
Novel Bifunctional Acylase from Actinoplanes utahensis: A Versatile Enzyme to Synthesize Antimicrobial Compounds and Use in Quorum Quenching Processes

 , , and
, , and
Abstract
:1. Introduction
2. Results
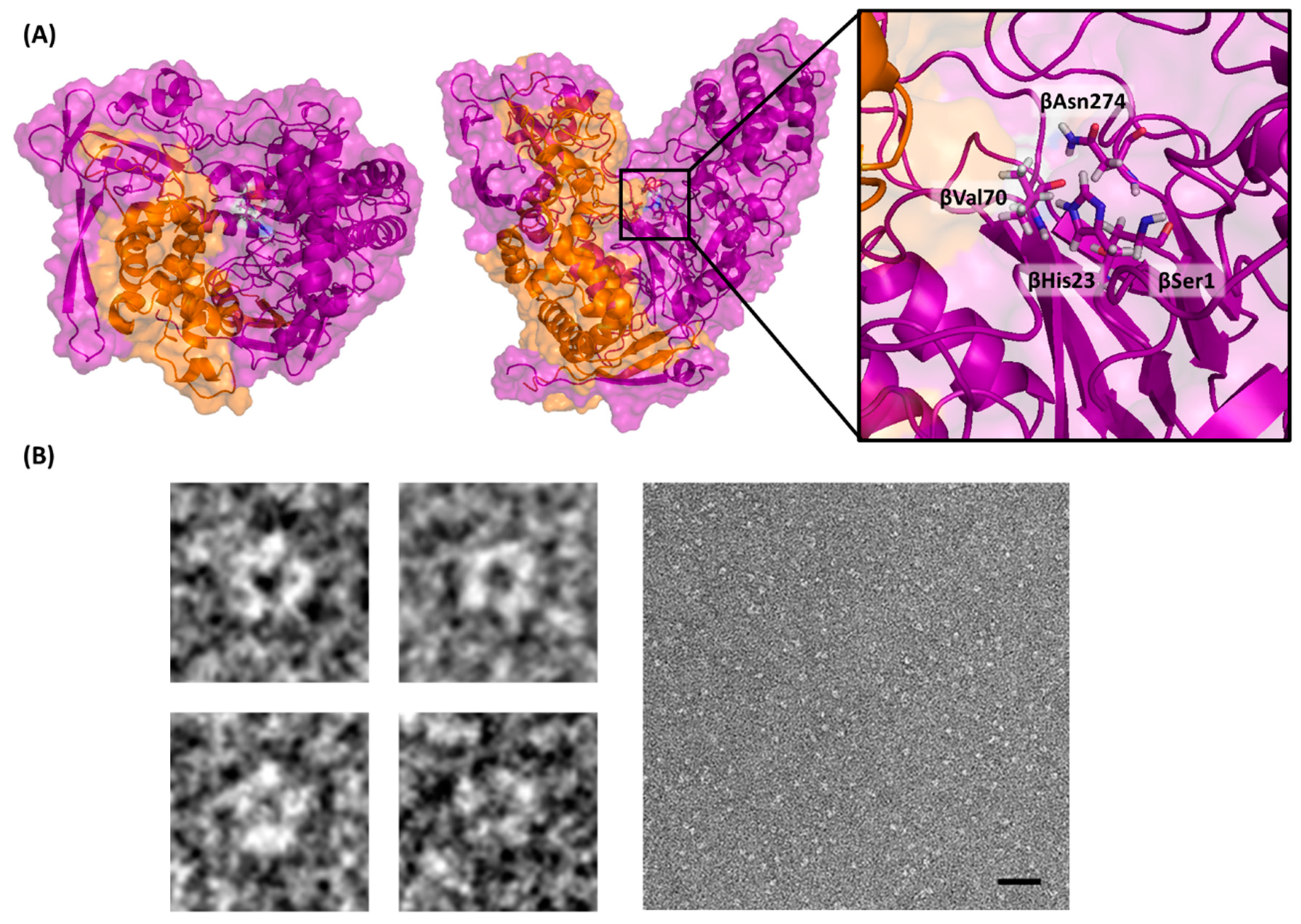
2.1. Identification of the Putative ahla Gene in the Genome of Actinoplanes utahensis NRRL 12052
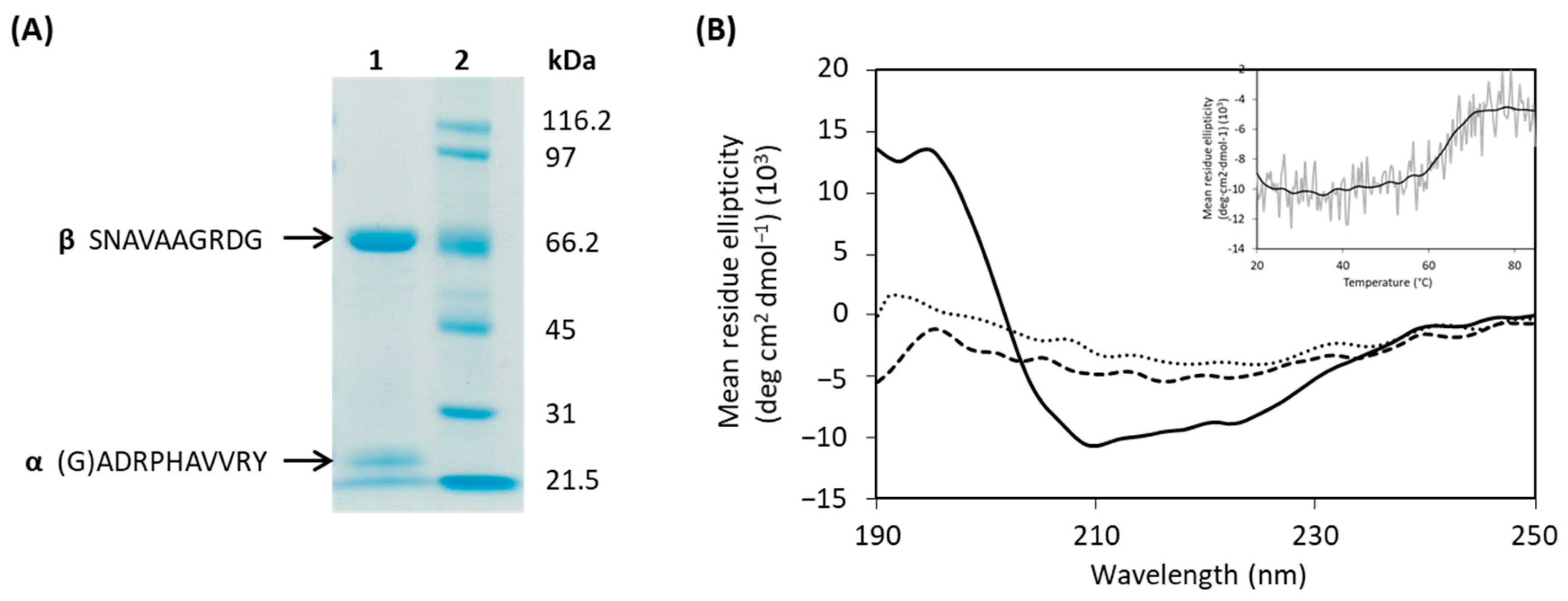
2.2. Heterologous Expression and Purification of AuAHLA
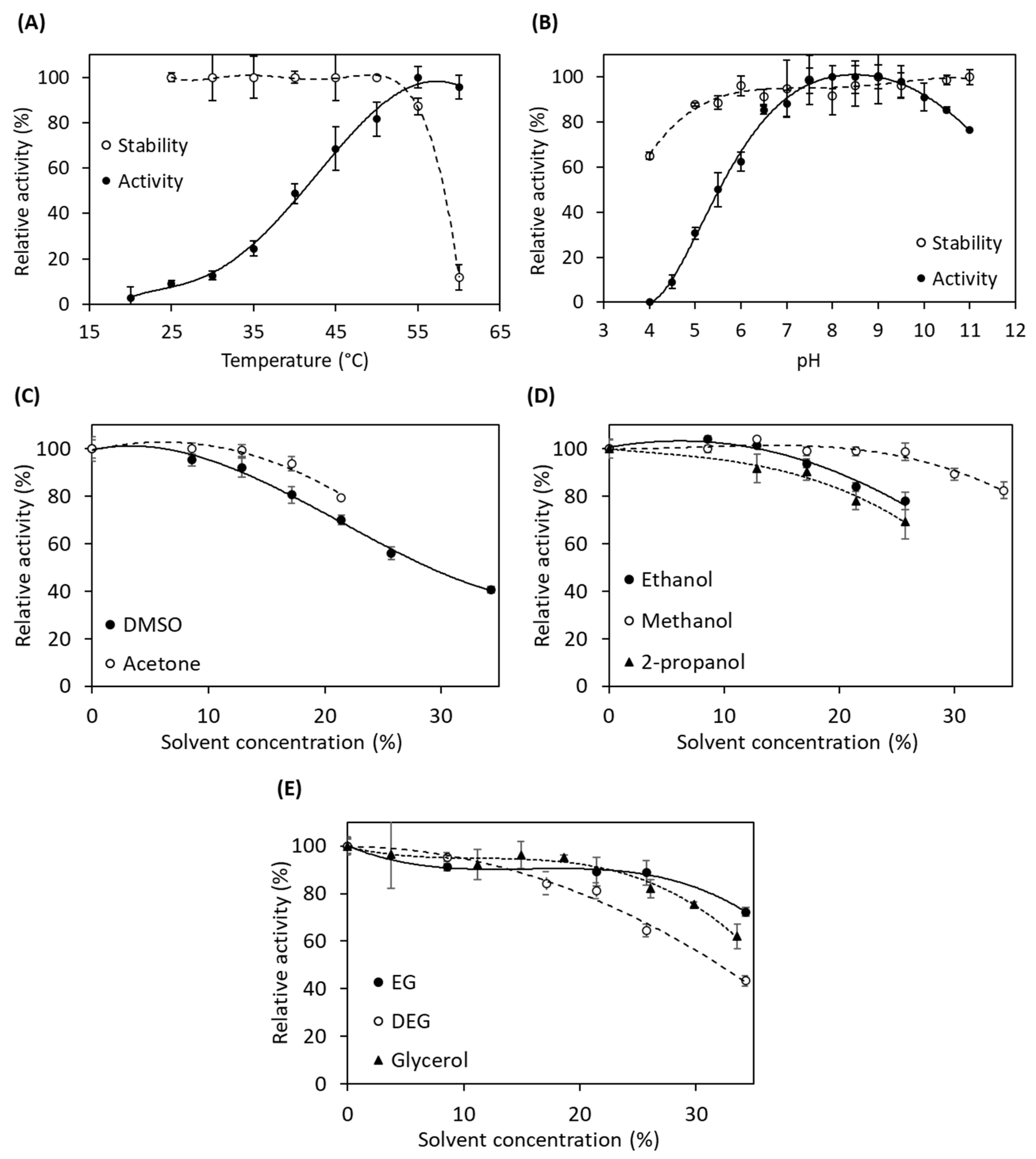
2.3. Characterization of Recombinant AuAHLA
2.4. Kinetic Parameters of AuAHLA for the Hydrolysis of Different Substrates
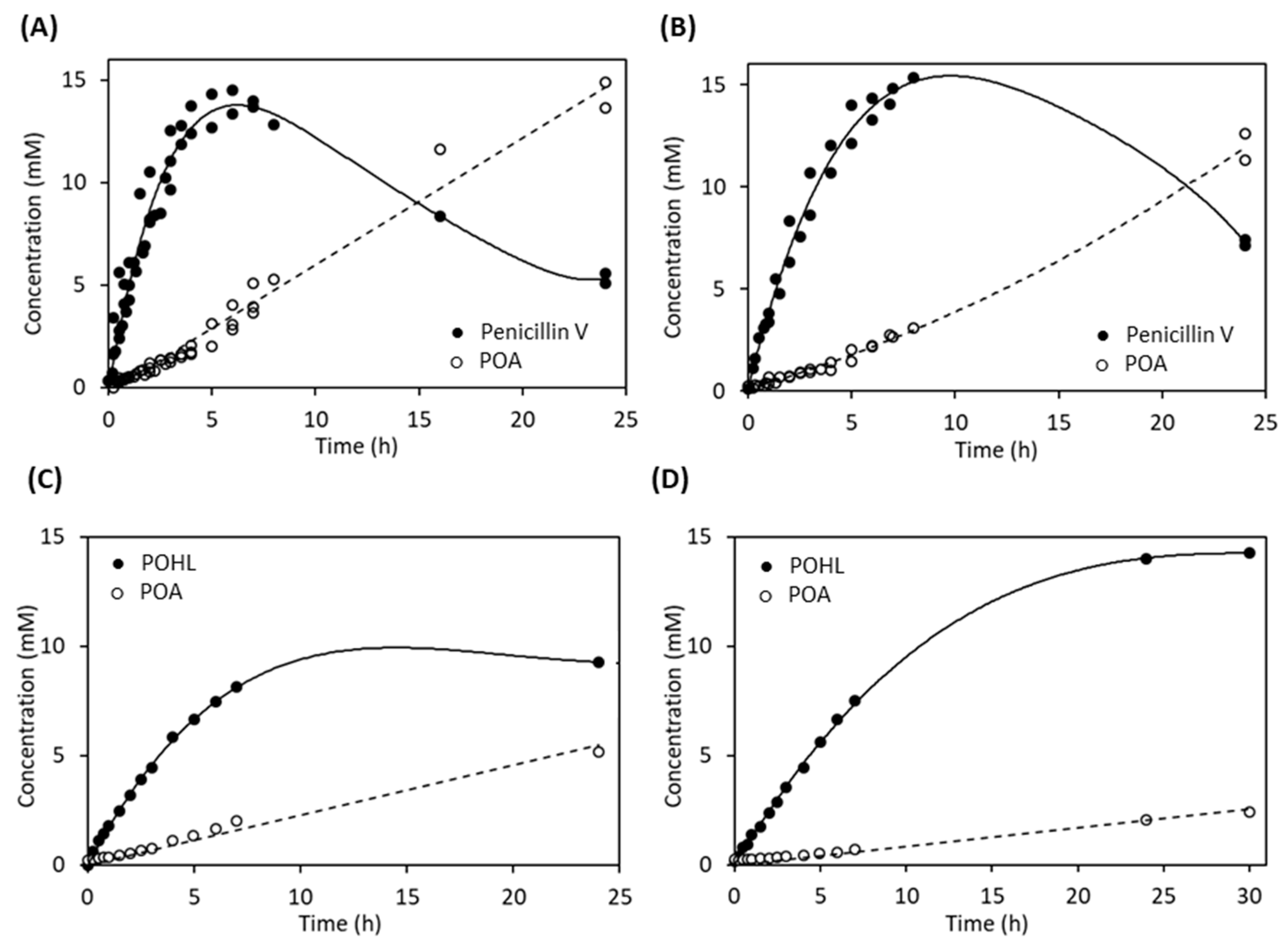
2.5. Synthetic Activity of AuAHLA
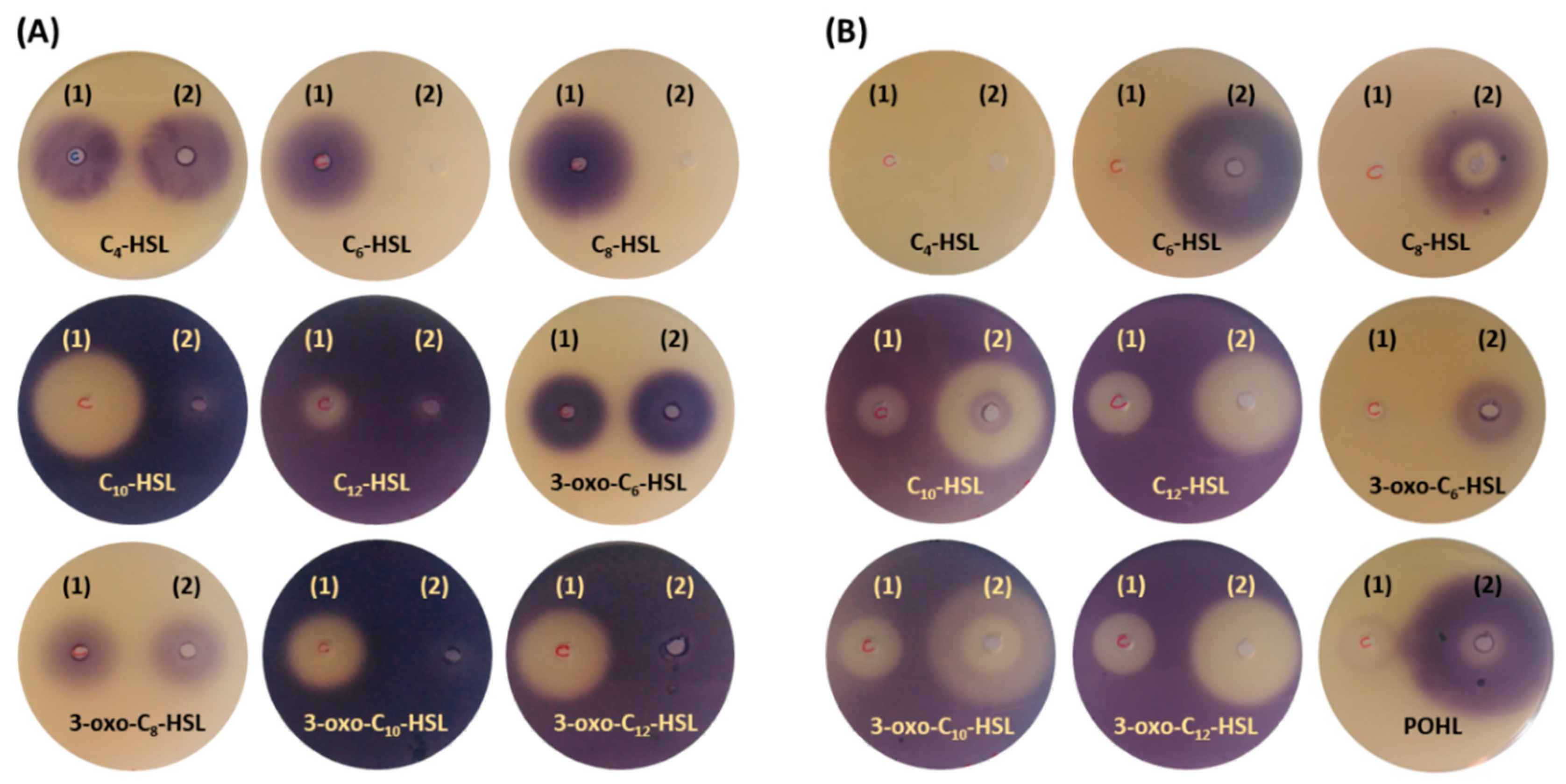
2.6. Bioassay Activity of Hydrolyzed and Synthetized AHLs by AuAHLA
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents
4.2. Microorganisms, Culture Conditions, Plasmids, and DNA Manipulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain, Plasmid, or Primer | Relevant Description or Sequence | Reference |
|---|---|---|
| Actinoplanes utahensis NRRL 12052 | Native N-acyl-homoserine lactone acylase producer | [23,24,71] |
| Escherichia coli DH5α | Host for recombinant plasmid F− λ- φ80dlacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17(rK− mK+) supE44 thi-1 gyrA96 relA1 | [64] |
| Rhodococcus sp. T104 KACC 21099 | Host for gene expression (Kns) | [38] |
| Rhodococcus pENV19ahla | Recombinant strain harboring the plasmid pENV19ahla | This study |
| Chromobacterium violaceum CV026 CECT 5999 | cvil::mini-Tn5-mutant of Chromobacterium violaceum ATCC 31532 (KnR) | [67] |
| pENV19 | Shuttle vector for E. coli-Rhodococcus with the constitutive permE* promoter (5.1 Kb, KnR pAL5000ori permE * ColE1ori) | [29,38,39] |
