Europium Nanoparticle-Based Lateral Flow Strip Biosensors Combined with Recombinase Polymerase Amplification for Simultaneous Detection of Five Zoonotic Foodborne Pathogens

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture Preparation and DNA Extraction
2.2. Reagents and Apparatus
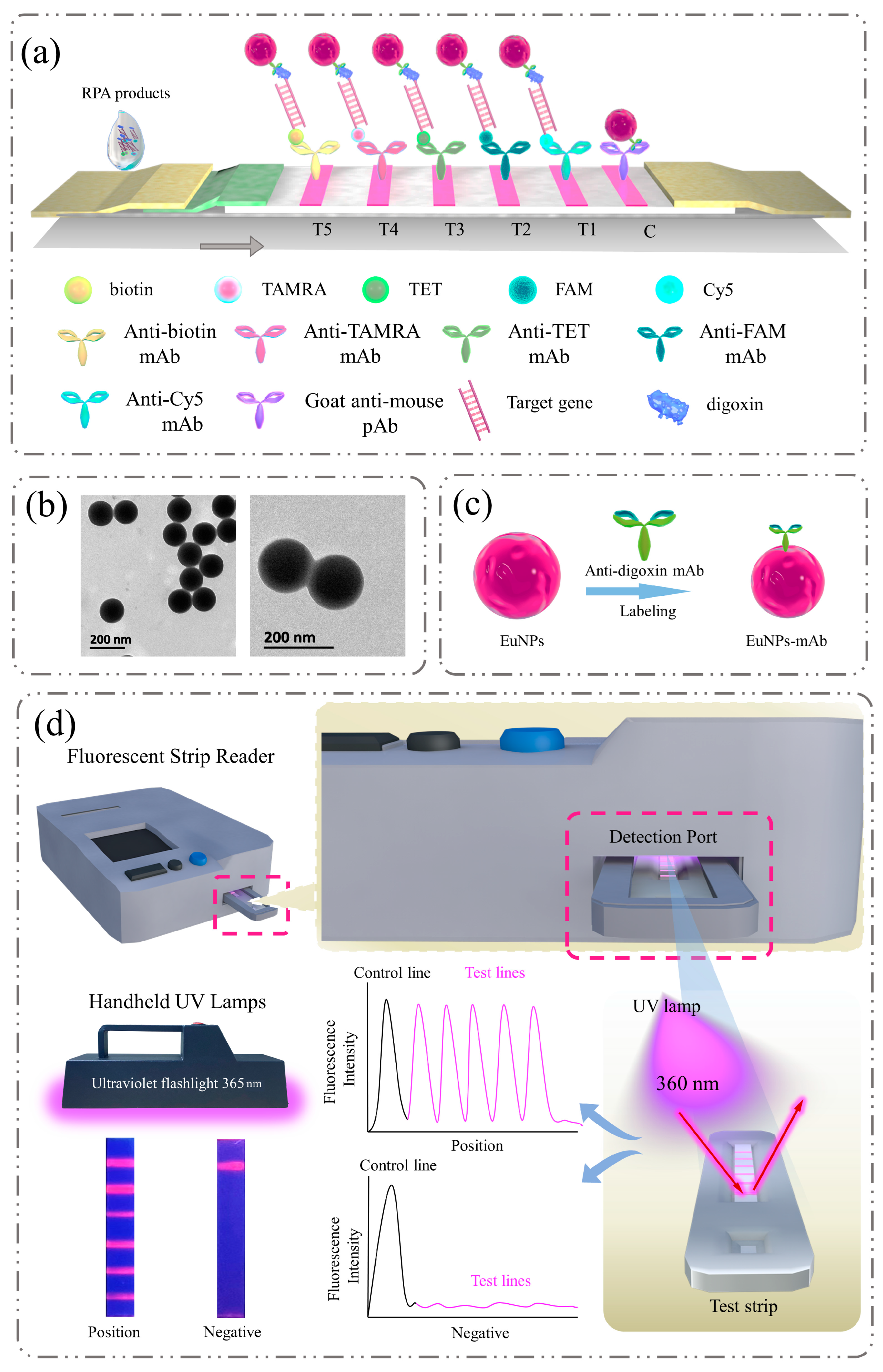
2.3. Preparation of EuNP-mAb
