
Presence of the Extended-Spectrum-β-Lactamase and Plasmid-Mediated AmpC-Encoding Genes in Escherichia coli from Companion Animals—A Study from a University-Based Veterinary Hospital in Taipei, Taiwan


Abstract
:1. Introduction
2. Results
2.1. Occurrence of the ESBL/pAmpC-Producing E. coli and Their bla Genotypes
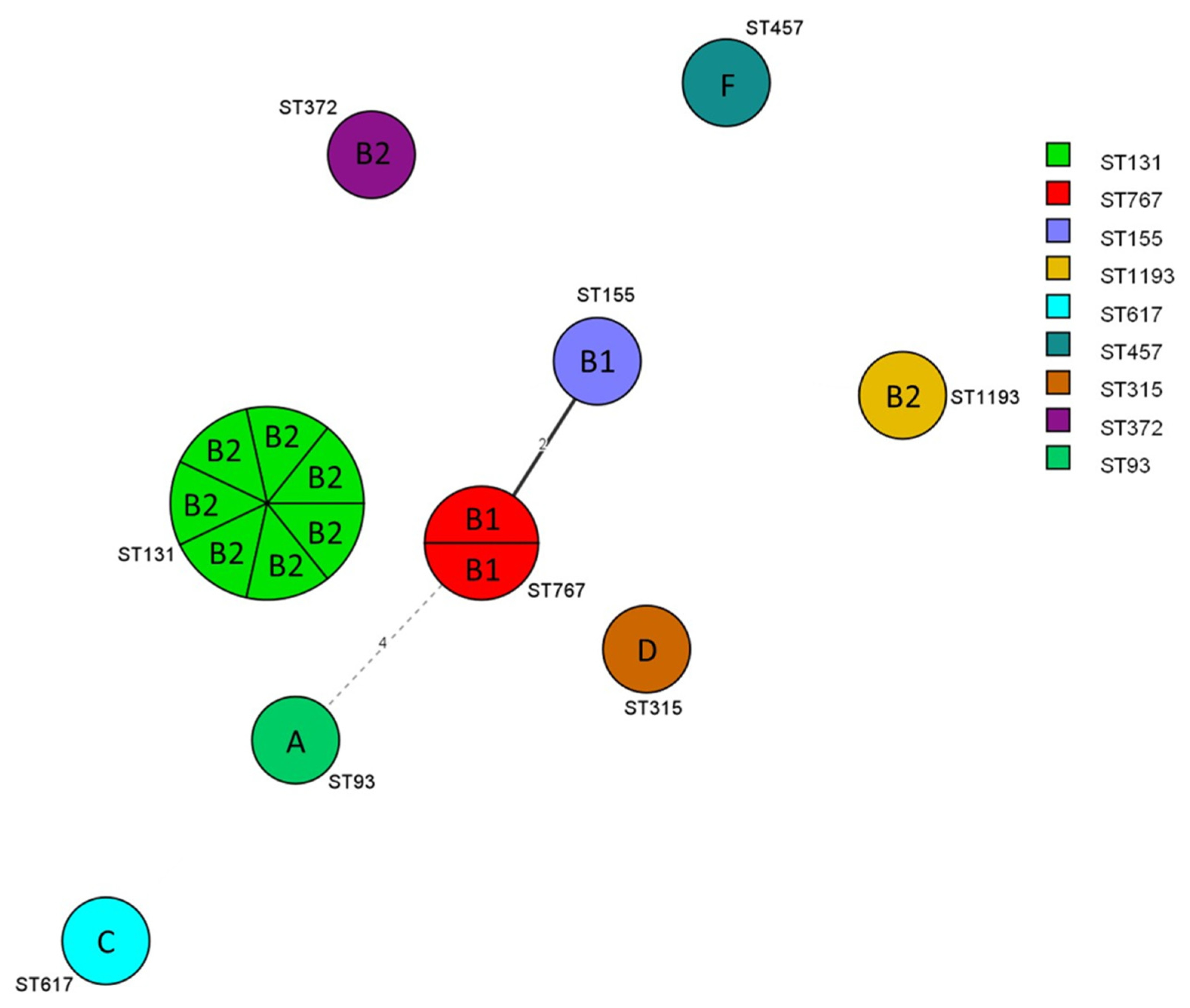
2.2. Phylogenetic Grouping, Genotyping, and Phylogenetic Analysis
2.3. E. coli ST131 O25b Detection
2.4. Conjugation Test
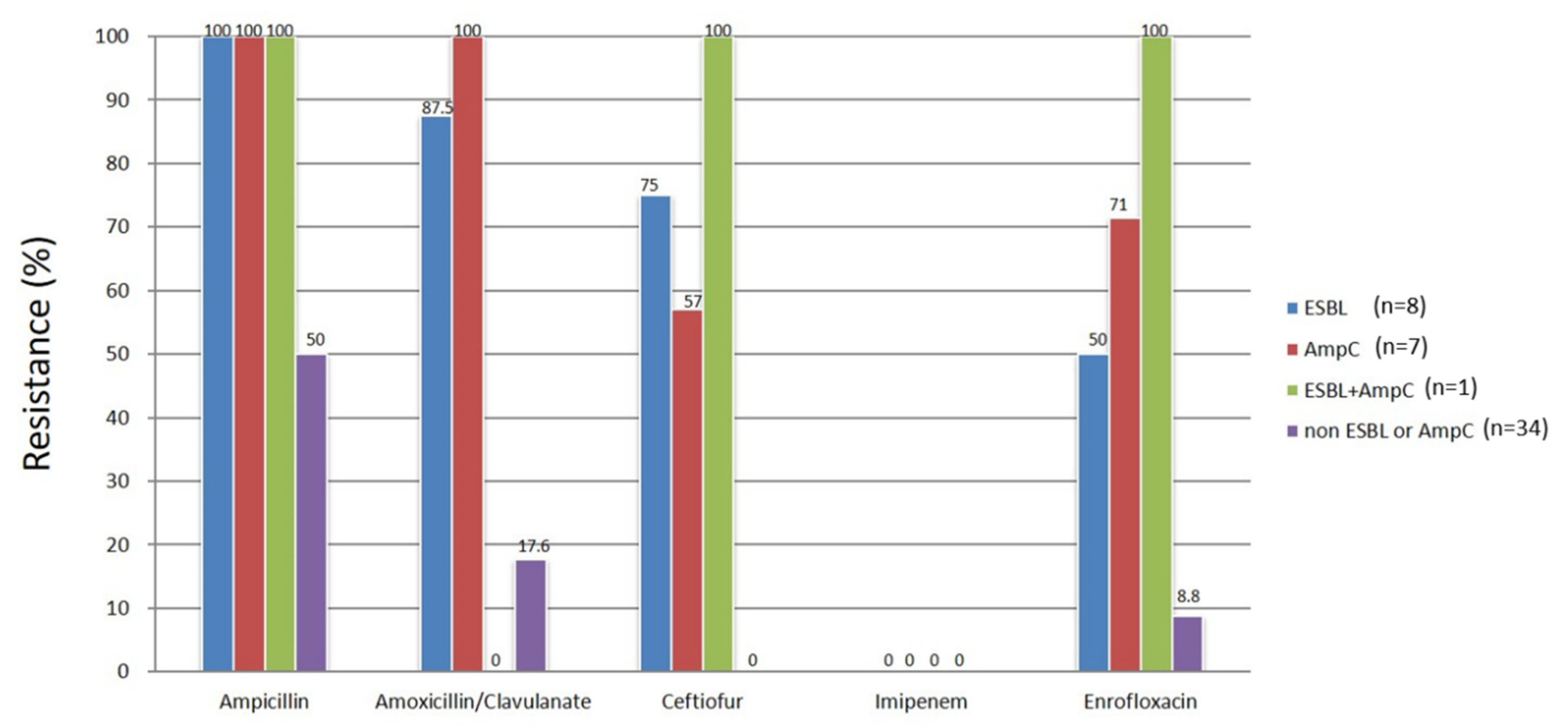
2.5. Antimicrobial Susceptibility Test
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. ESBL Screening and Phenotype Confirmation
4.3. Phylogenetic Grouping
4.4. Genotyping and Phylogenetic Analysis
4.5. E. coli ST131 O25b Detection
4.6. Conjugation Test
4.7. Antimicrobial Susceptibility Test
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaishnavi, C. Translocation of gut flora and its role in sepsis. Indian J. Med. Microbiol. 2013, 31, 334–342. [Google Scholar] [CrossRef] [PubMed]
- WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR). Critically Important Antimicrobials for Human Medicine, 5th ed.; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Bush, K. Past and present perspectives on beta-Lactamases. Antimicrob. Agents Chemother. 2018, 62, 10. [Google Scholar] [CrossRef] [Green Version]
- Philippon, A.; Arlet, G.; Jacoby, G.A. Plasmid-determined AmpC-type beta-lactamases. Antimicrob. Agents Chemother. 2002, 46, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honore, N.; Nicolas, M.H.; Cole, S.T. Inducible cephalosporinase production in clinical isolates of Enterobacter cloacae is controlled by a regulatory gene that has been deleted from Escherichia coli. EMBO J. 1986, 5, 3709–3714. [Google Scholar] [CrossRef]
- Papanicolaou, G.A.; Medeiros, A.A.; Jacoby, G.A. Novel plasmid-mediated beta-lactamase (MIR-1) conferring resistance to oxyimino- and alpha-methoxy beta-lactams in clinical isolates of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 1990, 34, 2200–2209. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.H.; Kuan, N.L.; Yeh, K.S. Characteristics of extended-spectrum beta-lactamase-producing Escherichia coli from dogs and cats admitted to a veterinary teaching hospital in Taipei, Taiwan from 2014 to 2017. Front. Vet. Sci. 2020, 7, 395. [Google Scholar] [CrossRef]
- Salgado-Caxito, M.; Benavides, J.A.; Adell, A.D.; Paes, A.C.; Moreno-Switt, A.I. Global prevalence and molecular characterization of extended-spectrum beta-lactamase producing-Escherichia coli in dogs and cats—A scoping review and meta-analysis. One Health 2021, 12, 100236. [Google Scholar] [CrossRef]
