
Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
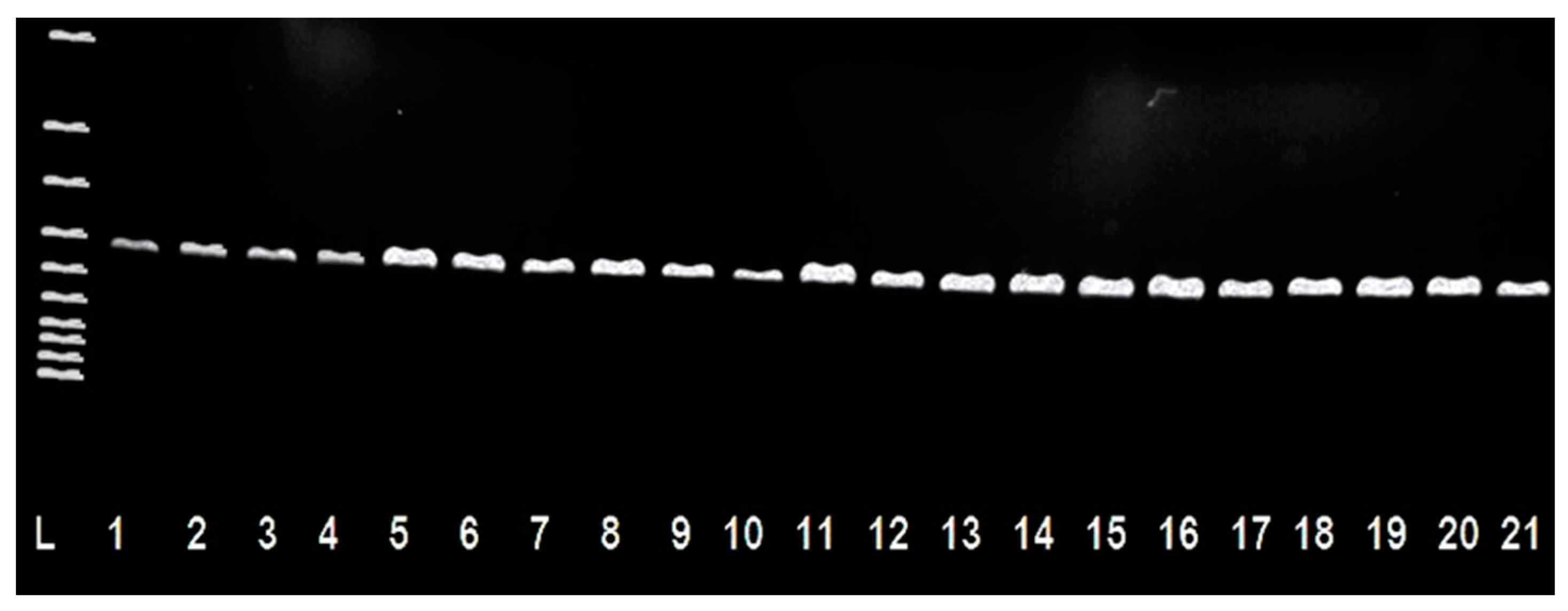
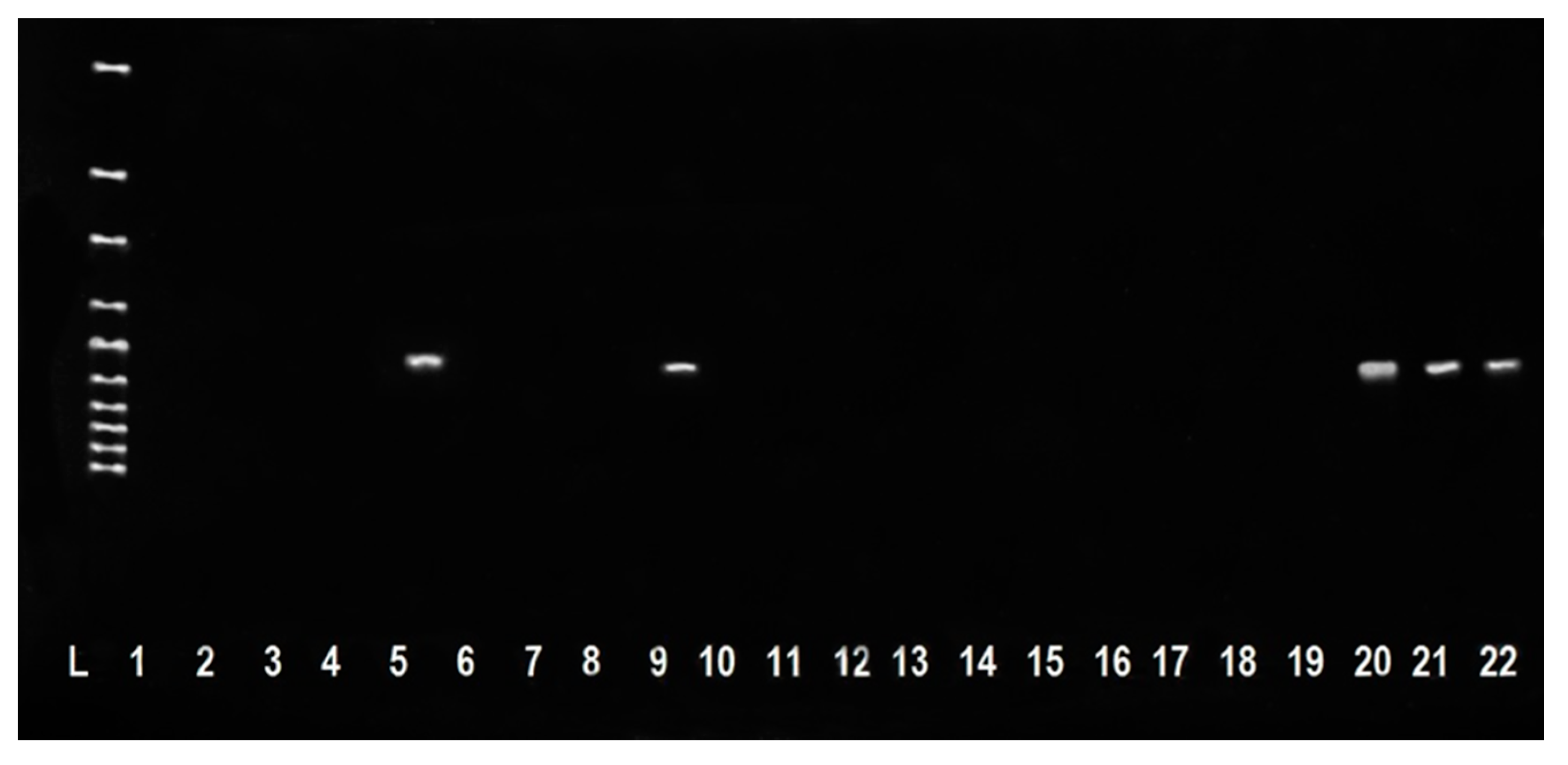
2. Results
3. Discussion
4. Materials and Methods
4.1. Isolation of Staphylococci
4.2. Identification of Isolates
4.3. Detection of Antimicrobial Resistance
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cannas, A.; Tedeschi, L.O.; Atzori, A.S.; Lunesu, M.F. How can nutrition models increase the production efficiency of sheep and goat operations? Anim. Front. 2019, 9, 33–44. [Google Scholar] [CrossRef]
- FAOSTAT. Compare Data Food and Agriculture Organization of the United Nations. 2020. Available online: http://faostat.fao.org/ (accessed on 14 April 2021).
- Balthazar, C.F.; Pimentel, T.C.; Ferrão, L.L.; Almada, C.N.; Santillo, A.; Albenzio, M.; Mollakhalili, N.; Mortazavian, A.M.; Nascimento, J.S.; Silva, M.C.; et al. Sheep milk: Physicochemical characteristics and relevance for functional food development. Compr. Rev. Food Sci. Food Saf. 2017, 16, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Balthazar, C.F.; Santillo, A.; Guimarães, J.T.; Capozzi, V.; Russo, P.; Caroprese, M.; Marino, R.; Esmerino, E.A.; Raices, R.S.L.; Silva, M.C.; et al. Novel milk–juice beverage with fermented sheep milk and strawberry (Fragaria × ananassa): Nutritional and functional characterization. J. Dairy Sci. 2019, 102, 10724–10736. [Google Scholar] [CrossRef] [Green Version]
- Gelasakis, A.I.; Mavrogianni, V.S.; Petridis, I.G.; Vasileiou, N.G.C.; Fthenakis, G.C. Mastitis in sheep—The last 10 years and the future of research. Vet. Microbiol. 2015, 185, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, D.B.; Naushad, S.; Naqvi, S.A.; Condas, L.A.; Saini, V.; Kastelic, J.P.; Luby, C.; De Buck, J.; Barkema, H.W. Prevalence and genetic basis of antimicrobial resistance in non-aureus staphylococci isolated from Canadian dairy herds. Front. Microbiol. 2018, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Vasileiou, N.G.C.; Cripps, P.J.; Ioannidi, K.S.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Orfanou, D.C.; Politis, A.P.; Calvo Gonzalez-Valerio, T.; Argyros, S.; et al. Extensive countrywide field investigation of subclinical mastitis in sheep in Greece. J. Dairy Sci. 2018, 101, 7297–7310. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Zhao, H.; Nobrega, D.B.; Cobo, E.R.; Han, B.; Zhao, Z.; Li, S.; Li, M.; Barkema, H.W.; Gao, J. Molecular epidemiology and distribution of antimicrobial resistance genes of Staphylococcus species isolated from Chinese dairy cows with clinical mastitis. J. Dairy. Sci. 2019, 102, 1571–1583. [Google Scholar] [CrossRef] [Green Version]
- Fišarová, L.; Pantůček, R.; Botka, T.; Doškař, J. Variability of resistance plasmids in coagulase-negative staphylococci and their importance as a reservoir of antimicrobial resistance. Res. Microbiol. 2019, 170, 105–111. [Google Scholar] [CrossRef]
- Wendlandt, S.; Feßler, A.T.; Monecke, S.; Ehricht, R.; Schwarz, S.; Kadlec, K. The diversity of antimicrobial resistance genes among staphylococci of animal origin. Int. J. Med. Microbiol. 2013, 303, 338–349. [Google Scholar] [CrossRef]
- Marogna, G.; Rolesu, S.; Lollai, S.; Tola, S.; Leori, G. Clinical findings in sheep farms affected by recurrent bacterial mastitis. Small Rumin. Res. 2010, 88, 119–125. [Google Scholar] [CrossRef]