| pENV19ahla | pENV19-containing ahla gene from A. utahensis NRRL 12052 | This study |
| AHLA1 | 5’-GCTCTAGAGGAGGTGCCGCCGTGGCCCGTCCGTTCA-3’ | This study |
| AHLA2 | 5’-CGGAATTCCTCACCGCGGCGCTCGCTCGGTCAGTCTGAT-3’ | This study |
4.3. Overexpression of ahla Gene and Purification of Recombinant AuAHLA from Rhodococcus sp. T104
4.4. Protein Sequence and Structure Analysis
4.5. Acylase Activity Assay
4.6. Biochemical Characterization and Kinetic Parameters
4.7. Enzymatic Synthesis of Penicillin V and AHLs
4.8. Bioassay Activity of Hydrolyzed and Synthetized AHLs Using Chromobacterium violaceum CV026
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Dong, Y.H.; Zhang, L.H. Quorum sensing and quorum-quenching enzymes. J. Microbiol. 2005, 43, 101–109. [Google Scholar] [PubMed]
- LaSarre, B.; Federle, M.J. Exploiting Quorum Sensing to Confuse Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: The yin and yang of bacterial communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef]
- Uroz, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Chiou, J.; Chatterjee, P.; Otto, M. Alternative approaches to treat bacterial infections: Targeting quorum-sensing. Expert Rev. Anti Infect. Ther. 2020, 18, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Bzdrenga, J.; Daude, D.; Remy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabriere, E. Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef]