- McLain, J.E.; Cytryn, E.; Durso, L.M.; Young, S. Culture-based methods for detection of antibiotic resistance in agroecosystems: Advantages, challenges, and gaps in knowledge. J. Environ. Qual. 2016, 45, 432–440. [Google Scholar] [CrossRef]
- Tsuyuki, Y.; Kurita, G.; Murata, Y.; Takahashi, T.; Veterinary Infection Control Association Sepsis Working Group. Bacteria isolated from companion animals in Japan (2014–2016) by blood culture. J. Infect. Chemother. 2018, 24, 583–587. [Google Scholar] [CrossRef]
- Sun, Y.; Zeng, Z.; Chen, S.; Ma, J.; He, L.; Liu, Y.; Deng, Y.; Lei, T.; Zhao, J.; Liu, J.H. High prevalence of bla(CTX-M) extended-spectrum beta-lactamase genes in Escherichia coli isolates from pets and emergence of CTX-M-64 in China. Clin. Microbiol. Infect. 2010, 16, 1475–1481. [Google Scholar] [CrossRef]
- Ejaz, H.; Younas, S.; Abosalif, K.O.A.; Junaid, K.; Alzahrani, B.; Alsrhan, A.; Abdalla, A.E.; Ullah, M.I.; Qamar, M.U.; Hamam, S.S.M. Molecular analysis of blaSHV, blaTEM, and blaCTX-M in extended-spectrum β-lactamase producing Enterobacteriaceae recovered from fecal specimens of animals. PLoS ONE. 2021, 16, e0245126. [Google Scholar] [CrossRef] [PubMed]
- Okeke, I.N.; Lamikanra, A.; Edelman, R. Socioeconomic and behavioral factors leading to acquired bacterial resistance to antibiotics in developing countries. Emerg. Infect. Dis. 1999, 5, 18–27. [Google Scholar] [CrossRef]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejas, D.; Fernandez Canigia, L.; Quinteros, M.; Giovanakis, M.; Vay, C.; Lascialandare, S.; Mutti, D.; Pagniez, G.; Almuzara, M.; Gutkind, G.; et al. Plasmid-encoded AmpC (pAmpC) in Enterobacteriaceae: Epidemiology of microorganisms and resistance markers. Rev. Argent. Microbiol. 2012, 44, 182–186. [Google Scholar]
- Rumi, M.V.; Mas, J.; Elena, A.; Cerdeira, L.; Munoz, M.E.; Lincopan, N.; Gentilini, E.R.; Di Conza, J.; Gutkind, G. Co-occurrence of clinically relevant beta-lactamases and MCR-1 encoding genes in Escherichia coli from companion animals in Argentina. Vet. Microbiol. 2019, 230, 228–234. [Google Scholar] [CrossRef]
- Huber, H.; Zweifel, C.; Wittenbrink, M.M.; Stephan, R. ESBL-producing uropathogenic Escherichia coli isolated from dogs and cats in Switzerland. Vet. Microbiol. 2013, 162, 992–996. [Google Scholar] [CrossRef]
