
Chemical, Cytotoxic, and Anti-Inflammatory Assessment of Honey Bee Venom from Apis mellifera intermissa


 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Peptide and Enzymatic Composition of Honey Bee Venom
2.2. Metal Content of Honey Bee Venom
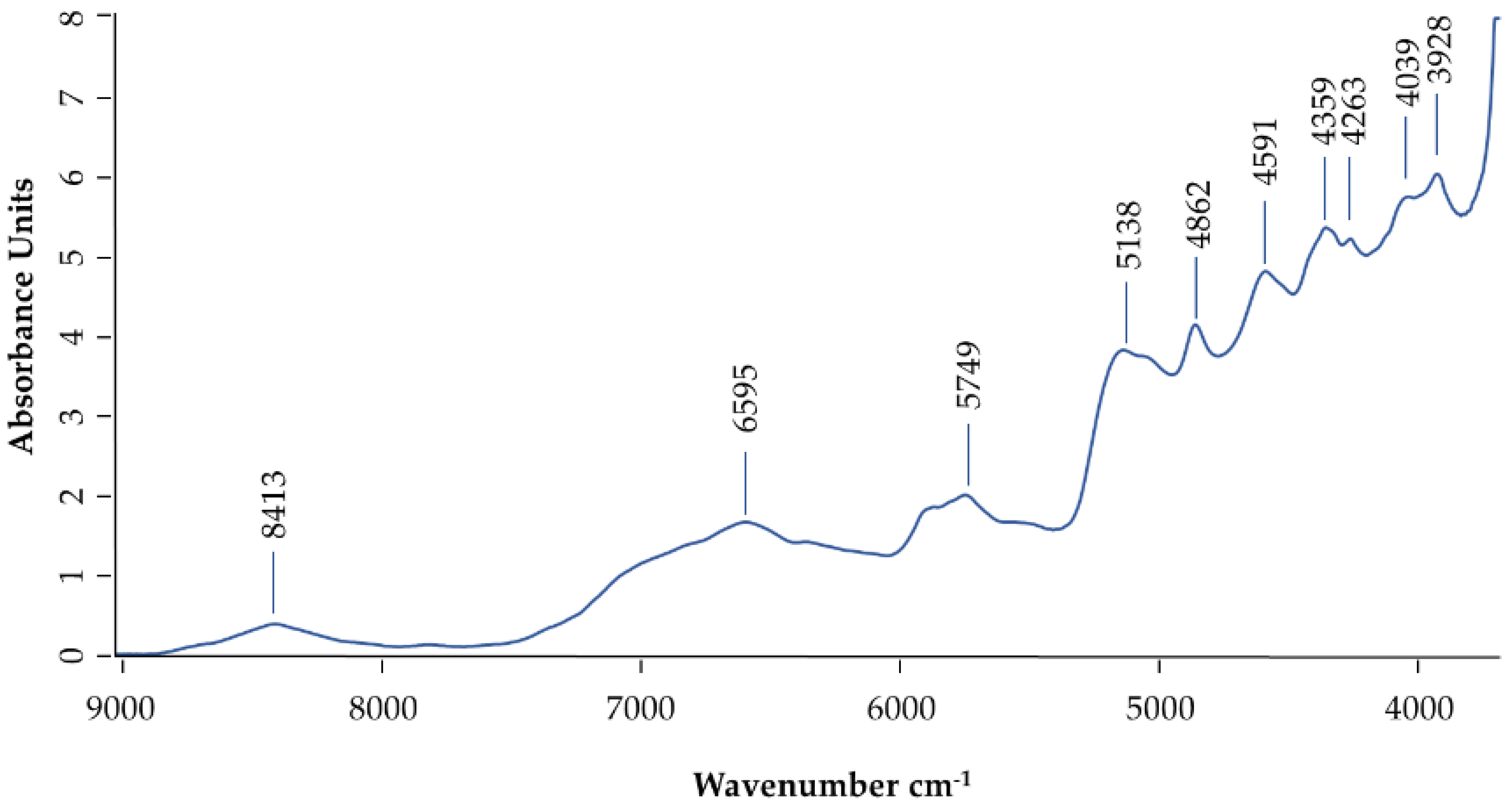
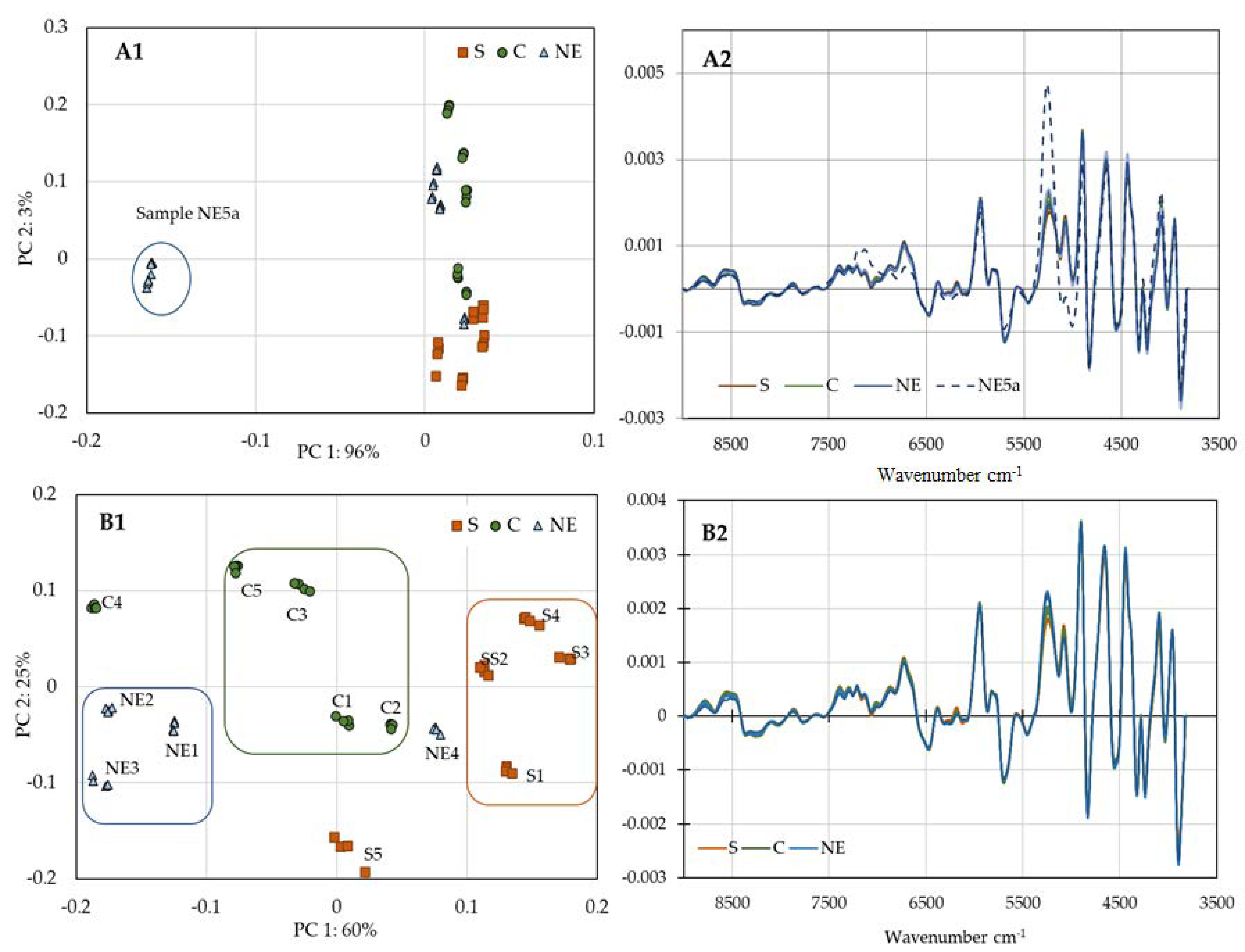
2.3. NIR Evaluation of Honey Bee Venom
2.4. Biological Activity
2.4.1. Cytotoxic Activity
2.4.2. Anti-Inflammatory Activity
3. Materials and Methods
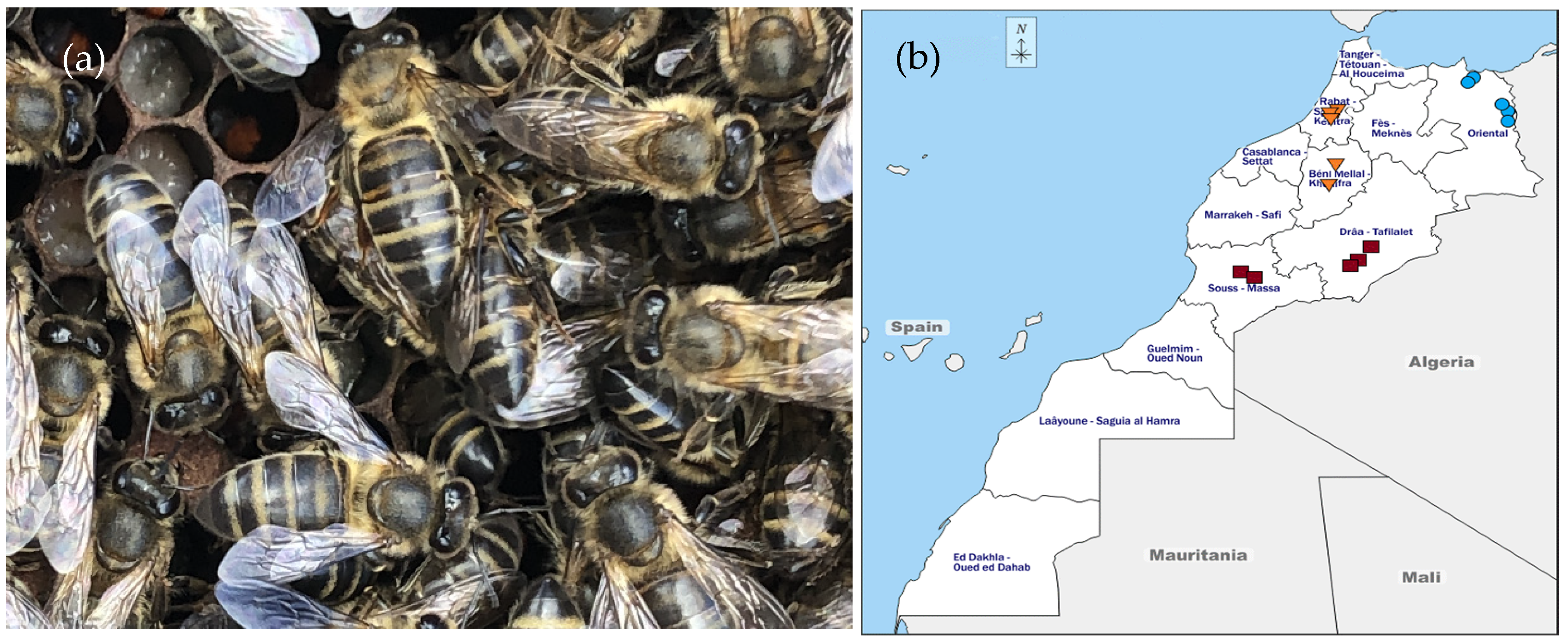
3.1. Sample Collection
3.2. Standards and Reagents
3.3. Cell Lines
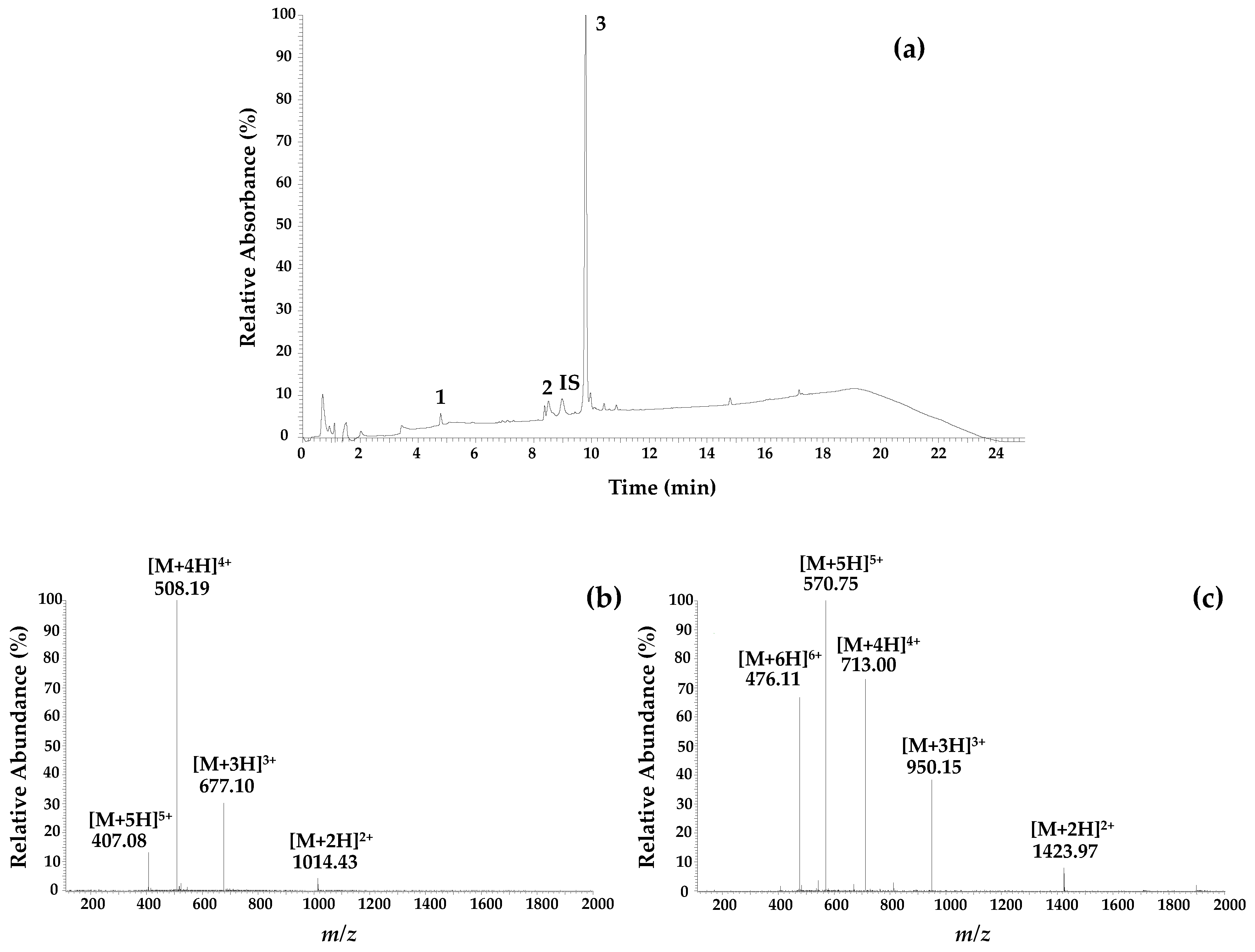
3.4. Chemical Characterization of the Samples by LC-ESI/MSn
3.5. Metal Content by Atomic Absorption Spectroscopy (AAS)
3.6. Near-Infrared Analysis of Honey Bee Venom (NIR):
3.7. Citotoxic Activity
3.8. Anti-Inflamatory Activity
3.9. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gajski, G.; Garaj-Vrhovac, V. Bee venom induced cytogenetic damage and decreased cell viability in human white blood cells after treatment in vitro: A multi-biomarker approach. Environ. Toxicol. Pharmacol. 2011, 32, 201–211. [Google Scholar] [CrossRef]
- Lee, Y.M.; Cho, S.-N.; Son, E.; Song, C.-H.; Kim, D.-S. Apamin from bee venom suppresses inflammation in a murine model of gouty arthritis. J. Ethnopharmacol. 2020, 257, 112860. [Google Scholar] [CrossRef]
- Matysiak, J.; Hajduk, J.; Swiatly, A.; Naskret, N.; Kokot, Z. Proteomic analysis of Apis mellifera venom determined by liquid chromatography (LC) coupled with nano-LC-MALDI-TOF/TOF MS. Acta Pol. Pharm. Drug Res. 2017, 74, 53–65. [Google Scholar]
- Oršolić, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef]
- Rady, I.; Siddiqui, I.A.; Rady, M.; Mukhtar, H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. 2017, 402, 16–31. [Google Scholar] [CrossRef] [Green Version]
- Resende, V.M.F.; Vasilj, A.; Santos, K.S.; Palma, M.S.; Shevchenko, A. Proteome and phosphoproteome of Africanized and European honeybee venoms. Proteomics 2013, 13, 2638–2648. [Google Scholar] [CrossRef]
- Laxme, R.S.; Suranse, V.; Sunagar, K. Arthropod venoms: Biochemistry, ecology and evolution. Toxicon 2019, 158, 84–103. [Google Scholar] [CrossRef]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Kil Lee, C.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef]
- Yang, J.; Lee, K.S.; Kim, B.Y.; Choi, Y.S.; Yoon, H.J.; Jia, J.; Jin, B.R. Anti-fibrinolytic and anti-microbial activities of a serine protease inhibitor from honeybee (Apis cerana) venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 201, 11–18. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Ye, Y.; Wang, X.-R.; Lin, L.-T.; Xiao, L.-Y.; Zhou, P.; Shi, G.-X.; Liu, C.-Z. Bee venom therapy: Potential mechanisms and therapeutic applications. Toxicon 2018, 148, 64–73. [Google Scholar] [CrossRef]