- Soler, A.; Arregui, L.; Arroyo, M.; Mendoza, J.A.; Muras, A.; Alvarez, C.; Garcia-Vera, C.; Marquina, D.; Santos, A.; Serrano, S. Quorum Sensing versus Quenching Bacterial Isolates Obtained from MBR Plants Treating Leachates from Municipal Solid Waste. Int. J. Environ. Res. Public Health 2018, 15, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köse-Mutlu, B.; Ergön-Can, T.; Koyuncu, I.; Lee, C.-H. Quorum quenching for effective control of biofouling in membrane bioreactor: A comprehensive review of approaches, applications, and challenges. Environ. Eng. Res. 2019, 24, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Geske, G.D.; O’Neill, J.C.; Blackwell, H.E. Expanding dialogues: From natural autoinducers to non-natural analogues that modulate quorum sensing in Gram-negative bacteria. Chem. Soc. Rev. 2008, 37, 1432–1447. [Google Scholar] [CrossRef] [PubMed]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Duggleby, H.J.; Tolley, S.P.; Hill, C.P.; Dodson, E.J.; Dodson, G.; Moody, P.C. Penicillin acylase has a single-amino-acid catalytic centre. Nature 1995, 373, 264–268. [Google Scholar] [CrossRef]
- Oinonen, C.; Rouvinen, J. Structural comparison of Ntn-hydrolases. Protein Sci. 2000, 9, 2329–2337. [Google Scholar] [CrossRef] [Green Version]
- Sudhakaran, V.K.; Deshpande, B.S.; Ambedkar, S.S.; Shewale, J.G. Molecular aspects of penicillin and cephalosporin acylases. Process. Biochem. 1992, 27, 131–143. [Google Scholar] [CrossRef]
- Sunder, A.V.; Utari, P.D.; Ramasamy, S.; van Merkerk, R.; Quax, W.; Pundle, A. Penicillin V acylases from gram-negative bacteria degrade N-acylhomoserine lactones and attenuate virulence in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 2383–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utari, P.D.; Vogel, J.; Quax, W.J. Deciphering Physiological Functions of AHL Quorum Quenching Acylases. Front. Microbiol. 2017, 8, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruggink, A.; Roos, E.C.; de Vroom, E. Penicillin acylase in the industrial production of beta-lactam antibiotics. Org. Process. Res. Dev. 1998, 2, 128–133. [Google Scholar] [CrossRef]