2.4. Primer Design and Assembly of Quintuple RPA-EuNP-LFSBs
2.5. Multiplex Reaction Protocols for RPA
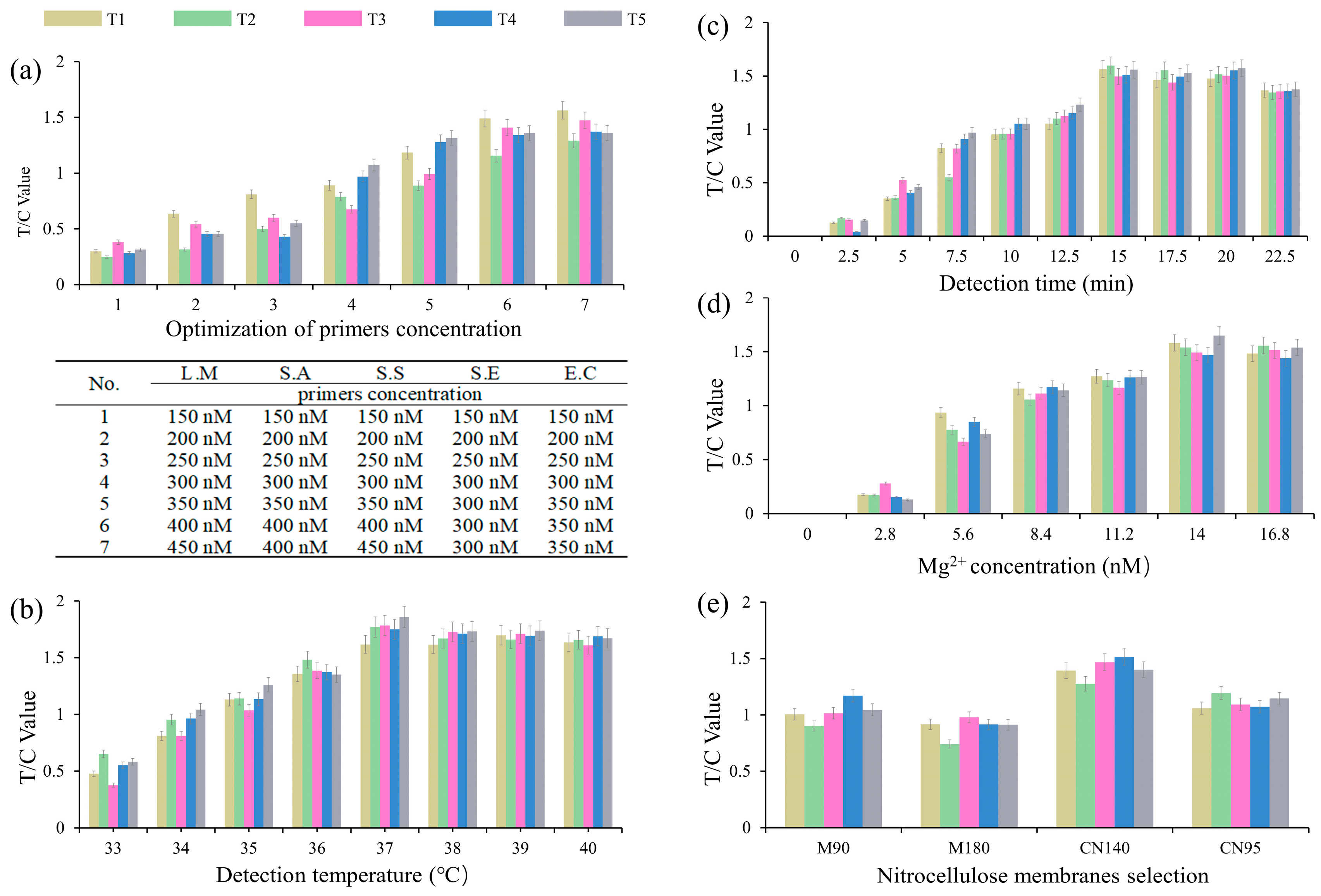
2.6. Optimization of Quintuple RPA-EuNP-LFSBs
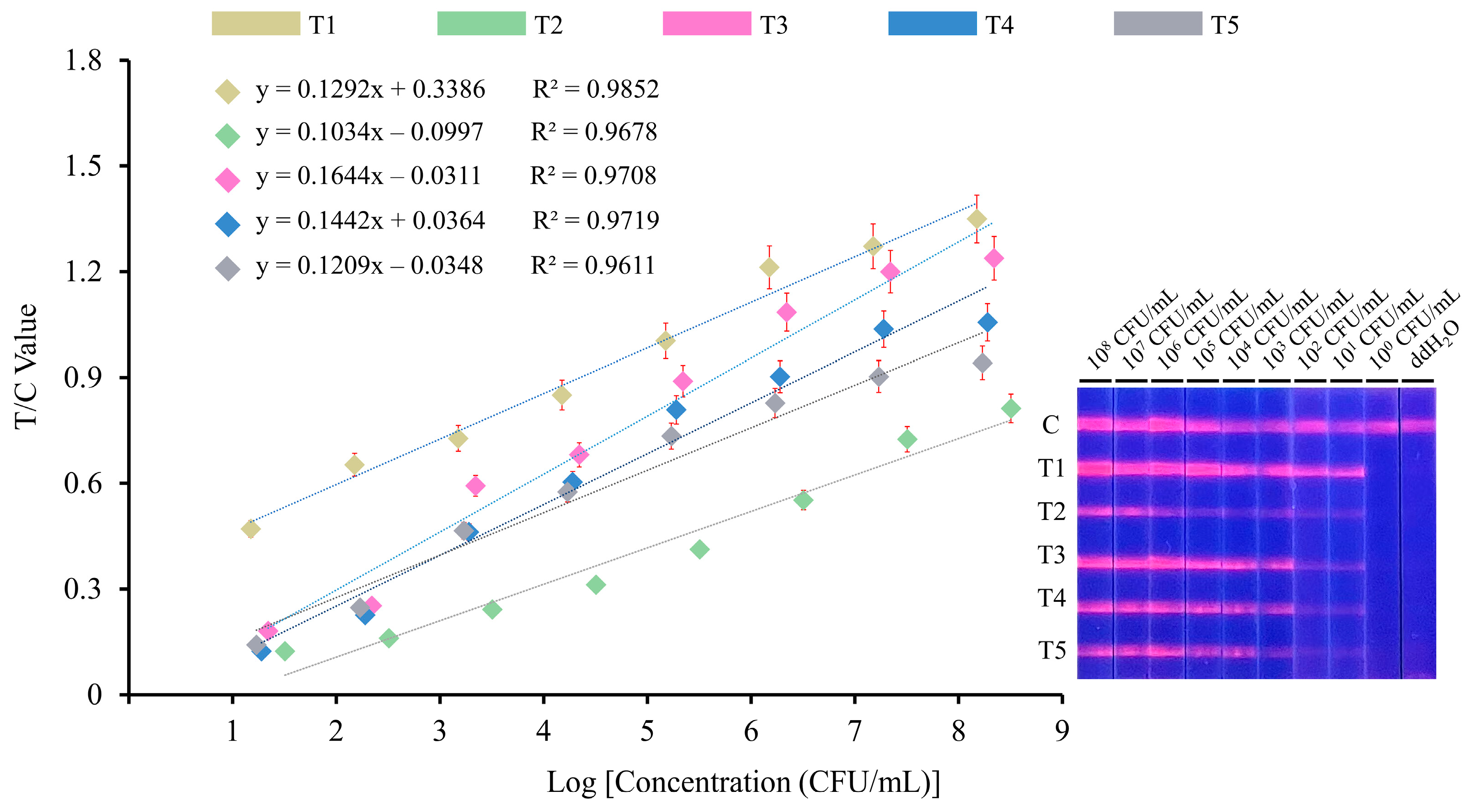
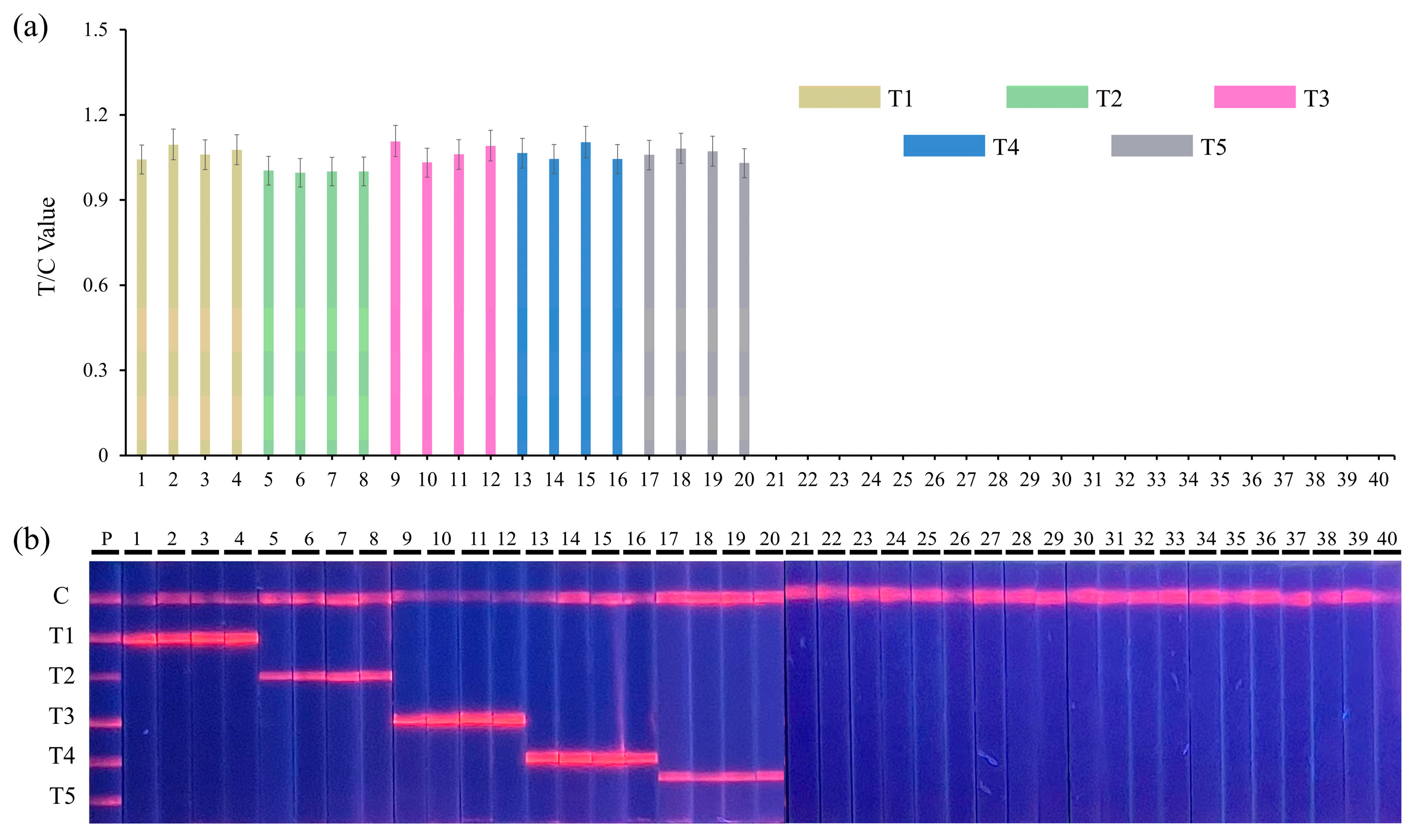
2.7. Sensitivity and Specificity
2.8. Artificially Contaminated Food Samples
2.9. Analysis of Quintuple RPA-EuNP-LFSBs in Field Samples
2.10. Statistical Analysis
3. Results and Discussion
3.1. Assay Principle
3.2. Optimization of the Quintuple RPA-EuNP-LFSBs
3.3. Sensitivity and Specificity
3.4. Detection of Quintuple RPA-EuNP-LFSBs in Artificially Contaminated Food
3.5. Detection of Quintuple RPA-EuNP-LF/SBs in Actual Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/zoonoses (accessed on 1 January 2023).