- Onni, T.; Vidili, A.; Bandino, E.; Marogna, G.; Schianchi, S.; Tola, S. Identification of coagulase-negative staphylococci isolated from caprine milk samples by PCR-RFLP of groEL gene. Small Rumin. Res. 2012, 104, 185–190. [Google Scholar] [CrossRef]
- Vanderhaeghen, W.; Piepers, S.; Leroy, F.; Van Coillie, E.; Haesebrouck, F.; De Vliegher, S. Identification, typing, ecology and epidemiology of coagulase negative staphylococci associated with ruminants. Vet. J. 2015, 203, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Martins, K.B.; Faccioli, P.Y.; Bonesso, M.F.; Fernandes, S.; Oliveira, A.A.; Dantas, A.; Zafalon, L.F.; Maria de Lourdes, R.S. Characteristics of resistance and virulence factors in different species of coagulase-negative staphylococci isolated from milk of healthy sheep and animals with subclinical mastitis. Int. J. Dairy Sci. 2017, 100, 2184–2195. [Google Scholar] [CrossRef] [Green Version]
- Von Eiff, C.; Peters, G.; Heilmann, C. Pathogenesis of infections due to coagulasenegative staphylococci. Lancet Infect. Dis. 2002, 2, 677–685. [Google Scholar] [CrossRef]
- Onni, T.; Sanna, G.; Larsen, J.; Tola, S. Antimicrobial susceptibilities and population structure of Staphylococcus epidermidis associated with ovine mastitis. Vet. Microbiol. 2011, 148, 45–50. [Google Scholar] [CrossRef]
- Turchi, B.; Bertelloni, F.; Marzoli, F.; Cerri, D.; Tola, S.; Azara, E.; Longheu, C.M.; Tassi, R.; Schiavo, M.; Cilia, G.; et al. Coagulase negative staphylococci from ovine milk: Genotypic and phenotypic characterization of susceptibility to antibiotics, disinfectants and biofilm production. Small Rumin. Res. 2020, 183, 106030. [Google Scholar] [CrossRef]
- Matsuhashi, M.; Song, M.D.; Ishino, F.; Wachi, M.; Doi, M.; Inoue, M.; Ubukata, K.; Yamashita, N.; Konno, M. Molecular cloning of the gene of a penicillin-binding protein supposed to cause high resistance to beta-lactam antibiotics in Staphylococcus aureus. J. Bacteriol. 1986, 167, 975–980. [Google Scholar] [CrossRef] [Green Version]
- Saber, H.; Jasni, A.S.; Jamaluddin, T.Z.M.T.; Ibrahim, R. A review of staphylococcal cassette chromosome mec (SCCmec) types in coagulase-negative staphylococci (CoNS) species. Malays. J. Med. Sci. 2017, 24, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Katayama, Y.; Ito, T.; Hiramatsu, K. Genetic organization of the chromosome region surrounding mecA in clinical staphylococcal strains: Role of IS431-mediated mecI deletion in expression of resistance in mecA-carrying, low-level methicillin-resistant Staphylococcus haemolyticus. Antimicrob. Agents Chemother. 2001, 45, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballhausen, B.; Kriegeskorte, A.; Schleimer, N.; Peters, G.; Becker, K. The mecA homolog mecC confers resistance against β-lactams in Staphylococcus aureus irrespective of the genetic strain background. Antimicrob. Agents Chemother. 2014, 58, 3791–3798. [Google Scholar] [CrossRef] [Green Version]
- Juhász-Kaszanyitzky, É.; Jánosi, S.; Somogyi, P.; Dán, Á.; Van Bloois, L.V.; Van Duijkeren, E.; Wagenaar, J.A. MRSA Transmission between Cows and Humans. Emerg. Infect. Dis. 2007, 13, 630–632. [Google Scholar] [CrossRef]