- Arroyo, M.; de la Mata, I.; Acebal, C.; Castillón, M.P. Biotechnological applications of penicillin acylases: State-of-the-art. Appl. Microbiol. Biotechnol. 2003, 60, 507–514. [Google Scholar] [CrossRef]
- Velasco-Bucheli, R.; del Cerro, C.; Hormigo, D.; Acebal, C.; Arroyo, M.; García, J.L.; de la Mata, I. Draft genome sequence of Actinoplanes utahensis NRRL 12052, a microorganism involved in industrial production of pharmaceutical intermediates. Genome Announc. 2015, 3, e01411–e01414. [Google Scholar] [CrossRef] [Green Version]
- Inokoshi, J.; Takeshima, H.; Ikeda, H.; Omura, S. Cloning and sequencing of the aculeacin A acylase-encoding gene from Actinoplanes utahensis and expression in Streptomyces lividans. Gene 1992, 119, 29–35. [Google Scholar] [CrossRef]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Torres-Bacete, J.; Hormigo, D.; Stuart, M.; Arroyo, M.; Torres, P.; Castillón, M.P.; Acebal, C.; García, J.L.; de la Mata, I. Newly discovered penicillin acylase activity of aculeacin A acylase from Actinoplanes utahensis. Appl. Environ. Microbiol. 2007, 73, 5378–5381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Bacete, J.; Hormigo, D.; Torres-Gúzman, R.; Arroyo, M.; Castillón, M.P.; García, J.L.; Acebal, C.; de la Mata, I. Overexpression of penicillin V acylase from Streptomyces lavendulae and elucidation of its catalytic residues. Appl. Environ. Microbiol. 