- Ali, S.; Alsayeqh, A.F. Review of major meat-borne zoonotic bacterial pathogens. Front. Public Health 2022, 10, 1045599. [Google Scholar] [CrossRef] [PubMed]
- Milbank, C.; Vira, B. Wildmeat consumption and zoonotic spillover: Contextualising disease emergence and policy responses. Lancet Planet. Health 2022, 6, e439–e448. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Campylobacter. Available online: https://www.who.int/publications/i/item/9789241565165 (accessed on 1 January 2023).
- Cascio, A.; Bosilkovski, M.; Rodriguez-Morales, A.J.; Pappas, G. The socio-ecology of zoonotic infections. Clin. Microbiol. Infect. 2011, 17, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Zelenik, K.; Avberšek, J.; Pate, M.; Lušicky, M.; Krt, B.; Ocepek, M.; Zdovc, I. Cutaneous listeriosis in a veterinarian with the evidence of zoonotic transmission—A case report. Zoonoses Public Health 2014, 61, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Ariza-Miguel, J.; Hernández, M.; Fernández-Natal, I.; Rodríguez-Lázaro, D. Methicillin-resistant Staphylococcus aureus harboring mecC in livestock in Spain. J. Clin. Microbiol. 2014, 52, 4067–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banky, J.P.; Ostergaard, L.; Spelman, D. Chronic relapsing Salmonella osteomyelitis in an immunocompetent patient: Case report and literature review. J. Infect. 2002, 44, 44–47. [Google Scholar] [CrossRef]
- Alhadlaq, M.A.; Mujallad, M.I.; Alajel, S.M.I. Detection of Escherichia coli O157:H7 in imported meat products from Saudi Arabian ports in 2017. Sci. Rep. 2023, 13, 4222. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China. Available online: https://www.moa.gov.cn/govpublic/xmsyj/202208/t20220819_6407309 (accessed on 11 March 2023).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, e07209. [Google Scholar]
- Sant’Ana, A.S.; Franco, B.; Schaffner, D.W. Risk of infection with Salmonella and Listeria monocytogenes due to consumption of ready-to-eat leafy vegetables in brazil. Food Control 2014, 42, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.; Sousa, S.; Cabanes, D. How listeria monocytogenes organizes its surface for virulence. Front. Cell. Infect. Microbiol. 2014, 4, 00048. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, B.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Aklilu, E.; Zunita, Z.; Hassan, L.; Chen, H. Phenotypic and genotypic characterization of methicillin-resistant Staphylococcus aureus (MRSA) isolated from dogs and cats at University Veterinary Hospital, Universiti Putra Malaysia. Trop Biomed. 2010, 27, 483–492. [Google Scholar]
- Fitzgerald, J.R. Livestock-associated staphylococcus aureus: Origin, evolution and public health threat. Trends Microbiol. 2012, 20, 192–198. [Google Scholar] [CrossRef]
- Arunachalam, K.; Pandurangan, P.; Shi, C.; Lagoa, R. Regulation of Staphylococcus aureus virulence and application of Nanotherapeutics to Eradicate S. aureus infection. Pharmaceutics 2023, 15, 310. [Google Scholar] [CrossRef] [PubMed]
- Algammal, A.M.; Hetta, H.F.; Elkelish, A.; Alkhalifah, D.H.H.; Hozzein, W.N.; Batiha, G.E.; Nahhas, N.; Mabrok, M.A. Methicillin-resistant Staphylococcus aureus (MRSA): One Health perspective approach to the bacterium epidemiology, virulence factors, antibiotic-resistance, and zoonotic impact. Infect. Drug Resist. 2020, 13, 3255–3265. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, P.; Novais, C.; Peixe, L. Food-to-humans bacterial transmission. Microbiol. Spectr. 2020, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.P.; Humphrey, T.J.; Maskell, D.J. Molecular insights into farm animal and zoonotic Salmonella infections. Philos. Trans. R Soc. Lond B Biol. Sci. 2009, 364, 2709–2723. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC) Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 19 January 2023).