- Fisher, E.A.; Paterson, G.K. Prevalence and characterization of methicillin-resistant staphylococci from bovine bulk tank milk in England and Wales. J. Glob. Antimicrob. Resist. 2020, 22, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Waller, K.P.; Aspan, A.; Nyman, A.; Persson, Y.; Andersson, U.G. CNS species and antimicrobial resistance in clinical and subclinical bovine mastitis. Vet. Microbiol. 2011, 152, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eid, R.; El Jakee, J.; Rashidy, A.; Asfour, H.; Omara, S.; Kandil, M.M.I.; Mahmood, Z.; Hahne, J.; Seida, A.A. Potential antimicrobial activities of probiotic Lactobacillus strains isolated from raw milk. J. Prob. Health 2016, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pantoflickova, D.; Corthésy-Theulaz, I.; Stolte, M.; Isler, P.; Rochat, F.; Enslen, M.; Blum, A.L. Favourable effect of regular intake of fermented milk containing Lactobacillus johnsonii on Helicobacter pylori associated gastritis. Aliment. Pharmacol. Ther. 2003, 18, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Bencúrová, E.; Bhide, M.; Dolinská, S.; Hreško, S.; Mlynárčik, P.; Mucha, R.; Pulzová, L. (Eds.) New Trends in the Use of Bioinformatics Analysis in Genomics and Proteomics (Nové Trendy vo Využívaní Bioinformatických Analýz v Genomike a Proteomike), 1st ed.; University of Veterinary Medicine and Pharmacy in Košice: Košice, Slovakia, 2013; pp. 23–24. [Google Scholar]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization-time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [Green Version]
- Okuma, K.; Iwakawa, K.; Turnidge, J.D.; Grubb, W.B.; Bell, J.M.; O’Brien, F.G.; Coombs, G.W.; Pearman, J.W.; Tenover, F.C.; Kapi, M.; et al. Dissemination of new methicillin-resistant Staphylococcus aureus clones in the community. J. Clin. Microbiol. 2002, 11, 4289–4294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickerson, S.C.; Boddie, R.L. Effect of naturally occurring coagulase-negative staphylococcal infections on experimental challenge with major mastitis pathogens. J. Dairy Sci. 1994, 77, 2526–2536. [Google Scholar] [CrossRef]
- Alves, V.F.; Niño-Arias, F.C.; Pitondo-Silva, A.; Araújo Frazilio, D.; Gonçalves, L.O.; Toubas, L.C.; De Martinis, E.C.P. Molecular characterisation of Staphylococcus aureus from some artisanal Brazilian dairies. Int. Dairy J. 2018, 85, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Supré, K.; De Vliegher, S.; Sampimon, O.C.; Zadoks, R.N.; Vaneechoutte, M.; Baele, M.; De Graef, E.; Piepers, S.; Haesebrouck, F. Technical note: Use of transfer RNA-intergenic spacer PCR combined with capillary electrophoresis to identify coagulase-negative Staphylococcus species originating from bovine milk and teat apices. J. Dairy Sci. 2009, 92, 3204–3210. [Google Scholar] [CrossRef]
- Braem, G.; De Vliegher, S.; Supré, K.; Haesebrouck, F.; Leroy, F.; De Vuyst, L. (GTG) 5-PCR fingerprinting for the classification and identification of coagulase-negative Staphylococcus species from bovine milk and teat apices: A comparison of type strains and field isolates. Vet. Microbiol. 2011, 147, 67–74. [Google Scholar] [CrossRef]
- De Visscher, A.; Supré, K.; Haesebrouck, F.; Zadoks, R.N.; Piessens, V.; Van Coillie, E.; Piepers, S.; De Vliegher, S. Further evidence for the existence of environmental and host-associated species of coagulase-negative staphylococci in dairy cattle. Vet. Microbiol. 2014, 172, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Fry, P.R.; Middleton, J.R.; Dufour, S.; Perry, J.; Scholl, D.; Dohoo, I. Association of coagulase-negative staphylococcal species, mammary quarter milk somatic cell count, and persistence of intramammary infection in dairy cattle. J. Dairy Sci. 2014, 97, 4876–4885. [Google Scholar] [CrossRef]