- Nader, R.; Mackieh, R.; Wehbe, R.; El Obeid, D.; Sabatier, J.; Fajloun, Z. Beehive Products as Antibacterial Agents: A Review. Antibiot. 2021, 10, 717. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M. Bee products-chemical and biological properties. In Bee Products-Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 1–306. [Google Scholar]
- El-Hanoun, A.; El-Komy, A.; El-Sabrout, K.; Abdella, M. Effect of bee venom on reproductive performance and immune response of male rabbits. Physiol. Behav. 2020, 223, 112987. [Google Scholar] [CrossRef]
- Frangieh, J.; Salma, Y.; Haddad, K.; Mattei, C.; Legros, C.; Fajloun, Z.; El Obeid, D. First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region. Toxins 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; An, H.-J.; Kim, J.-Y.; Kim, W.-H.; Gwon, M.-G.; Kim, H.-J.; Han, S.M.; Park, I.; Park, S.C.; Leem, J.; et al. Bee venom attenuates Porphyromonas gingivalis and RANKL-induced bone resorption with osteoclastogenic differentiation. Food Chem. Toxicol. 2019, 129, 344–353. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M.; Kaleli, S. Anti-inflammatory and antioxidative properties of honey bee venom on Freund’s Complete Adjuvant-induced arthritis model in rats. Toxicon 2019, 161, 4–11. [Google Scholar] [CrossRef]
- Lee, J.E.; Shah, V.K.; Lee, E.-J.; Oh, M.-S.; Choi, J.J. Melittin—A bee venom component—Enhances muscle regeneration factors expression in a mouse model of skeletal muscle contusion. J. Pharmacol. Sci. 2019, 140, 26–32. [Google Scholar] [CrossRef]
- Yang, W.; Hu, F.L.; Xu, X.F. Bee venom and SARS-CoV-2. Toxicon 2020, 181, 69–70. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M. Anti-fungal properties and mechanisms of melittin. Appl. Microbiol. Biotechnol. 2020, 104, 6513–6526. [Google Scholar] [CrossRef]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Barour, C.; Tahar, A.; Radloff, S.E.; Hepburn, H.R. Multivariate analysis of honeybees, Apis mellifera Linnaeus (Hymenoptera: Apidae) of the northeastern and southern regions of Algeria. Afr. Entomol. 2005, 13, 17–23. [Google Scholar]
- Gupta, R.K.; Reybroeck, W.; van Veen, J.W.; Gupta, A. Beekeeping for Poverty Alleviation and Livelihood Security: Technological Aspects of Beekeeping; Springer Science+Business Media: Dordrecht, The Netherlands, 2014; Volume 1. [Google Scholar]