2015, 81, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Koreishi, M.; Imanaka, H.; Imamura, K.; Nakanishi, K. Cloning and characterization of penicillin V acylase from Streptomyces mobaraensis. J. Biotechnol. 2007, 128, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, S.; Shibata, T.; Ito, K.; Oohata, N.; Yamashita, M.; Hino, M.; Yamada, M.; Isogai, Y.; Hashimoto, S. Cloning and expression of the FR901379 acylase gene from Streptomyces sp. no. 6907. J. Antibiot. 2011, 64, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelhoch, H. Spectroscopic determination of tryptophan and tyrosine in proteins. Biochemistry 1967, 6, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- Shannon, G.; Marples, C.R.; Toofanny, R.D.; Williams, P.M. Evolutionary drivers of protein shape. Sci. Rep. 2019, 9, 11873. [Google Scholar] [CrossRef] [PubMed]
- Hormigo, D.; Lopez-Conejo, M.T.; Serrano-Aguirre, L.; García-Martín, A.; Saborido, A.; de la Mata, I.; Arroyo, M. Kinetically controlled acylation of 6-APA catalyzed by penicillin acylase from Streptomyces lavendulae: Effect of reaction conditions in the enzymatic synthesis of penicillin V. Biocatal. Biotransform. 2020, 38, 253–262. [Google Scholar] [CrossRef]
- Geske, G.D.; Mattmann, M.E.; Blackwell, H.E. Evaluation of a focused library of N-aryl L-homoserine lactones reveals a new set of potent quorum sensing modulators. Bioorg. Med. Chem. Lett. 2008, 18, 5978–5981. [Google Scholar] [CrossRef] [Green Version]
- García-Hidalgo, J.; Hormigo, D.; Prieto, M.; Arroyo, M.; de la Mata, I. Extracellular production of Streptomyces exfoliatus poly(3-hydroxybutyrate) depolymerase in Rhodococcus sp. T104: Determination of optimal biocatalyst conditions. Appl. Microbiol. Biotechnol. 2012, 93, 1975–1988. [Google Scholar] [CrossRef]