- Zigo, F.; Laktičová, K.V.; Vargová, M.M. The occurrence of bacterial agents causing mastitis in dairy sheep and their resistance to antibiotics. J. Med. Res. Health Sci. 2020, 3. [Google Scholar] [CrossRef]
- Virdis, S.; Scarano, C.; Cossu, F.; Spanu, V.; Spanu, C.; De Santis, E.P.L. Antibiotic resistance in Staphylococcus aureus and coagulase negative staphylococci isolated from goats with subclinical mastitis. Vet. Med. Int. 2010, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sharoud, W.M.; Spano, G. Diversity and enterotoxigenicity of Staphylococcus spp. associated with Domiati cheese. J. Food Protect. 2008, 71, 2567–2571. [Google Scholar] [CrossRef] [PubMed]
- Coton, E.; Desmonts, M.H.; Leroy, S.; Coton, M.; Jamet, E.; Christieans, S.; Donnio, P.Y.; Lebert, I.; Talon, R. Biodiversity of coagulase-negative staphylococci in French cheeses, dry fermented sausages, processing environments and clinical samples. Int. J. Food Microbiol. 2010, 137, 221–229. [Google Scholar] [CrossRef]
- Rahmdel, S.; Hosseinzadeh, S.; Shekarforoush, S.S.; Torriani, S.; Gatto, V.; Pashangeh, S. Safety hazards in bacteriocinogenic Staphylococcus strains isolated from goat and sheep milk. Microb. Pathog. 2018, 116, 100–108. [Google Scholar] [CrossRef]
- Yoon, Y.; Lee, S.; Choi, K.H. Microbial benefits and risks of raw milk cheese. Food Control 2016, 63, 201–215. [Google Scholar] [CrossRef]
- Seng, P.; Boushab, B.M.; Romain, F.; Gouriet, F.; Bruder, N.; Martin, C.; Papazian, L. Emerging role of Raoultella ornithinolytica in human infections: A series of cases and review of the literature. Int. J. Inf. Dis. 2016, 45, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Sampimon, O.C. Coagulase-Negative Staphylococci Mastitis in Dutch Dairy Herds. Ph.D. Thesis, Utrecht University Repository, Ultrecht, The Netherlands, 2009. [Google Scholar]
- Sawant, A.A.; Gillespie, B.E.; Oliver, S.P. Antimicrobial susceptibility of coagulase-negative Staphylococcus species isolated from bovine milk. Vet. Microbiol. 2009, 134, 73–81. [Google Scholar] [CrossRef] [PubMed]
- CLSI Document M100–S31. Performance Standards for Antimicrobial Susceptibility Testing; Thirty–First Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021; p. 350. [Google Scholar]
- The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters; Version 11.0; The European Committee on Antimicrobial Susceptibility Testing: Basel, Switzerland, 2021; Available online: http://www.eucast.org (accessed on 15 February 2021).
- Roychoudhury, S.; Twinem, T.L.; Makin, K.M.; Nienaber, M.A.; Li, C.; Morris, T.W.; Ledoussal, B.; Catrenich, C.E. Staphylococcus aureus mutants isolated via exposure to nonfluorinated quinolones: Detection of known and unique mutations Antimicrob. Agents Chemother. 2001, 45, 3422–3426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI Document M2–A13. Performance Standards for Antimicrobial Disk Susceptibility Tests; Thirteenth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; p. 92. [Google Scholar]