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer: Heidelberg, Germany, 1988. [Google Scholar]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhao, J.; Zhang, S.; Shen, J.; Qi, Y.; Xue, X.; Li, Y.; Wu, L.; Zhang, J.; Chen, F.; et al. Quantification of melittin and apamin in bee venom lyophilized powder from Apis mellifera by liquid chromatography–diode array detector–tandem mass spectrometry. Anal. Biochem. 2010, 404, 171–178. [Google Scholar] [CrossRef]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.J.R.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Ionete, R.E.; Dinca, O.R.; Tamaian, R.; Geana, E.I. Exploring Apis Mellifera venom compounds using highly efficient methods. Prog. Cryog. Isot. Sep. 2013, 16, 89–100. [Google Scholar]
- Kokot, Z.; Matysiak, J. Simultaneous Determination of Major Constituents of Honeybee Venom by LC-DAD. Chromatographia 2009, 69, 1401–1405. [Google Scholar] [CrossRef]
- Rybak-Chmielewska, H.; Szczêsna, T. HPLC study of chemical composition of honeybee (Apis mellifera L.) venom. J. Apic. Sci. 2004, 48, 103–109. [Google Scholar]
- Kokot, Z.; Matysiak, J. Inductively coupled plasma mass spectrometry determination of metals in honeybee venom. J. Pharm. Biomed. Anal. 2008, 48, 955–959. [Google Scholar] [CrossRef]
- Raghuram, P.; Sriramulu, J. Heavy metals testing in active pharmaceutical ingredients: An alternate approach. Pharmazie 2010, 65, 15–18. [Google Scholar]
- Balaram, V. Recent advances in the determination of elemental impurities in pharmaceuticals—Status, challenges and moving frontiers. TrAC Trends Anal. Chem. 2016, 80, 83–95. [Google Scholar] [CrossRef]
- Wollein, U.; Bauer, B.; Habernegg, R.; Schramek, N. Potential metal impurities in active pharmaceutical substances and finished medicinal products—A market surveillance study. Eur. J. Pharm. Sci. 2015, 77, 100–105. [Google Scholar] [CrossRef]
- Teasdale, A.; Thompson, S. ICH Q3D Elemental Impurities. In ICH Quality Guidelines; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 233–280. [Google Scholar]
- Anjos, O.; Caldeira, I.; Roque, R.; Pedro, S.; Lourenço, S.; Canas, S. Screening of Different Ageing Technologies of Wine Spirit by Application of Near-Infrared (NIR) Spectroscopy and Volatile Quantification. Processes 2020, 8, 736. [Google Scholar] [CrossRef]
- Barra, I.; Kharbach, M.; Qannari, E.M.; Hanafi, M.; Cherrah, Y.; Bouklouze, A. Predicting cetane number in diesel fuels using FTIR spectroscopy and PLS regression. Vib. Spectrosc. 2020, 111, 103157. [Google Scholar] [CrossRef]
- Pasquini, C. Near infrared spectroscopy: A mature analytical technique with new perspectives—A review. Anal. Chim. Acta 2018, 1026, 8–36. [Google Scholar] [CrossRef]
- Osborne, B.G.; Fearn, T.; Hindle, P.H. Practical NIR Spectroscopy with Applications in Food and Beverage Analysis; Longman Scientific & Technical: Essex, UK, 1993. [Google Scholar]