- Dierikx, C.M.; van Duijkeren, E.; Schoormans, A.H.W.; van Essen-Zandbergen, A.; Veldman, K.; Kant, A.; Huijsdens, X.W.; van der Zwaluw, K.; Wagenaar, J.A.; Mevius, D.J. Occurrence and characteristics of extended-spectrum-β-lactamase and AmpC-producing clinical isolates derived from companion animals and horses. J. Antimicrob. Chemother. 2012, 67, 1368–1374. [Google Scholar] [CrossRef]
- Bogaerts, P.; Huang, T.D.; Bouchahrouf, W.; Bauraing, C.; Berhin, C.; El Garch, F.; Glupczynski, Y.; ComPath Study Group. Characterization of ESBL- and AmpC-producing Enterobacteriaceae from diseased companion animals in Europe. Microb. Drug. Resist. 2015, 21, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Maeyama, Y.; Taniguchi, Y.; Hayashi, W.; Ohsaki, Y.; Osaka, S.; Koide, S.; Tamai, K.; Nagano, Y.; Arakawa, Y.; Nagano, N. Prevalence of ESBL/AmpC genes and specific clones among the third-generation cephalosporin-resistant Enterobacteriaceae from canine and feline clinical specimens in Japan. Vet. Microbiol. 2018, 216, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaheen, B.W.; Nayak, R.; Foley, S.L.; Kweon, O.; Deck, J.; Park, M.; Rafii, F.; Boothe, D.M. Molecular characterization of resistance to extended-spectrum cephalosporins in clinical Escherichia coli isolates from companion animals in the United States. Antimicrob. Agents Chemother. 2011, 555, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.C.; Barbosa, A.V.; Arais, L.R.; Ribeiro, P.F.; Carneiro, V.C.; Cerqueira, A.M. Resistance patterns, ESBL genes, and genetic relatedness of Escherichia coli from dogs and owners. Braz. J. Microbiol. 2016, 47, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Teunis, P.F.M.; Evers, E.G.; Hengeveld, P.D.; Dierikx, C.M.; Wielders, C.C.H.; van Duijkeren, E. Time to acquire and lose carriership of ESBL/pAmpC producing E. coli in humans in the Netherlands. PLoS ONE 2018, 13, e0193834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathers, A.J.; Peirano, G.; Pitout, J.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [Green Version]
- Pomba, C.; da Fonseca, J.D.; Baptista, B.C.; Correia, J.D.; Martinez-Martinez, L. Detection of the pandemic O25-ST131 human virulent Escherichia coli CTX-M-15-producing clone harboring the qnrB2 and aac(6′)-Ib-cr genes in a dog. Antimicrob. Agents Chemother. 2009, 53, 327–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofte, D.; Maciuca, I.E.; Kemmett, K.; Wattret, A.; Williams, N.J. Detection of the human pandemic Escherichia coli B2-O25b-ST131 in UK dogs. Vet. Rec. 2014, 17, 4352. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokova, V.; Radey, M.; Chattopadhyay, S.; Larson, L.; Weaver, J.L.; Kisiela, D.; Sokurenko, E.V. Pandemic fluoroquinolone resistant Escherichia coli clone ST1193 emerged via simultaneous homologous recombinations in 11 gene loci. Proc. Natl. Acad. Sci. USA 2019, 116, 14740–14748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Johnston, B.D.; Porter, S.B.; Clabots, C.; Bender, T.L.; Thuras, P.; Trott, D.J.; Cobbold, R.; Mollinger, J.; Ferrieri, P.; et al. Rapid emergence, subsidence, and molecular detection of Escherichia coli sequence type 1193-fimH64, a new disseminated multidrug-resistant commensal and extraintestinal pathogen. J. Clin. Microbiol. 2019, 57, e01664-18. [Google Scholar] [CrossRef] [Green Version]
- Platell, J.L.; Trott, D.J.; Johnson, J.R.; Heisig, P.; Heisig, A.; Clabots, C.R.; Johnston, B.; Cobbold, R.N. Prominence of an O75 clonal group (clonal complex 14) among non-ST131 fluoroquinolone-resistant Escherichia coli causing extraintestinal infections in humans and dogs in Australia. Antimicrob. Agents Chemother. 2012, 56, 3898–3904. [Google Scholar] [CrossRef] [Green Version]
- Valenza, G.; Werner, M.; Eisenberger, D.; Nickel, S.; Lehner-Reindl, V.; Holler, C.; Bogdan, C. First report of the new emerging global clone ST1193 among clinical isolates of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli from Germany. J. Glob. Antimicrob. Resist. 2019, 17, 305–308. [Google Scholar] [CrossRef]
- Kim, Y.; Oh, T.; Nam, Y.S.; Cho, S.Y.; Lee, H.J. Prevalence of ST131 and ST1193 among bloodstream isolates of Escherichia coli not susceptible to ciprofloxacin in a tertiary care university hospital in Korea, 2013–2014. Clin. Lab. 2017, 63, 1541–1543. [Google Scholar] [CrossRef]
- Wu, J.; Lan, F.; Lu, Y.; He, Q.; Li, B. Molecular characteristics of ST1193 clone among phylogenetic group B2 non-ST131 fluoroquinolone-resistant Escherichia coli. Front. Microbiol. 2017, 8, 2294. [Google Scholar] [CrossRef] [Green Version]
- Tchesnokova, V.L.; Rechkina, E.; Larson, L.; Ferrier, K.; Weaver, J.L.; Schroeder, D.W.; She, R.; Butler-Wu, S.M.; Aguero-Rosenfeld, M.E.; Zerr, D.; et al. Rapid and extensive expansion in the United States of a new multidrug-resistant Escherichia coli clonal group, sequence type 1193. Clin. Infect. Dis. 2019, 68, 334–337. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, X.; Zheng, S.; Yu, F.; Kong, H.; Yang, Q.; Cui, D.; Chen, N.; Lou, B.; Li, X.; et al. Serotypes, genotypes and antimicrobial resistance patterns of human diarrhoeagenic Escherichia coli isolates circulating in southeastern China. Clin. Microbiol. Infect. 2014, 20, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar]
