| Plasmodium falciparum | 2′-fPy-RNA | - | [33] |
| DNA | 42 nM | [43] | |
| Trypanosoma brucei | 2′-NH2-RNA | 70 ± 15 nM | [44] |
| Trypanosoma cruzi | 2′-fPy-RNA | 40–400 nM | [32] |
| Detection Method | Target Microorganisms | Detection Range | Limit of Detection | Real Samples | References |
|---|---|---|---|---|---|
| Optical | |||||
| Fluorescence | Escherichia coli | ~100/s detection rate on single E. coli cells | - | - | [72] |
| Fluorescence | Pseudomonas aeruginosa | 1.28 × 103–2.00 × 107 cfu/mL | 100 cfu/mL | Drinking water, orange juice, popsicle | [73] |
| Chemiluminescence | Escherichia coli O157:H7 | 104–107 cfu/mL | 4.5 × 103 cfu/mL | Skim milk | [75] |
| Colorimetric | Salmonella typhimurium | 25–105 cfu/mL | 10 cfu/mL | Milk | [77] |
| Colorimetric | Shigella flexneri | 102–106 cfu/mL | 80 cfu/mL | Salmon | [79] |
| Colorimetric | Staphylococcus aureus | 50–104 cfu/mL | 20 cfu/mL | Milk | [78] |
| Surface plasmon resonance (SPR) | AIV H5N1 | 0.128–1.280 HAU | 0.128 HAU | Poultry | [81] |
| Surface-enhanced Raman scattering (SERS) | Salmonella typhimurium | 101–105 cfu/mL | 4 cfu/mL | Pork | [83] |
| Surface-enhanced Raman scattering (SERS) | Salmonella typhimurium, Staphylococcus aureus | 102–107 cfu/mL | 15 cfu/mL; 35 cfu/mL | Pork | [84] |
| Electrical | |||||
| Differential pulse voltammetry | Staphylococcus aureus | 10–1 × 106 cfu/mL | 1 cfu/mL | Tap and river water | [86] |
| Differential pulse voltammetry | AIV H5N1 | 100 fM–10 pM | 100 fM | Human serum | [87] |
| Potentiometry | Escherichia coli | 4.0–2.4 × 104 cfu/mL | 4.0 cfu/mL | Milk, apple juice | [89] |
| Potentiometry | Staphylococcus aureus | 8 × 102–108 cfu/mL;107–108 cfu/mL | 8 × 102 cfu/mL; 107 cfu/mL | Pig skin | [88] |
| Impedance | Escherichia coli O157:H7 | 100–105 cfu/mL | 10 cfu/mL | Tap water, juice, fecal | [92] |
| Impedance | Salmonella typhimurium | 1 × 101–1 × 108 cfu/mL | 6 cfu/mL | Apple juice | [91] |
| Piezoelectricity | AIV H5N1 | - | 0.0128 HAU | - | [94] |
| Piezoelectricity | Tat protein of HIV-1 virus | 0–1.25 mg/L | 0.25 mg/L | - | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, Z.; Roslan, M.A.M.; Abd Shukor, M.Y.; Halim, M.; Yasid, N.A.; Abdullah, J.; Md Yasin, I.S.; Wasoh, H. Advances in Aptamer-Based Biosensors and Cell-Internalizing SELEX Technology for Diagnostic and Therapeutic Application. Biosensors 2022, 12, 922. https://doi.org/10.3390/bios12110922
Gan Z, Roslan MAM, Abd Shukor MY, Halim M, Yasid NA, Abdullah J, Md Yasin IS, Wasoh H. Advances in Aptamer-Based Biosensors and Cell-Internalizing SELEX Technology for Diagnostic and Therapeutic Application. Biosensors. 2022; 12(11):922. https://doi.org/10.3390/bios12110922
Chicago/Turabian StyleGan, Zixuen, Muhamad Aidilfitri Mohamad Roslan, Mohd Yunus Abd Shukor, Murni Halim, Nur Adeela Yasid, Jaafar Abdullah, Ina Salwany Md Yasin, and Helmi Wasoh. 2022. "Advances in Aptamer-Based Biosensors and Cell-Internalizing SELEX Technology for Diagnostic and Therapeutic Application" Biosensors 12, no. 11: 922. https://doi.org/10.3390/bios12110922
APA StyleGan, Z., Roslan, M. A. M., Abd Shukor, M. Y., Halim, M., Yasid, N. A., Abdullah, J., Md Yasin, I. S., & Wasoh, H. (2022). Advances in Aptamer-Based Biosensors and Cell-Internalizing SELEX Technology for Diagnostic and Therapeutic Application. Biosensors, 12(11), 922. https://doi.org/10.3390/bios12110922