- Marois, C.; Bougeard, S.; Gottschalk, M.; Kobisch, M. Multiplex PCR assay for detection of Streptococcus suis species and serotypes 2 and 1/2 in tonsils of live and dead pigs. J. Clin. Microbiol. 2004, 42, 3169–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, M. Streptococcus suis: An emerging human threat. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.X.; Sun, L.; Grenier, D.; Yi, L. Streptococcus suis biofilm: Regulation, drug-resistance mechanisms, and disinfection strategies. Appl. Microbiol. Biotechnol. 2018, 102, 9121–9129. [Google Scholar] [CrossRef]
- Chekabab, S.M.; Paquin-Veillette, J.; Dozois, C.M.; Harel, J. The ecological habitat and transmission of Escherichia coli o157:H7. FEMS Microbio Lett. 2013, 341, 1–12. [Google Scholar] [CrossRef]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, united states, 1982-2002. Emerg. Infect Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Feng, C.; Qiu, S.; Xu, K.; Wang, C.; Liu, X.; Lv, J.; Yu, H.; Wu, S. Application and prospect of semiconductor biosensors in detection of viral zoonoses. J. Semicond. 2023, 44, 023102. [Google Scholar] [CrossRef]
- Arora, P.; Sindhu, A.; Kaur, H.; Dilbaghi, N.; Chaudhury, A. An overview of transducers as platform for the rapid detection of foodborne pathogens. Appl. Microbiol. Biotechnol. 2013, 97, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, S.; Gupta, P.; Kumar, N.; Kant, K. Progress in fluorescence biosensing and food safety towards point-of-detection (pod) system. Biosensors 2023, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, W.; Rong, Z.; Liu, X.; Gu, B.; Xiao, R.; Wang, S. Layer-by-layer assembly of magnetic-core dual quantum dot-shell nanocomposites for fluorescence lateral flow detection of bacteria. Nanoscale 2020, 12, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, J.; Xu, X.; Guo, L.; Xu, L.; Sun, M.; Hu, S.; Kuang, H.; Xu, C.; Li, A. An overview for the nanoparticles-based quantitative lateral flow assay. Small Methods 2022, 6, e2101143. [Google Scholar] [CrossRef]
- Chan, W.C.W.; Maxwell, D.J.; Gao, X.; Bailey, R.E.; Han, M.; Nie, S. Luminescent quantum dots for multiplexed biological detection and imaging. Curr. Opin. Biotechnol. 2002, 13, 40–46. [Google Scholar] [CrossRef]
- Tan, H.; Li, Q.; Ma, C.; Song, Y.; Xu, F.; Chen, S.; Wang, L. Lanthanide based dual-emission fluorescent probe for detection of mercury (II) in milk. Biosens. Bioelectron. 2015, 63, 566–571. [Google Scholar] [CrossRef]
- Liang, R.L.; Xu, X.P.; Liu, T.C.; Zhou, J.W.; Wang, X.G.; Ren, Z.Q.; Hao, F.; Wu, Y.S. Rapid and sensitive lateral flow immunoassay method for determining alpha fetoprotein in serum using europium (III) chelate microparticles-based lateral flow test strips. Anal. Chim. Acta 2015, 891, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Juntunen, E.; Myyryläinen, T.; Salminen, T.; Soukka, T.; Pettersson, K. Performance of fluorescent europium(III) nanoparticles and colloidal gold reporters in lateral flow bioaffinity assay. Anal. Biochem. 2012, 428, 31–38. [Google Scholar] [CrossRef]
- Huang, D.; Ying, H.; Jiang, D.; Liu, F.; Tian, Y.; Du, C.; Zhang, L.; Pu, X. Rapid and sensitive detection of interleukin-6 in serum via time-resolved lateral flow immunoassay. Anal Biochem. 2020, 588, 113468. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ma, B.; Wang, Y.; Chen, E.; Li, J.; Zhang, M. Research on rapid detection technology for β(2)-agonists: Multi-residue fluorescence immunochromatography based on dimeric artificial antigen. Foods 2022, 11, 863. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, B.; Liu, M.; Chen, E.; Xu, Y.; Zhang, M. Europium fluorescent nanoparticles-based multiplex lateral flow immunoassay for simultaneous detection of three antibiotic families residue. Front. Chem. 2021, 9, 793355. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, Z.; Liu, B.; Wang, C.; Wang, Q.; Zhang, L.; Wang, Z.; Chen, C.; Fu, Y.; Li, C.; et al. A time-resolved fluorescence lateral flow immunoassay for rapid and quantitative serodiagnosis of brucella infection in humans. J. Pharm. Biomed. Anal. 2021, 200, 114071. [Google Scholar] [CrossRef]