- Vogt, D.; Overesch, G.; Endimiani, A.; Collaud, A.; Thomann, A.; Perreten, V. Occurrence and genetic characteristics of third-generation cephalosporin-resistant Escherichia coli in Swiss retail meat. Microb. Drug Resist. 2014, 20, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzouvelekis, L.S.; Tzelepi, E.; Mentis, A.F.; Tsakris, A. Identification of a novel plasmid-mediated beta-lactamase with chromosomal cephalosporinase characteristics from Klebsiella pneumoniae. J. Antimicrob. Chemother. 1993, 31, 645–654. [Google Scholar] [CrossRef]
- Wu, S.W.; Dornbusch, K.; Kronvall, G.; Norgren, M. Characterization and nucleotide sequence of a Klebsiella oxytoca cryptic plasmid encoding a CMY-type beta-lactamase: Confirmation that the plasmid-mediated cephamycinase originated from the Citrobacter freundii AmpC beta-lactamase. Antimicrob. Agents Chemother. 1999, 43, 1350–1357. [Google Scholar] [CrossRef] [Green Version]
- Barker, C.W.; Zhang, W.; Sanchez, S.; Budsberg, S.C.; Boudinot, F.D.; Stevenson, M.A.M. Pharmacokinetics of imipenem in dogs. Am. J. Vet. Res. 2003, 64, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Han, J.I.; Jeong, S.H. First detection of New Delhi metallo-beta-lactamase-5-producing Escherichia coli from companion animals in Korea. Microb. Drug. Resist. 2019, 25, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.D.; Peak, L.; Tyson, G.H.; Reimschuessel, R.; Ceric, O.; Rankin, S.C. New Delhi metallo-beta-lactamase-5-producing Escherichia coli in companion animals, United States. Emerg. Infect. Dis. 2020, 26, 381–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornsey, M.; Phee, L.; Woodford, N.; Turton, J.; Meunier, D.; Thomas, C.; Wareham, D.W. Evaluation of three selective chromogenic media, CHROMagar ESBL, CHROMagar CTX-M and CHROMagar KPC, for the detection of Klebsiella pneumoniae producing OXA-48 carbapenemase. J. Clin. Pathol. 2013, 66, 348–350. [Google Scholar] [CrossRef] [Green Version]
- CLSI. M-100. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, B.W.; Oyarzabal, O.A.; Boothe, D.M. The role of class 1 and 2 integrons in mediating antimicrobial resistance among canine and feline clinical E. coli isolates from the US. Vet. Microbiol. 2010, 144, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, J.G. Nucleotide-sequence of ampicillin resistance gene of Escherichia coli plasmid pBR322. Proc. Natl. Acad. Sci. USA 1978, 75, 3737–3741. [Google Scholar] [CrossRef] [Green Version]