- Workman, J.J., Jr. Interpretive spectroscopy for near infrared. Appl. Spectrosc. Rev. 1996, 31, 251–320. [Google Scholar] [CrossRef]
- Johansson, J.; Larsson, M.H.; Hornberg, J.J. Predictive in vitro toxicology screening to guide chemical design in drug discovery. Curr. Opin. Toxicol. 2019, 15, 99–108. [Google Scholar] [CrossRef]
- McKim Jr, J.M. Building a tiered approach to in vitro predictive toxicity screening: A focus on assays with in vivo relevance. Comb. Chem. High Throughput Screen. 2010, 13, 188–206. [Google Scholar] [CrossRef] [Green Version]
- Niles, A.L.; Moravec, R.A.; Riss, T.L. Update on in vitro cytotoxicity assays for drug development. Expert Opin. Drug Discov. 2008, 3, 655–669. [Google Scholar] [CrossRef]
- Cai, L.; Qin, X.; Xu, Z.; Song, Y.; Jiang, H.; Wu, Y.; Ruan, H.; Chen, J. Comparison of Cytotoxicity Evaluation of Anticancer Drugs between Real-Time Cell Analysis and CCK-8 Method. ACS Omega 2019, 4, 12036–12042. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, D.; Xie, L.; Zhang, R. Effect of honey bee venom on proliferation of K1735M2 mouse melanoma cells in-vitro and growth of murine B16 melanomas in-vivo. J. Pharm. Pharmacol. 2010, 54, 1083–1089. [Google Scholar] [CrossRef]
- Hu, H.; Chen, D.; Li, Y.; Zhang, X. Effect of polypeptides in bee venom on growth inhibition and apoptosis induction of the human hepatoma cell line SMMC-7721 in-vitro and Balb/c nude mice in-vivo. J. Pharm. Pharmacol. 2010, 58, 83–89. [Google Scholar] [CrossRef]
- Moon, D.-O.; Park, S.-Y.; Lee, K.-J.; Heo, M.-S.; Kim, K.-C.; Kim, M.-O.; Lee, J.-D.; Choi, Y.H.; Kim, G.-Y. Bee venom and melittin reduce proinflammatory mediators in lipopolysaccharide-stimulated BV2 microglia. Int. Immunopharmacol. 2007, 7, 1092–1101. [Google Scholar] [CrossRef]
- Tu, W.-C.; Wu, C.-C.; Hsieh, H.-L.; Chen, C.-Y.; Hsu, S.-L. Honeybee venom induces calcium-dependent but caspase-independent apoptotic cell death in human melanoma A2058 cells. Toxicon 2008, 52, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jin, H.; Qian, Y.; Qi, S.; Luo, H.; Luo, Q.; Zhang, Z. Hybrid Melittin Cytolytic Peptide-Driven Ultrasmall Lipid Nanoparticles Block Melanoma Growth in Vivo. ACS Nano 2013, 7, 5791–5800. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.E.; Hwang, C.J.; Gu, S.M.; Park, M.H.; Kim, J.H.; Park, J.H.; Ahn, Y.J.; Kim, J.Y.; Song, M.J.; Song, H.S.; et al. Cancer Cell Growth Inhibitory Effect of Bee Venom via Increase of Death Receptor 3 Expression and Inactivation of NF-kappa B in NSCLC Cells. Toxins 2014, 6, 2210–2228. [Google Scholar] [CrossRef] [PubMed]