- García-Hidalgo, J.; Hormigo, D.; Arroyo, M.; de la Mata, I. Novel extracellular PHB depolymerase from Streptomyces ascomycinicus: PHB copolymers degradation in acidic conditions. PLoS ONE 2013, 8, e71699. [Google Scholar] [CrossRef] [Green Version]
- Torres, R.; de la Mata, I.; Castillón, M.P.; Arroyo, M.; Torres, J.; Acebal, C. Purification and characterization of penicillin V acylase from Streptomyces lavendulae. In Progress in Biotechnology; Ballesteros, A., Plou, F.J., Iborra, J.L., Halling, P.J., Eds.; Elsevier: Cordoba, Spain, 1998; pp. 719–724. [Google Scholar]
- Koch, G.; Nadal-Jimenez, P.; Cool, R.H.; Quax, W.J. Deinococcus radiodurans can interfere with quorum sensing by producing an AHL-acylase and an AHL-lactonase. FEMS Microbiol. Lett. 2014, 356, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-N.; Chen, C.-J.; Liao, C.-T.; Lee, C.-Y. A probable aculeacin A acylase from the Ralstonia solanacearum GMI1000 is N-acyl-homoserine lactone acylase with quorum-quenching activity. BMC Microbiol. 2009, 9, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-H.; Xu, J.-L.; Hu, J.; Wang, L.-H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.-H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol. 2003, 47, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.C.; Romero, M.; Salto, R.; Camara, M.; Llamas, I. AhaP, A Quorum Quenching Acylase from Psychrobacter sp. M9-54-1 That Attenuates Pseudomonas aeruginosa and Vibrio coralliilyticus Virulence. Mar. Drugs 2021, 19, 16. [Google Scholar] [CrossRef]