- Chia, J.H.; Chu, C.; Su, L.H.; Chiu, C.H.; Kuo, A.J.; Sun, C.F.; Wu, T.L. Development of a multiplex PCR and SHV melting-curve mutation detection system for detection of some SHV and CTX-M beta-lactamases of Escherichia coli, Klebsiella pneumoniae, and Enterobacter cloacae in Taiwan. J. Clin. Microbiol. 2005, 43, 4486–4491. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Ji, S.; Chen, Y.; Zhou, W.; Wei, Z.; Li, L.; Ma, Y. Resistance of strains producing extended-spectrum beta-lactamases and genotype distribution in China. J. Infect. 2007, 54, 53–57. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zheng, B.; Zhao, L.; Wei, Z.; Ji, J.; Li, L.; Xiao, Y. Nationwide high prevalence of CTX-M and an increase of CTX-M-55 in Escherichia coli isolated from patients with community-onset infections in Chinese county hospitals. BMC Infect. Dis. 2014, 14, 659. [Google Scholar] [CrossRef]
- Javier Pérez-Pérez, F.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Characterization of an anonymous molecular marker strongly linked to Escherichia coli strains causing neonatal meningitis. J. Clin. Microbiol. 2004, 42, 1770–1772. [Google Scholar] [CrossRef] [Green Version]
- Lescat, M.; Clermont, O.; Woerther, P.L.; Glodt, J.; Dion, S.; Skurnik, D.; Djossou, F.; Dupont, C.; Perroz, G.; Picard, B.; et al. Commensal Escherichia coli strains in Guiana reveal a high genetic diversity with host-dependant population structure. Environ. Microbiol. Rep. 2013, 5, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Lescat, M.; O’Brien, C.L.; Gordon, D.M.; Tenaillon, O.; Denamur, E. Evidence for a human-specific Escherichia coli clone. Environ. Microbiol. 2008, 10, 1000–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppe, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, M.D.; Nam, H.M.; Jang, G.C.; Kim, S.R.; Chae, M.H.; Jung, S.C.; Byun, J.W.; Park, Y.H.; Lim, S.K. Molecular characterization of extended-spectrum-beta-lactamase-producing and plasmid-mediated AmpC beta-lactamase-producing Escherichia coli isolated from stray dogs in South Korea. Antimicrob. Agents Chemother. 2012, 56, 2705–2712. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Case No. | Species | Source | Phylogroup | ST Type | bla Genes |
|---|---|---|---|---|---|
| 001 | cat | urine | B2 | ST131 | blaTEM-215 + blaCTX-M-235 (ESBL) |
| 002 | dog | urine | A | ST93 | blaCTX-M-55 + blaCMY-171 (ESBL + pAmpC) |
| 004 | cat | ascites | B2 | ST131 | blaCTX-M-235 (ESBL) |
| 008 | dog | urine | B1 | ST155 | blaCMY-171 (pAmpC) |
| 010 | dog | urine | D | ST315 | blaCMY-171 (pAmpC) |
| 011 | cat | urine | B2 | ST131 | blaTEM-215 + blaCTX-M-55 + blaCTX-M-235 (ESBL) |