- Amini, E.; Baharara, J.; Nikdel, N.; Abdollahi, F.S. Cytotoxic and Pro-Apoptotic Effects of Honey Bee Venom and Chrysin on Human Ovarian Cancer Cells. Asia Pac. J. Med Toxicol. 2015, 4, 68–73. [Google Scholar]
- Kuete, V.; Karaosmanoğlu, O.; Sivas, H. Anticancer Activities of African Medicinal Spices and Vegetables, in Medicinal Spices and Vegetables from Africa: Therapeutic Potential against Metabolic, Inflammatory, Infectious and Systemic Diseases; Academic Press: London, UK, 2017; pp. 271–297. [Google Scholar]
- Orellana, E.A.; Kasinski, A.L. Sulforhodamine B (SRB) Assay in Cell Culture to Investigate Cell Proliferation. Bio-protocol 2016, 6, e1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, R.M.; Ferreira, I.; Calhelha, R.C.; Lima, R.T.; Vasconcelos, M.H.; Adega, F.; Chaves, R.; Queiroz, M.-J.R. Anti-hepatocellular carcinoma activity using human HepG2 cells and hepatotoxicity of 6-substituted methyl 3-aminothieno[3,2-b]pyridine-2-carboxylate derivatives: In vitro evaluation, cell cycle analysis and QSAR studies. Eur. J. Med. Chem. 2011, 46, 5800–5806. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.-H.; Shin, M.-C.; Lim, S.; Han, S.-M.; Park, H.-J.; Shin, I.; Lee, J.-S.; Kim, K.-A.; Kim, E.-H.; Kim, C.-J. Bee Venom Induces Apoptosis and Inhibits Expression of Cyclooxygenase-2 mRNA in Human Lung Cancer Cell Line NCI-H1299. J. Pharmacol. Sci. 2003, 91, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Oršolić, N. Potentiation of Bleomycin Lethality in HeLa and V79 Cells by Bee Venom. Arch. Ind. Hyg. Toxicol. 2009, 60, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, W.A.; Abd-Elhakim, Y.M.; Ismail, S.A. Involvement of the anti-inflammatory, anti-apoptotic, and anti-secretory activity of bee venom in its therapeutic effects on acetylsalicylic acid-induced gastric ulceration in rats. Toxicology 2019, 419, 11–23. [Google Scholar] [CrossRef]
- Rached, I.C.F.S.; Castro, F.M.; Guzzo, M.L.; de Mello, S.B.V. Anti-inflammatory effect of bee venom on antigen-induced arthritis in rabbits: Influence of endogenous glucocorticoids. J. Ethnopharmacol. 2010, 130, 175–178. [Google Scholar] [CrossRef]
- Yoon, S.-Y.; Kim, H.-W.; Roh, D.-H.; Kwon, Y.-B.; Jeong, T.-O.; Han, H.-J.; Lee, H.-J.; Choi, S.-M.; Ryu, Y.-H.; Beitz, A.J.; et al. The anti-inflammatory effect of peripheral bee venom stimulation is mediated by central muscarinic type 2 receptors and activation of sympathetic preganglionic neurons. Brain Res. 2005, 1049, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Ceremuga, M.; Stela, M.; Janik, E.; Gorniak, L.; Synowiec, E.; Sliwinski, T.; Sitarek, P.; Saluk-Bijak, J.; Bijak, M. Melittin—A Natural Peptide from Bee Venom Which Induces