- Kusada, H.; Tamaki, H.; Kamagata, Y.; Hanada, S.; Kimura, N. A Novel Quorum-Quenching N-Acylhomoserine Lactone Acylase from Acidovorax sp Strain MR-S7 Mediates Antibiotic Resistance. Appl. Environ. Microbiol. 2017, 83, e00080-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, R.W.; Lindow, S.E. Two dissimilar N-acyl-homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm morphology. Appl. Environ. Microbiol. 2009, 75, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Bokhove, M.; Jimenez, P.N.; Quax, W.J.; Dijkstra, B.W. The quorum-quenching N-acyl homoserine lactone acylase PvdQ is an Ntn-hydrolase with an unusual substrate-binding pocket. Proc. Natl. Acad. Sci. USA 2010, 107, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morohoshi, T.; Nakazawa, S.; Ebata, A.; Kato, N.; Ikeda, T. Identification and characterization of N-acylhomoserine lactone-acylase from the fish intestinal Shewanella sp. strain MIB015. Biosci. Biotechnol. Biochem. 2008, 72, 1887–1893. [Google Scholar] [CrossRef] [Green Version]
- Avinash, V.S.; Pundle, A.V.; Ramasamy, S.; Suresh, C.G. Penicillin acylases revisited: Importance beyond their industrial utility. Crit. Rev. Biotechnol. 2016, 36, 303–316. [Google Scholar] [CrossRef]
- Philem, P.D.; Yadav, Y.; Vellore Sunder, A.; Ghosh, D.; Prabhune, A.; Ramasamy, S. Structural and enzymatic analysis of a dimeric cholylglycine hydrolase like acylase active on N-acyl homoserine lactones. Biochimie 2020, 177, 108–116. [Google Scholar] [CrossRef]
- Liu, N.; Yu, M.; Zhao, Y.; Cheng, J.; An, K.; Zhang, X.H. PfmA, a novel quorum-quenching N-acylhomoserine lactone acylase from Pseudoalteromonas flavipulchra. Microbiology 2017, 163, 1389–1398. [Google Scholar] [CrossRef]