- Zhuang, L.; Gong, J.; Ji, Y.; Tian, P.; Kong, F.; Bai, H.; Gu, N.; Zhang, Y. Lateral flow fluorescent immunoassay based on isothermal amplification for rapid quantitative detection of Salmonella spp. Analyst 2020, 145, 2367–2377. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- Lillis, L.; Lehman, D.; Singhal, M.C.; Cantera, J.; Singleton, J.; Labarre, P.; Toyama, A.; Piepenburg, O.; Parker, M.; Wood, R.; et al. Non-instrumented incubation of a recombinase polymerase amplification assay for the rapid and sensitive detection of proviral HIV-1 DNA. PLoS ONE 2014, 9, e108189. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Huang, J.; Zhang, F.; Zhou, Y.; Huang, H. Detection of shrimp hemocyte iridescent virus by recombinase polymerase amplification assay. Mol. Cell. Probes 2020, 49, 101475. [Google Scholar] [CrossRef]
- Lillis, L.; Siverson, J.; Lee, A.; Cantera, J.; Parker, M.; Piepenburg, O.; Lehman, D.A.; Boyle, D.S. Factors influencing recombinase polymerase amplification (RPA) assay outcomes at point of care. Mol. Cell. Probes. 2016, 30, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Analyt. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Najafzadeh, M.J.; de Hoog, G.S.; de Camargo, Z.P. Rapid identification of emerging human-pathogenic Sporothrix species with rolling circle amplification. Front. Microbiol. 2015, 6, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Liu, M.; Tram, K.; Gu, J.; Salena, B.J.; Jiang, Y.; Li, Y. Optimal DNA templates for rolling circle amplification revealed by in vitro selection. Chemistry 2015, 21, 8069–8074. [Google Scholar] [CrossRef]
- Jaroenram, W.; Kiatpathomchai, W.; Flegel, T.W. Rapid and sensitive detection of white spot syndrome virus by loop-mediated isothermal amplification combined with a lateral flow dipstick. Mol. Cell. Probes. 2009, 23, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Vogel, R.F.; Niessen, L. Development and application of a loop-mediated isothermal amplification assay for rapid identification of aflatoxigenic molds and their detection in food samples. Int. J. Food Microbiol. 2012, 159, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Kolm, C.; Martzy, R.; Führer, M.; Mach, R.L.; Krska, R.; Baumgartner, S.; Farnleitner, A.H.; Reischer, G.H. Detection of a microbial source tracking marker by isothermal helicase-dependent amplification and a nucleic acid lateral-flow strip test. Sci. Rep. 2019, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, C.C.; Luo, X.; Ma, F.; Zhang, C.Y. 5-hydroxymethylcytosine glucosylation-triggered helicase-dependent amplification-based fluorescent biosensor for sensitive detection of β-glucosyltransferase with zero background signal. Anal. Chem. 2020, 92, 16307–16313. [Google Scholar] [CrossRef]
- Walker, G.T.; Fraiser, M.S.; Schram, J.L.; Little, M.C.; Nadeau, J.G.; Malinowski, D.P. Strand displacement amplification--an isothermal, in vitro DNA amplification technique. Nucleic Acids Res. 1992, 20, 1691–1696. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Liu, Y.; Yan, Y.; Serge, Y.O.; Zhou, X.; Xiang, X.; Ma, C. A highly sensitive method for the detection of alkaline phosphatase based on thioflavin t/g-quadruplex and strand displacement amplification. Microchem. J. 2022, 181, 107655. [Google Scholar] [CrossRef]
- Jin, B.; Ma, B.; Li, J.; Hong, Y.; Zhang, M. Simultaneous detection of five foodborne pathogens using a mini automatic nucleic acid extractor combined with recombinase polymerase amplification and lateral flow immunoassay. Microorganisms 2022, 10, 1352. [Google Scholar] [CrossRef] [PubMed]
- BAM Chapter 10. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-10-detection-listeria-monocytogenes-foods-and-environmental-samples-and-enumeration (accessed on 1 January 2023).
- BAM Chapter 12. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-12-staphylococcus-aureus (accessed on 1 January 2023).
- BAM Chapter 5. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-5-salmonella (accessed on 1 January 2023).
- BAM Chapter 4. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-4-enumeration-escherichia-coli-and-coliform-bacteria (accessed on 1 January 2023).
- GB/T 19915.2-2005; National Inspection and Quarantine Standards for Procedure for Isolation and Identification of Streptococcus suis Type 2. Standards Press of China: Beijing, China, 2005.