- CLSI Document M7–A11. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Eleventh Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; p. 91. [Google Scholar]
- Barry, A.L.; Jones, R.N. Reliability of high-content disks and modified broth dilution tests for detecting staphylococcal resistance to the penicilinase-resistant penicillins. J. Clin. Microbiol. 1987, 25, 1897–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauwelier, B.; Gordts, B.; Descheemaecker, P.; Van Landuyt, H. Evaluation of a disk diffusion method with cefoxitin (30 microg) for detection of methicillin-resistant Staphyloccocus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 389–392. [Google Scholar] [CrossRef]
- Boutiba-Ben Boubaker, I.; Ben Abbes, R.; Ben Abdallah, H.; Mamlouk, K.; Mahjoubi, F.; Kammoun, A.; Hammami, A.; Ben Redjeb, S. Evaluation of a cefoxitin disk diffusion test for the routine detection of methicillin-resistant Staphyloccocus aureus. Clin. Microbiol. Infect. 2004, 10, 762–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swenson, J.M.; Tenover, F.C. Cefoxitin disk study group. Results of disk diffusion testing with cefoxitin correlate with presence of mecA in Staphylococcus spp. J. Clin. Microbiol. 2005, 43, 3818–3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, S.; Sistla, S.; Manoharan, M.; Sugumar, M.; Nagasundaram, N.; Parija, S.C.; Pallab, R.; Bakthavatchalam, Y.D.; Veeraraghavan, B.; Kapil, A.; et al. Prevalence and genetic mechanisms of antimicrobial resistance in Staphylococcus species: A multicentre report of the indian council of medical research antimicrobial resistance surveillance network. Indian J Med. Microbiol. 2017, 35, 53–60. [Google Scholar] [CrossRef]
- Loncaric, I.; Kübber-Heiss, A.; Posautz, A.; Stalder, G.L.; Hoffmann, D.; Rosengarten, R.; Walzer, C. Characterization of methicillin-resistant Staphylococcus spp. carrying the mecC gene, isolated from wildlife. J. Antimicrob. Chemother. 2013, 68, 2222–2225. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-encoded transferable mecB-mediated methicillin resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sanz, E.; Schwendener, S.; Thomann, A.; Brawand, S.G.; Perreten, V. First staphylococcal cassette chromosome mec containing a mecB-carrying gene complex independent of transposon Tn6045 in a Macrococcus caseolyticus isolate from a canine infection. Antimicrob. Agents Chemother. 2015, 59, 4577–4583. [Google Scholar] [CrossRef] [Green Version]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC), Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents. Chemother. 2009, 53, 4961–4967. [CrossRef] [Green Version]
- Tsubakishita, S.; Kuwahara-Arai, K.; Sasaki, T.; Hiramatsu, K. Origin and molecular evolution of the determinant of methicillin resistance in staphylococci. Antimicrob. Agents. Chemother. 2010, 54, 4352–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Founou, L.L.; Founou, R.C.; Essack, S.Y.; Djoko, C.F. Mannitol-fermenting methicillin-resistant staphylococci (MRS) in pig abattoirs in Cameroon and South Africa: A serious food safety threat. Int. J. Food Microbiol. 2018, 285, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Saka, E.; Gulel, G.T. Detection of enterotoxin genes and methicillin-resistance in Staphylococcus aureus isolated from water buffalo milk and dairy products. J. Food Sci. 2018, 83, 1716–1722. [Google Scholar] [CrossRef] [PubMed]