| 025 | dog | nasal discharge | B1 | ST767 | blaTEM-243 + blaCMY-171 (pAmpC) |
| 031 | dog | urine | B2 | ST131 | blaTEM-215 + blaCTX-M-211 + blaCTX-M-235 (ESBL) |
| 032 | cat | pus | C | ST617 | blaCMY-171 (pAmpC) |
| 034 | dog | urine | B2 | ST131 | blaCTX-M-238 (ESBL) |
| 038 | dog | urine | F | ST457 | blaCMY-171 (pAmpC) |
| 040 | dog | urine | B2 | ST131 | blaCTX-M-235 (ESBL) |
| 042 | dog | urine | B2 | ST131 | blaCTX-M-238 (ESBL) |
| 049 | dog | nasal discharge | B1 | ST767 | blaCMY-171 (pAmpC) |
| 050 | dog | oral mass | B2 | ST1193 | blaTEM-215 + blaCTX-M-199 (ESBL) |
| 051 | dog | urine | B2 | ST372 | blaCMY-171 (pAmpC) |
| Case No. | bla Genes in the Donor Strain | bla Genes in the Transconjugant Strain |
|---|---|---|
| 001 | blaTEM-215, blaCTX-M-235 | blaTEM-215, blaCTX-M-235 |
| 002 | blaCTX-M-55, blaCMY-171 | blaCTX-M-55, blaCMY-171 |
| 004 | blaCTX-M-235 | blaCTX-M-235 |
| 008 | blaCMY-171 | blaCMY-171 |
| 010 | blaCMY-171 | blaCMY-171 |
| 011 | blaTEM-215, blaCTX-M-55, blaCTX-M-235 | blaTEM-215, blaCTX-M-235 |
| 025 | blaTEM-243, blaCMY-171 | blaCMY-171 |
| 031 | blaTEM-215, blaCTX-M-211, blaCTX-M-235 | blaTEM-215, blaCTX-M-211, blaCTX-M-235 |
| 032 | blaCMY-171 | blaCMY-171 |
| 034 | blaCTX-M-238 | blaCTX-M-238 |
| 038 | blaCMY-171 | blaCMY-171 |
| 040 | blaCTX-M-235 | blaCTX-M-235 |
| 042 | blaCTX-M-238 | blaCTX-M-238 |
| 049 | blaCMY-171 | -- † |
| 050 | blaTEM-215, blaCTX-M-199 | blaTEM-215, blaCTX-M-199 |
| 051 | blaCMY-171 | blaCMY-171 |
| PCR Target | Primer | Sequences (5′-3′) | Annealing Tm (°C) | Predicted PCR Size (bp) | Reference |
|---|---|---|---|---|---|
| blaTEM | TEM-F | TCGGGGAAATGTGCGCG | 55 | 972 | [47] |
| TEM-R | TGCTTAATCATGAGGCACC | ||||
| blaSHV | SHV-F | GCCTTTATCGGCCCTCATCAA | 54 | 819 | [48] |
| SHV-R | TCCCGCAGATAAATCACCACAATG | ||||
| blaCTX-M-1-group | CTX-M-1-F | CCCATGGTTAAAAAATCACTGC | 54 | 942 | [49] |
| CTX-M-1-R | CAGCGCTTTTGCCGTCTAAG | ||||
| blaCTX-M-2-group | CTX-M-2-F | CGACGCTACCCCTGCTATT | 52 | 552 | [50] |
| CTX-M-2-R | CCAGCGTCAGATTTTTCAGG | ||||
| blaCTX-M-8-group | CTX-M-8-F | TCGCGTTAAGCGGATGATGC | 52 | 666 | [50] |
| CTX-M-8-R | AACCCACGATGTGGGTAGC | ||||
| blaCTX-M-9-group | CTX-M-9-F | ATGGTGACAAAGAGAGTGCAAC | 55 | 876 | [51] |
| CTX-M-9-R | TTACAGCCCTTCGGCGATGATT | ||||
| blaCTX-M-25-group | CTX-M-25-F | GCACGATGACATTCGGG | 52 | 327 | [50] |
| CTX-M-25-R | AACCCACGATGTGGGTAGC | ||||
| blapAmpC | CIT-M-F | TGGCCAGAACTGACAGGCAAA | 64 | 462 | [52] |
| CIT-M-R | TTTCTCCTGAACGTCGCTGGC | ||||
| blapAmpC | MOX-M-F | GCTGCTCAAGGAGCACAGGAT | 64 | 520 | [52] |