Apoptosis in Human Leukaemia Cells. Biomolecules 2020, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Daniluk, K.; Kutwin, M.; Grodzik, M.; Wierzbicki, M.; Strojny, B.; Szczepaniak, J.; Bałaban, J.; Sosnowska, M.; Chwalibog, A.; Sawosz, E.; et al. Use of Selected Carbon Nanoparticles as Melittin Carriers for MCF-7 and MDA-MB-231 Human Breast Cancer Cells. Materials 2019, 13, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, H.; Han, S.M.; Park, K.-K. Therapeutic Effects of Apamin as a Bee Venom Component for Non-Neoplastic Disease. Toxins 2020, 12, 195. [Google Scholar] [CrossRef] [Green Version]
- Wehbe, R.; Frangieh, J.; Rima, M.; El Obeid, D.; Sabatier, J.-M.; Fajloun, Z. Bee Venom: Overview of Main Compounds and Bioactivities for Therapeutic Interests. Molecules 2019, 24, 2997. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.R.; Ren, Y.F.; Dong, G.M.; Yang, R.J.; Liu, H.X.; Du, Y.H.; Zhang, W.Y. Determination of Methanol in Alcoholic Beverages by Two-Dimensional Near-Infrared Correlation Spectroscopy. Anal. Lett. 2016, 49, 2279–2289. [Google Scholar] [CrossRef]
- Alonso-Esteban, J.I.; Pinela, J.; Barros, L.; Ciric, A.; Sokovic, M.; Calhelha, R.C.; Torija-Isasa, E.; Sánchez-Mata, M.D.C.; Ferreira, I.C. Phenolic composition and antioxidant, antimicrobial and cytotoxic properties of hop (Humulus lupulus L.) Seeds. Ind. Crop. Prod. 2019, 134, 154–159. [Google Scholar] [CrossRef]
- Falcão, S.I.; Calhelha, R.C.; Touzani, S.; Lyoussi, B.; Ferreira, I.C.F.R.; Vilas-Boas, M. In Vitro Interactions of Moroccan Propolis Phytochemical’s on Human Tumor Cell Lines and Anti-Inflammatory Properties. Biomolecules 2019, 9, 315. [Google Scholar] [CrossRef] [Green Version]
 northeast,
northeast,  center and
center and  southern samples.
northeast, center and southern samples.
southern samples.
northeast, center and southern samples.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Apamin (µg/mL) | PLA2 * (µg/mL) | Melittin (µg/mL) |
|---|---|---|---|
| NE1 | 2.08 | 9.13 | 67.16 |
| NE2 | 1.93 | 6.66 | 73.43 |
| NE3 | 2.29 | 6.57 | 78.14 |
| NE4 | 1.99 | 7.4 | 73.85 |
| NE5 | 1.96 | 8.54 | 65.97 |
| Average ± SD | 2.1 ± 0.1 | 8 ± 1 | 72 ± 5 |
| C1 | 1.86 | 4.21 | 71.78 |
| C2 | 2.05 | 5.05 | 73.66 |
| C3 | 1.96 | 7.14 | 71.98 |
| C4 | 2.10 | 8.03 | 75.10 |
| C5 | 1.99 | 6.42 | 70.15 |
| Average ± SD | 2.0 ± 0.1 | 6 ± 1 | 73 ± 2 |
| S1 | 1.53 | 4.52 | 66.78 |
| S2 | 1.69 | 4.18 | 71.29 |
| S3 | 2.55 | 7.81 | 87.70 |
| S4 | 1.79 | 1.79 | 70.29 |
| S5 | 1.45 | 4.45 | 62.34 |
| Average ± SD | 1.8 ± 0.4 | 5 ± 2 | 72 ± 9 |
| [26] | 1.5 | 8 | 65 |
| [27] | 4.1 | 15 | 58 |
| [28] | 2.6 | 13 | 54 |
| [29] | 3.5 | 13 | 65 |