- Adame-Gómez, R.; Toribio-Jimenez, J.; Vences-Velázquez, A.; Rodríguez-Bataz, E.; Dionisio, M.C.S.; Ramírez-Peralta, A. Methicillin-Resistant Staphylococcus aureus (MRSA) in Artisanal Cheeses in México. Int. J. Microbiol. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo, A.; Carriedo, J.A.; Beneitez, E.; Juarez, M.T.; De La Fuente, L.F.; San, F. Primitivo short communication: Bulk tank total bacterial count in dairy sheep: Factors of variation and relationship with somatic cell count. J. Dairy Sci. 2006, 89, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Van Hoek, A.H.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J. Acquired antibiotic resistance genes: An overview. Front. Microbiol. 2011, 2, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.C.; Sutcliffe, J.; Courvalin, P.; Jensen, L.B.; Rood, J.; Seppala, H. Nomenclature for macrolide and macrolide-lincosamide-streptogramin B resistance determinants. Antimicrob. Agents Chemother. 1999, 43, 2823–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, A.F.; Fitzgerald, J.R.; Penadés, J.R. Staphylococcus aureus in Animals. In Gram-Positive Pathogens, 3rd ed.; Fischetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A., Braunstein, M., Rood, J.I., Eds.; American Society for Microbiology: Washington, DC, USA, 2019; pp. 731–746. [Google Scholar] [CrossRef] [Green Version]
- Vautor, E.; Carsenti-Dellamonica, H.; Sabah, M.; Mancini, G.; Pépin, M.; Dellamonica, P. Characterization of Staphylococcus aureus isolates recovered from dairy sheep farms (agr group, adherence, slime, resistance to antibiotics). Small Rumin. Res. 2007, 72, 197–199. [Google Scholar] [CrossRef]
- Ergün, Y.; Öztürk, F.; Aslantaş, Ö.; Ceylan, A.; Kireççi, E.; Boyar, Y. Antimicrobial susceptibility, presence of resistance genes and biofilm formation in coagulase negative staphlococci ısolated from subclinical sheep mastitis. Kafkas Univer. Veter. Fakult. Derg. 2012, 18, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Ünal, N.; Askar, Ş.; Macun, H.C.; Sakarya, F.; Altun, B.; Yıldırım, M. Panton–Valentine leukocidin and some exotoxins of Staphylococcus aureus and antimicrobial susceptibility profiles of staphylococci isolated from milks of small ruminants. Trop. Anim. Health Prod. 2012, 44, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Vasileiou, N.G.; Sarrou, S.; Papagiannitsis, C.; Chatzopoulos, D.C.; Malli, E.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Antimicrobial agent susceptibility and typing of staphylococcal isolates from subclinical mastitis in ewes. Microb. Drug Resist. 2019, 25, 1099–1110. [Google Scholar] [CrossRef]
- Azara, E.; Longheu, C.; Sanna, G.; Tola, S. Biofilm formation and virulence factor analysis of Staphylococcus aureus isolates collected from ovine mastitis. J. Appl. Microbiol. 2017, 123, 372–379. [Google Scholar] [CrossRef] [PubMed]
- França, C.A.; Peixoto, R.M.; Cavalcante, M.B.; Melo, N.F.; Oliveira, C.J.B.; Veschi, J.L.A.; Mota, R.A.; Costa, M.M. Antimicrobial resistance of Staphylococcus spp. from small ruminant mastitis in Brazil. Pesqui. Vet. Bras. 2012, 32, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Lüthje, P.; Schwartz, S. Antimicrobial resistance of coagulase-negative staphylococci from bovine subclinical mastitis with particular reference to macrolide–lincosamide resistance phenotypes and genotypes. J. Antimicrob. Chemother. 2006, 57, 966–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.I.; Kim, S.D.; Park, J.H.; Yang, S.J. Species distribution, antimicrobial resistance, and enterotoxigenicity of Non-aureus staphylococci in retail chicken meat. Antibiotics 2020, 11, 809. [Google Scholar] [CrossRef]