| MOX-M-R | CACATTGACATAGGTGTGGTGC | ||||
| blapAmpC | DHA-M-F | AACTTTCACAGCTGTGCTGGGT | 64 | 405 | [52] |
| DHA-M-R | CCGTACGCATACTGGCTTTGC | ||||
| blapAmpC | CMY-F | ATGATGAAAAAATCGTTATGCT | 64 | 1146 | [52] |
| CMY-R | TTATTGCAGCTTTTCAAGAATGCG | ||||
| chuA | chuA.1b | ATGGTACCGGACGAACCAAC | 59 | 288 | [45,53] |
| chuA.2 | TGCCGCCAGTACCAAAGACA | ||||
| yjaA | yjaA.1b | CAAACGTGAAGTGTCAGGAG | 59 | 211 | [45] |
| yjaA.2b | AATGCGTTCCTCAACCTGTG | ||||
| TspE4.C2 | TspE4C2.1b | CACTATTCGTAAGGTCATCC | 59 | 152 | [45] |
| TspE4C2.2b | AGTTTATCGCTGCGGGTCGC | ||||
| arpA | AceK.f | AACGCTATTCGCCAGCTTGC | 59 | 400 | [45,54] |
| ArpA1.r | TCTCCCCATACCGTACGCTA | ||||
| arpA | ArpAgpE.f | GATTCCATCTTGTCAAAATATGCC | 57 | 301 | [55] |
| ArpAgpE.r | GAAAAGAAAAAGAATTCCCAAGAG | ||||
| trpA | trpAgpC.1 | AGTTTTATGCCCAGTGCGAG | 59 | 219 | [55] |
| trpAgpC.2 | TCTGCGCCGGTCACGCCC | ||||
| trpA | trpBA.f | CGGCGATAAAGACATCTTCAC | 59 | 489 | [56] |
| trpBA.r | GCAACGCGGCCTGGCGGAAG | ||||
| pabB | O25pabBspe.F | TCCAGCAGGTGCTGGATCGT | 65 | 347 | [57] |
| O25pabBspe.R | GCGAAATTTTTCGCCGTACTGT | ||||
| trpA | trpA.F | GCTACGAATCTCTGTTTGCC | 65 | 427 | [57] |
| trpA2.R | GCAACGCGGCCTGGCGGAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.-L.; Kuan, N.-L.; Yeh, K.-S. Presence of the Extended-Spectrum-β-Lactamase and Plasmid-Mediated AmpC-Encoding Genes in Escherichia coli from Companion Animals—A Study from a University-Based Veterinary Hospital in Taipei, Taiwan. Antibiotics 2021, 10, 1536. https://doi.org/10.3390/antibiotics10121536
Liu F-L, Kuan N-L, Yeh K-S. Presence of the Extended-Spectrum-β-Lactamase and Plasmid-Mediated AmpC-Encoding Genes in Escherichia coli from Companion Animals—A Study from a University-Based Veterinary Hospital in Taipei, Taiwan. Antibiotics. 2021; 10(12):1536. https://doi.org/10.3390/antibiotics10121536
Chicago/Turabian StyleLiu, Fang-Ling, Nan-Ling Kuan, and Kuang-Sheng Yeh. 2021. "Presence of the Extended-Spectrum-β-Lactamase and Plasmid-Mediated AmpC-Encoding Genes in Escherichia coli from Companion Animals—A Study from a University-Based Veterinary Hospital in Taipei, Taiwan" Antibiotics 10, no. 12: 1536. https://doi.org/10.3390/antibiotics10121536
APA StyleLiu, F. -L., Kuan, N. -L., & Yeh, K. -S. (2021). Presence of the Extended-Spectrum-β-Lactamase and Plasmid-Mediated AmpC-Encoding Genes in Escherichia coli from Companion Animals—A Study from a University-Based Veterinary Hospital in Taipei, Taiwan. Antibiotics, 10(12), 1536. https://doi.org/10.3390/antibiotics10121536