- Yang, W.; Cai, X.; Hao, Y.; Liu, Y.; Wang, S.; Xing, R.; Gu, J.; Li, C.; Yue, X.; Yuan, C.; et al. Characterization of Streptococcus suis serotype 2 blood infections using RT-qPCR to quantify glutamate dehydrogenase copy numbers. J. Microbiol. Methods 2010, 83, 326–329. [Google Scholar] [CrossRef]
- Härmä, H.; Soukka, T.; Lövgren, T. Europium nanoparticles and time-resolved fluorescence for ultrasensitive detection of pros-tate-specific antigen. Clin. Chem. 2001, 47, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.J.; Zhao, Q.; Zhou, R.; Liang, Y.C.; Zhao, W.B.; Shan, C.X. Ratiometric fluorescence sensor based on europium-grafted ZnO quantum dots for visual and colorimetric detection of tetracycline. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 259, 119901. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, J.; Li, Q.; Guo, J.; Zhang, G. Chapter 10-detection of microorganisms using recombinase polymerase amplification with lateral flow dipsticks. Methods Microbiol. 2020, 47, 319–349. [Google Scholar]
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase polymerase amplification for diagnostic applications. Clin. Chem. 2016, 62, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Munawar, M.A. Critical insight into recombinase polymerase amplification technology. Expert Rev. Mol. Diagn. 2022, 22, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Di, N.F.; Chiarello, M.; Cavalera, S.; Baggiani, C.; Anfossi, L. Ten years of lateral flow immunoassay technique applications: Trends, challenges and future perspectives. Sensors 2021, 21, 5185. [Google Scholar]
- Wang, Z.; Yao, X.; Wang, R.; Ji, Y.; Yue, T.; Sun, J.; Li, T.; Wang, J.; Zhang, D. Label-free strip sensor based on surface positively charged nitrogen-rich carbon nanoparticles for rapid detection of Salmonella enteritidis. Biosens. Bioelectron. 2019, 132, 360–367. [Google Scholar] [CrossRef]
- Niu, K.; Zheng, X.; Huang, C.; Xul, K.; Zhi, Y.; Shen, H.; Jia, N. A colloidal gold nanoparticle-based immunochromatographic test strip for rapid and convenient detection of Staphylococcus aureus. J. Nanosci. Nanotechnol. 2014, 14, 5151–5156. [Google Scholar] [CrossRef] [PubMed]
- Kanayeva, D.A.; Wang, R.; Rhoads, D.; Erf, G.F.; Slavik, M.F.; Tung, S.; Li, Y. Efficient separation and sensitive detection of listeria monocytogenes using an impedance immunosensor based on magnetic nanoparticles, a microfluidic chip, and an interdigitated microelectrode. J. Food Prot. 2012, 75, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Katani, R.; Li, L.; Hegde, N.; Roberts, E.L.; Kapur, V.; DebRoy, C. Rapid detection of Escherichia Coli O157 and Shiga toxins by lateral flow immunoassays. Toxins 2016, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Hao, H.J.; Xiong, G.H.; Geng, H.R.; Zheng, Y.L.; Wang, J.; Cao, Y.; Yang, Y.H.; Cai, X.H.; Jiang, Y.Q. Development of colloidal gold-based immunochromatographic assay for rapid detection of Streptococcus suis serotype 2. Vet. Immunol. Immunopathol. 2010, 133, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.B.; Du, X.J.; Zang, Y.X.; Li, P.; Wang, S. Sers-based lateral flow strip biosensor for simultaneous detection of listeria monocytogenes and Salmonella enterica serotype enteritidis. J. Agric. Food Chem. 2017, 65, 10290–10299. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Shin, J.H.; Park, J.K. Pressed paper-based dipstick for detection of foodborne pathogens with multistep reactions. Anal. Chem. 2016, 88, 3781–3788. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Song, D.; Xu, K.; Zheng, Y.; Xiao, H.; Liu, Y.; Li, J.; Song, X. Simultaneous detection of three zoonotic pathogens based on phage display peptide and multicolor quantum dots. Anal. Biochem. 2020, 608, 113854. [Google Scholar] [CrossRef]
- Hiremath, N.; Chin, B.A.; Park, M.K. Effect of competing foodborne pathogens on the selectivity and binding kinetics of a lytic phage for methicillin-resistant staphylococcus aureus detection. J. Electrochem. Soc. 2017, 164, B142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | Species | Serotype | ID of Strains | Quintuple RPA-EuNP-LFSBs Test Results | ||||
|---|---|---|---|---|---|---|---|---|
| hlyA | nuc | gdh | fimY | rfbE | ||||
| 1 | Listeria monocytogenes | 4b | ATCC 19115 | + | − | − | − | − |
| 2 | Listeria monocytogenes | 4b | ATCC 13932 | + | − | − | − | − |
| 3 | Listeria monocytogenes | 1/2a | ATCC 19111 | + | − | − | − | − |
| 4 | Listeria monocytogenes | 2 | ATCC 19112 | + | − | − | − | − |
| 5 | Staphylococcus aureus | ATCC 25923 | − | + | − | − | − | |
| 6 | Staphylococcus aureus | ATCC 35556 | − | + | − | − | − | |
| 7 | Staphylococcus aureus | 3 | CICC 12600 | − | + | − | − | − |
| 8 | Staphylococcus aureus | CICC 21648 | − | + | − | − | − | |
| 9 | Streptococcus suis | ATCC 700794 | − | − | + | − | − | |