- Devriese, L.A.; Baele, M.; Vaneechoutte, M.; Martel, A.; Haesebrouck, F. Identification and antimicrobial susceptibility of Staphylococcus chromogenes isolates from intramammary infections of dairy cows. Vet. Microbiol. 2002, 87, 175–182. [Google Scholar] [CrossRef]
- Catry, B.; Devriese, L.A.; Laevens, H.; De Vliegher, S.; Vaneechoutte, M.; Opsomer, G.; de Kruif, A. Antibiotic susceptibility and resistance of Staphylococcus chromogenes from bovine mastitis. Acta Vet. Scand. 2003, 44, 89. [Google Scholar] [CrossRef] [Green Version]
- Kenar, B.; Kuyucuoǧlu, Y.; Şeker, E. Antibiotic susceptibility of coagulase-negative staphylococci isolated from bovine subclinical mastitis in Turkey. Pak. Vet. J. 2012, 32, 390–393. [Google Scholar]
- Phophi, L.; Petzer, I.M.; Qekwana, D.N. Antimicrobial resistance patterns and biofilm formation of coagulase-negative Staphylococcus species isolated from subclinical mastitis cow milk samples submitted to the Onderstepoort Milk Laboratory. BMC Vet. Res. 2019, 15, 420. [Google Scholar] [CrossRef]
- Nunes, R.S.C.; Del Aguila, E.M.; Paschoalin, V.M.F. Safety evaluation of the coagulase-negative staphylococci microbiota of salami: Superantigenic Toxin Production and antimicrobial resistance. BioMed Res. Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; De Schaetzen, M.-A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [Green Version]
- Resch, M.; Nagel, V.; Hertel, C. Antibiotic resistance of coagulase-negative staphylococci associated with food and used in starter cultures. Int. J. Food Microbiol. 2008, 127, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Manuzon, M.; Lehman, M.; Wan, K.; Luo, H.; Wittum, T.E.; Yousef, A.; Bakaletz, L.O. Food commensal microbes as a potentially important avenue in transmitting antibiotic resistance genes. FEMS Microbiol. Lett. 2006, 254, 226–231. [Google Scholar] [CrossRef]
- Toomey, N.; Bolton, D.; Fanning, S. Characterisation and transferability of antibiotic resistance genes from lactic acid bacteria isolated from Irish pork and beef abattoirs. Res. Microbiol. 2010, 161, 127–135. [Google Scholar] [CrossRef]
- Potenski, C.J.; Gandhi, M.; Matthews, K.R. Exposure of Salmonella Enteritidis to chlorine or food preservatives decreases [corrected] susceptibility to antibiotics. FEMS Microbiol. Lett. 2003, 220, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Semjon, B.; Reitznerová, A.; Poláková, Z.; Výrostková, J.; Mal’ová, J.; Koréneková, B.; Dudriková, E.; Lovayová, V. The effect of traditional production methods on microbial, physico-chemical and sensory properties of ‘‘Slovenská bryndza’’ Protected Geographical Indication cheese. Int. J. Dairy Technol. 2018, 71, 709–716. [Google Scholar] [CrossRef]
- ISO Standard 6887–5. Microbiology of Food and Animal Feeding Stuffs. Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination. Part 5: Specific Rules for the Preparation of Milk and Milk Products (ISO 6887–5:2010); Slovak Standards Institute: Bratislava, Slovakia, 2010. [Google Scholar]