| Samples | K * (mg/g) | Na * (mg/g) | Ca * (mg/g) | Mg * (mg/g) | Zn * (mg/g) | Cu (µg/g) | Cd (µg/g) | Cr (µg/g) | Mn (µg/g) | Ni (µg/g) | Pb * (µg/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NE1 | 2.97 | 1.30 | 3.25 | 0.52 | 1.23 | 5.37 | - | 5.20 | 2.02 | 26.98 | 6.13 |
| NE2 | 3.01 | 1.34 | 1.78 | 0.44 | 1.39 | 24.72 | - | 9.46 | 4.46 | 4.32 | 5.15 |
| NE3 | 2.64 | 1.53 | 1.80 | 0.33 | 1.05 | 4.36 | - | 7.61 | 1.61 | 1.36 | 9.85 |
| NE4 | 2.16 | 1.00 | 0.95 | 0.32 | 1.04 | - | 2.26 | 3.86 | 1.10 | - | 8.08 |
| NE5 | 3.57 | 1.55 | 1.76 | 0.57 | 1.03 | 4.13 | - | 8.63 | 2.79 | 0.49 | 6.77 |
| Average ± SD | 2.9 ± 0.5 | 1.3 ± 0.2 | 1.9 ± 0.7 | 0.4 ± 0.1 | 1.2 ± 0.2 | 8 ± 9 | - | 7 ± 2 | 2.4 ± 1.2 | 8 ± 11 | 7.2 ± 1.6 |
| C1 | 1.62 | 1.93 | 4.55 | 0.42 | 1.03 | 9.45 | - | 4.59 | 2.47 | 2.83 | 3.81 |
| C2 | 1.73 | 1.01 | 2.16 | 0.27 | 1.12 | - | - | 3.41 | 1.59 | 0.64 | 3.56 |
| C3 | 3.02 | 1.84 | 3.22 | 0.55 | 1.42 | 20.91 | - | 6.71 | 3.10 | 0.62 | 3.99 |
| C4 | 3.03 | 1.43 | 2.32 | 0.56 | 1.27 | 6.83 | - | 4.99 | 2.07 | 4.49 | 4.84 |
| C5 | 2.50 | 0.98 | 2.59 | 0.38 | 1.53 | 7.30 | - | 8.29 | 3.76 | 1.78 | 5.16 |
| Average ± SD | 2.4 ± 0.6 | 1.4 ± 0.4 | 3.0 ± 0.9 | 0.4 ± 0.1 | 1.3 ± 0.2 | 9 ± 7 | - | 6 ± 2 | 2.6 ± 0.8 | 2.1 ± 1.5 | 4.3 ± 0.6 |
| S1 | 1.55 | 0.79 | 0.96 | 0.28 | 0.93 | - | - | 2.40 | 0.78 | - | 7.60 |
| S2 | 1.68 | 0.81 | 0.97 | 0.27 | 1.03 | 3.95 | - | 2.45 | 1.19 | - | 3.63 |
| S3 | 1.95 | 0.81 | 0.83 | 0.31 | 0.95 | 4.10 | 1.32 | 13.21 | 0.89 | - | 3.24 |
| S4 | 1.72 | 0.92 | 1.41 | 0.25 | 1.07 | - | - | 5.19 | 1.29 | - | 4.32 |
| S5 | 1.56 | 0.79 | 0.95 | 0.25 | 1.07 | 3.95 | - | 5.55 | 2.19 | - | 4.20 |
| Average ± SD | 1.7 ± 0.2 | 0.83 ± 0.05 | 1.0 ± 0.2 | 0.3 ± 0.0 | 1.1 ± 0.1 | 2.4 ± 2 | - | 6 ± 4 | 1.3 ± 0.5 | - | 4.6 ± 1.6 |
| Class | Oral | Parenteral | Inhalation | HBV Concentration | ||
|---|---|---|---|---|---|---|
| Cd | 1 | 0.5 | 0.2 | 0.3 | NE | a |
| C | - | |||||
| S | a | |||||
| Pb | 1 | 0.5 | 0.5 | 0.5 | NE | 7.3 * |
| C | 4.3 * | |||||
| S | 4.6 * | |||||
| Ni | 2A | 20 | 2 | 0.5 | NE | 8.2 |
| C | 2.1 | |||||
| S | - | |||||
| Cu | 3 | 300 | 30 | 3 | NE | 8.6 |
| C | 8.9 | |||||
| S | 2.4 | |||||
| Cr | 3 | 1100 | 110 | 0.3 | NE | 6.5 |
| C | 5.6 | |||||
| S | 5.8 | |||||
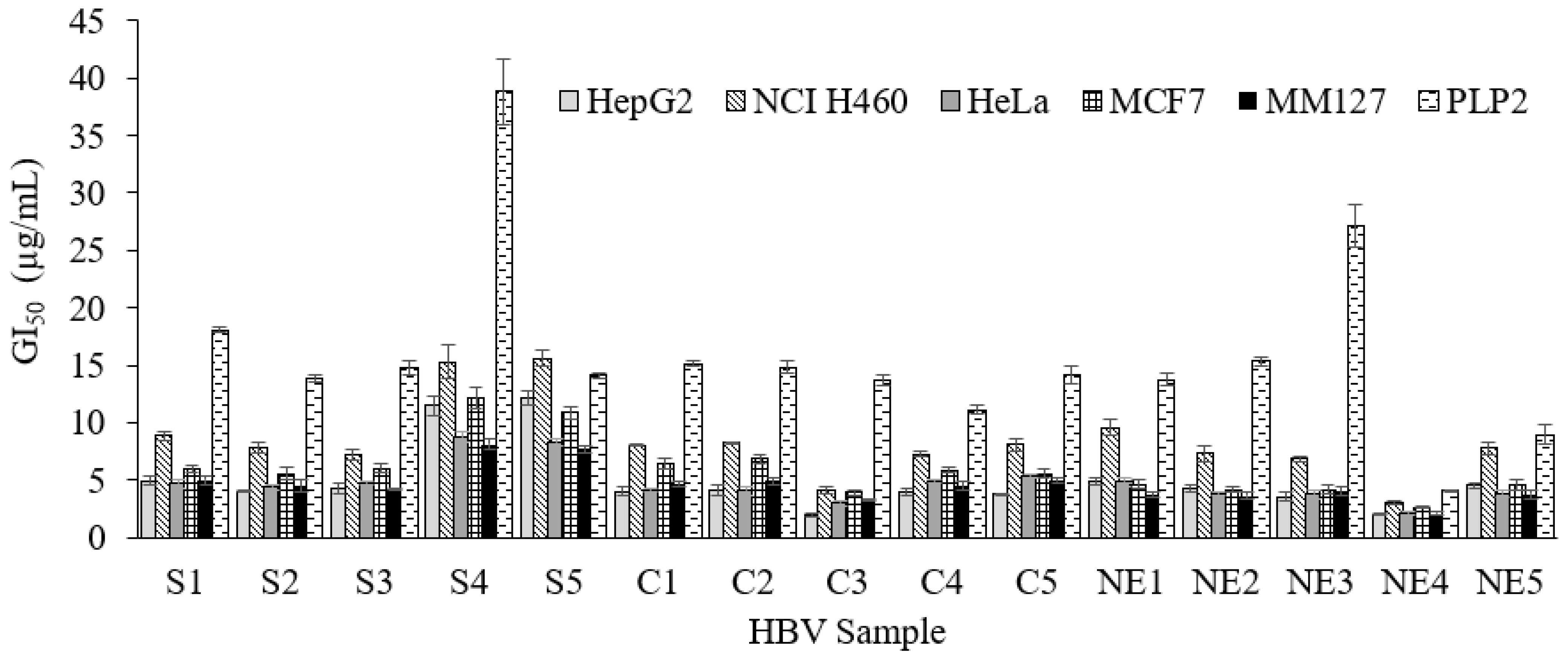
| Cytotoxic Activity (GI50, µg/mL) | Anti-Inflammatory Activity (IC50, µg/mL) | ||||||
|---|---|---|---|---|---|---|---|
| HepG2 | NCI_H460 | HeLa | MM127 * | MCF7 * | PLP2 | RAW264.7 * | |
| NE1 | 4.86 | 9.57 | 4.84 | 3.72 | 4.64 | 13.77 | 5.07 |
| NE2 | 4.32 | 7.35 | 3.88 | 3.64 | 4.2 | 15.33 | 4.89 |
| NE3 | 3.58 | 6.88 | 3.75 | 4.01 | 4.17 | 27.15 | 6.06 |