| 10 | Streptococcus suis | ATCC 700796 | − | − | + | − | − | |
| 11 | Streptococcus suis | CJ 10102 | − | − | + | − | − | |
| 12 | Streptococcus suis | CJ 10217 | − | − | + | − | − | |
| 13 | Salmonella enterica | Enteritidis | ATCC 13076 | − | − | − | + | − |
| 14 | Salmonella enterica | Enteritidis | CICC 21513 | − | − | − | + | − |
| 15 | Salmonella enterica | Enteritidis | ATCC 29629 | − | − | − | + | − |
| 16 | Salmonella enterica | Enteritidis | ATCC 29631 | − | − | − | + | − |
| 17 | Escherichia coli | O157:H7 | ATCC 35150 | − | − | − | − | + |
| 18 | Escherichia coli | O157:H7 | ATCC 35218 | − | − | − | − | + |
| 19 | Escherichia coli | O157:H7 | CICC 24187 | − | − | − | − | + |
| 20 | Escherichia coli | O157:H7 | CICC 21530 | − | − | − | − | + |
| 21 | Bacillus coagulans | CICC 20138 | − | − | − | − | − | |
| 22 | Bacillus cereus | ATCC 10876a | − | − | − | − | − | |
| 23 | Bacillus cereus | ATCC 9139 | − | − | − | − | − | |
| 24 | Bacillus cereus | CICC 21261 | − | − | − | − | − | |
| 25 | Bacillus vallismortis | CICC 21224 | − | − | − | − | − | |
| 26 | Cronobacter sakazakii | CICC 24338 | − | − | − | − | − | |
| 27 | Cronobacter sakazakii | CICC 24125 | − | − | − | − | − | |
| 28 | Campylobacter jejuni | CICC 22936 | − | − | − | − | − | |
| 29 | Campylobacter jejuni | ATCC 49349 | − | − | − | − | − | |
| 30 | Campylobacter jejuni | CICC 22937 | − | − | − | − | − | |
| 31 | Clostridium perfringens | ATCC 13124 | − | − | − | − | − | |
| 32 | Enterobacter aerogenes | CICC 10293 | − | − | − | − | − | |
| 33 | Enterobacter aerogenes | CICC 10418 | − | − | − | − | − | |
| 34 | Enterobacter aerogenes | CICC 20051 | − | − | − | − | − | |
| 35 | Streptococcus pyogenes | CICC 10373 | − | − | − | − | − | |
| 36 | Streptococcus pyogenes | CICC 10356 | − | − | − | − | − | |
| 37 | Streptococcus mutans | CICC 10387 | − | − | − | − | − | |
| 38 | Shigella flexneri | CICC 10865 | − | − | − | − | − | |
| 39 | Shigella flexneri | CICC 21534 | − | − | − | − | − | |
| 40 | Shigella sonnei | CICC 21535 | − | − | − | − | − | |
| Samples | L. monocytogenes | S. aureus | S. suis | S. enterica | E. coli O157:H7 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| RPA-EuNP-LFSBs | Culture Method a | RPA-EuNP-LFSBs | Culture Method a | RPA-EuNP-LFSBs | Culture Method b | RPA-EuNP-LFSBs | Culture Method a | RPA-EuNP-LFSBs | Culture Method a | |
| Chicken-1 | − | − | − | − | − | − | − | − | − | − |
| Chicken-2 | − | − | − | − | − | − | − | − | + | + |
| Chicken-3 | − | − | − | − | − | − | − | − | − | − |
| Pork-1 | − | − | + | + | − | − | − | − | + | + |
| Pork-2 | − | − | − | − | − | − | − | − | − | − |
| Pork-3 | − | − | − | − | − | − | − | − | − | − |
| Beef-1 | − | − | − | − | − | − | − | − | − | − |
| Beef-2 | − | − | − | − | − | − | − | − | − | − |
| Beef-3 | − | − | − | − | − | − | − | − | − | − |
| Lamb-1 | − | − | − | − | − | − | − | − | − | − |
| Lamb-2 | − | − | − | − | − | − | − | − | − | − |
| Lamb-3 | − | − | − | − | − | − | − | − | − | − |
| Duck-1 | − | − | − | − | − | − | − | − | − | − |
| Duck-2 | − | − | − | − | − | − | − | − | − | − |
| Duck-3 | − | − | − | − | − | − | − | − | − | − |
| Milk-1 | − | − | − | − | − | − | − | − | − | − |
| Milk-2 | − | − | − | − | − | − | − | − | − | − |
| Milk-3 | − | − | − | − | − | − | − | − | − | − |
| Total | − | − | 1 | 1 | − | − | − | − | 2 | 2 |
| Positive Detection rate | 0% | 0% | 6.7% | 6.7% | 0% | 0% | 0% | 0% | 13.3% | 13.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, B.; Ma, B.; Mei, Q.; Xu, S.; Deng, X.; Hong, Y.; Li, J.; Xu, H.; Zhang, M. Europium Nanoparticle-Based Lateral Flow Strip Biosensors Combined with Recombinase Polymerase Amplification for Simultaneous Detection of Five Zoonotic Foodborne Pathogens. Biosensors 2023, 13, 652. https://doi.org/10.3390/bios13060652
Jin B, Ma B, Mei Q, Xu S, Deng X, Hong Y, Li J, Xu H, Zhang M. Europium Nanoparticle-Based Lateral Flow Strip Biosensors Combined with Recombinase Polymerase Amplification for Simultaneous Detection of Five Zoonotic Foodborne Pathogens. Biosensors. 2023; 13(6):652. https://doi.org/10.3390/bios13060652
Chicago/Turabian StyleJin, Bei, Biao Ma, Qing Mei, Shujuan Xu, Xin Deng, Yi Hong, Jiali Li, Hanyue Xu, and Mingzhou Zhang. 2023. "Europium Nanoparticle-Based Lateral Flow Strip Biosensors Combined with Recombinase Polymerase Amplification for Simultaneous Detection of Five Zoonotic Foodborne Pathogens" Biosensors 13, no. 6: 652. https://doi.org/10.3390/bios13060652
APA StyleJin, B., Ma, B., Mei, Q., Xu, S., Deng, X., Hong, Y., Li, J., Xu, H., & Zhang, M. (2023). Europium Nanoparticle-Based Lateral Flow Strip Biosensors Combined with Recombinase Polymerase Amplification for Simultaneous Detection of Five Zoonotic Foodborne Pathogens. Biosensors, 13(6), 652. https://doi.org/10.3390/bios13060652