- ISO Standard 6888–1. Microbiology of Food and Animal Feeding Stuffs. Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species). Part 1: Technique Using Baird-Parker Agar Medium (ISO 6888–1:1999); Slovak Standards Institute: Bratislava, Slovakia, 1999. [Google Scholar]
- Hein, I.; Jorgensen, H.J.; Loncarevic, S.; Wagner, M. Quantification of Staphylococcus aureus in unpasteurised bovine and caprine milk by real-time PCR. Res. Microbiol. 2005, 156, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruker Daltonics. MALDI Biotyper 2.0. Software for Microorganism Identification and Classification User Manual; Bruker Scientific LLC: Billerica, MA, USA, 2008. [Google Scholar]
- Poulsen, A.B.; Skov, R.; Pallesen, L.V. Detection of methicillin resistance in coagulase-negative staphylococci and in staphylococci directly from simulated blood cultures using the EVIGENE MRSA Detection Kit. J. Antimicrob. Chemother. 2003, 51, 419–421. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Strains | MIC (mg/L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ATB | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | OR | p Value | |
| Staphylococcus chromogenes (n = 37) | PEN | 2 | 19 | 16 | - | - | - | - | - | - | 1.3854 | 0.4853 |
| OX | 2 | 5 | 25 a | 5 b | - | - | - | - | - | 13.3333 | <0.0001 | |
| TEC | - | - | - | - | - | 2 | 3 | 32 | - | - | - | |
| GN | - | - | - | - | 11 | 11 | 9 | 5 | 1 | 5.6250 | 0.1237 | |
| E | - | 1 | 3 | 1 | 1 | 2 | 18 | 11 | - | 2.2392 | 0.0981 | |
| TE | - | - | - | - | 1 | 3 | 1 | 15 | 17 | 0.8021 | 0.6390 | |
| OFX | - | - | 7 | 13 | 5 | 10 a | 2 b | - | - | 6.4815 | 0.0220 | |
| S. chromogenes | Antibiotics | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| PEN | OX | TEC | GN | E | TE | OFX | |||
| sample of sheep’s milk | (n = 23) | S | 2 | 5 | 4 | 15 | 3 | 3 | 15 |
| I | 0 | 0 | 18 | 4 | 3 | 0 | 2 | ||
| R | 21 | 18 | 1 | 4 | 17 | 20 | 6 | ||
| sample of sheep’s cheese | (n = 14) | S | 0 | 2 | 1 | 7 | 1 | 1 | 5 |
| I | 0 | 0 | 13 | 5 | 1 | 1 | 3 | ||
| R | 14 | 12 | 0 | 2 | 12 | 12 | 6 | ||
| Resistance Phenotype | Sheep’s Milk | Sheep’s Cheese |
|---|---|---|
| PEN-E | 1 | 0 |
| PEN-TE | 2 | 1 |
| PEN-OX | 3 * | 2 * |
| PEN-OX-E | 0 | 2 |
| PEN-OX-TE | 3 * | 0 |
| PEN-OX-TE-E | 2 | 3 |
| PEN-OX-TE-E-GN | 0 | 1 * |
| PEN-OX-TE-E-OFX | 1 | 0 |
| PEN-OX-TE-E-GN-OFX | 1 * | 1 * |
| ∑ | 13 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regecová, I.; Výrostková, J.; Zigo, F.; Gregová, G.; Kováčová, M. Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese. Antibiotics 2021, 10, 570. https://doi.org/10.3390/antibiotics10050570
Regecová I, Výrostková J, Zigo F, Gregová G, Kováčová M. Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese. Antibiotics. 2021; 10(5):570. https://doi.org/10.3390/antibiotics10050570
Chicago/Turabian StyleRegecová, Ivana, Jana Výrostková, František Zigo, Gabriela Gregová, and Mariana Kováčová. 2021. "Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese" Antibiotics 10, no. 5: 570. https://doi.org/10.3390/antibiotics10050570
APA StyleRegecová, I., Výrostková, J., Zigo, F., Gregová, G., & Kováčová, M. (2021). Detection of Antimicrobial Resistance of Bacteria Staphylococcus chromogenes Isolated from Sheep’s Milk and Cheese. Antibiotics, 10(5), 570. https://doi.org/10.3390/antibiotics10050570