| NE4 | 2.40 | 3.09 | 2.08 | 2.05 | 2.62 | 4.06 | 4.03 |
| NE5 | 4.56 | 7.77 | 3.81 | 3.80 | 4.55 | 8.96 | 4.86 |
| Average ± SD | 3.9 ± 0.9 | 7 ± 2 | 3.7 ± 0.9 | 3.4 ± 0.7 | 4.0 ± 0.7 | 14 ± 8 | 5.0 ± 0.7 |
| C1 | 4.02 | 8.06 | 4.15 | 4.64 | 6.40 | 15.13 | 7.92 |
| C2 | 4.1 | 8.26 | 4.06 | 4.92 | 6.86 | 14.84 | 7.78 |
| C3 | 1.99 | 4.12 | 2.97 | 3.33 | 3.91 | 13.74 | 6.12 |
| C4 | 3.94 | 7.27 | 4.94 | 4.44 | 5.9 | 11.13 | 6.26 |
| C5 | 3.79 | 8.08 | 5.36 | 4.95 | 5.59 | 14.14 | 6.11 |
| Average ± SD | 3.6 ± 0.8 | 7 ± 2 | 4.3 ± 0.8 | 4.5 ± 0.6 | 5.7 ± 1.0 | 14 ± 1 | 6.8 ± 0.8 |
| S1 | 4.95 | 8.85 | 4.67 | 4.99 | 5.94 | 18.03 | 9.08 |
| S2 | 3.98 | 7.85 | 4.36 | 4.49 | 5.52 | 13.93 | 7.40 |
| S3 | 4.33 | 7.17 | 4.67 | 4.24 | 6.02 | 14.86 | 6.65 |
| S4 | 11.46 | 15.3 | 8.71 | 8.09 | 12.1 | 38.85 | 15.5 |
| S5 | 15.63 | 15.6 | 8.35 | 7.68 | 10.8 | 14.12 | 15.0 |
| Average ± SD | 8.0 ± 4.7 | 11 ± 4 | 6.2 ± 1.9 | 5.9 ± 1.7 | 8.1 ± 2.8 | 20 ± 10 | 10.7 ± 3.8 |
| Dexamethasone | 15.5 | ||||||
| HepG2 | NCI-H460 | HeLa | MCF-7 | MM127 | PLP2 | RAW264.7 | |
|---|---|---|---|---|---|---|---|
| Apamin | −0.497 | −0.490 | −0.409 | −0.430 | −0.483 | −0.053 | −0.560 * |
| PLA2 | −0.412 | −0.443 | −0.391 | −0.555 * | −0.590 * | −0.382 | −0.652 ** |
| Melittin | −0.311 | −0.302 | −0.284 | −0.214 | −0.205 | 0.154 | −0.161 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Mehdi, I.; Falcão, S.I.; Harandou, M.; Boujraf, S.; Calhelha, R.C.; Ferreira, I.C.F.R.; Anjos, O.; Campos, M.G.; Vilas-Boas, M. Chemical, Cytotoxic, and Anti-Inflammatory Assessment of Honey Bee Venom from Apis mellifera intermissa. Antibiotics 2021, 10, 1514. https://doi.org/10.3390/antibiotics10121514
El Mehdi I, Falcão SI, Harandou M, Boujraf S, Calhelha RC, Ferreira ICFR, Anjos O, Campos MG, Vilas-Boas M. Chemical, Cytotoxic, and Anti-Inflammatory Assessment of Honey Bee Venom from Apis mellifera intermissa. Antibiotics. 2021; 10(12):1514. https://doi.org/10.3390/antibiotics10121514
Chicago/Turabian StyleEl Mehdi, Iouraouine, Soraia I. Falcão, Mustapha Harandou, Saïd Boujraf, Ricardo C. Calhelha, Isabel C. F. R. Ferreira, Ofélia Anjos, Maria G. Campos, and Miguel Vilas-Boas. 2021. "Chemical, Cytotoxic, and Anti-Inflammatory Assessment of Honey Bee Venom from Apis mellifera intermissa" Antibiotics 10, no. 12: 1514. https://doi.org/10.3390/antibiotics10121514
APA StyleEl Mehdi, I., Falcão, S. I., Harandou, M., Boujraf, S., Calhelha, R. C., Ferreira, I. C. F. R., Anjos, O., Campos, M. G., & Vilas-Boas, M. (2021). Chemical, Cytotoxic, and Anti-Inflammatory Assessment of Honey Bee Venom from Apis mellifera intermissa. Antibiotics, 10(12), 1514. https://doi.org/10.3390/antibiotics10121514