- Mukherji, R.; Varshney, N.K.; Panigrahi, P.; Suresh, C.G.; Prabhune, A. A new role for penicillin acylases: Degradation of acyl homoserine lactone quorum sensing signals by Kluyvera citrophila penicillin G acylase. Enzym. Microb. Technol. 2014, 56, 1–7. [Google Scholar] [CrossRef]
- Velasco-Bucheli, R.; Hormigo, D.; Fernández-Lucas, J.; Torres-Ayuso, P.; Alfaro-Ureña, Y.; Saborido, A.I.; Serrano-Aguirre, L.; García, J.L.; Ramón, F.; Acebal, C.; et al. Penicillin Acylase from Streptomyces lavendulae and Aculeacin A Acylase from Actinoplanes utahensis: Two Versatile Enzymes as Useful Tools for Quorum Quenching Processes. Catalysts 2020, 10, 730. [Google Scholar] [CrossRef]
- Torres-Guzmán, R.; de la Mata, I.; Torres-Bacete, J.; Arroyo, M.; Castillón, M.P.; Acebal, C. Substrate specificity of penicillin acylase from Streptomyces lavendulae. Biochem. Biophys. Res. Commun. 2002, 291, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Wahjudi, M.; Papaioannou, E.; Hendrawati, O.; van Assen, A.H.G.; van Merkerk, R.; Cool, R.H.; Poelarends, G.J.; Quax, W.J. PA0305 of Pseudomonas aeruginosa is a quorum quenching acylhomoserine lactone acylase belonging to the Ntn hydrolase superfamily. Microbiology 2011, 157, 2042–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, G.; Nadal-Jimenez, P.; Reis, C.R.; Muntendam, R.; Bokhove, M.; Melillo, E.; Dijkstra, B.W.; Cool, R.H.; Quax, W.J. Reducing virulence of the human pathogen Burkholderia by altering the substrate specificity of the quorum-quenching acylase PvdQ. Proc. Natl. Acad. Sci. USA 2014, 111, 1568–1573. [Google Scholar] [CrossRef] [Green Version]
- Koreishi, M.; Tani, K.; Ise, Y.; Imanaka, H.; Imamura, K.; Nakanishi, K. Enzymatic synthesis of b-lactam antibiotics and N-fatty-acylated amino compounds by the acyl-transfer reaction catalyzed by penicillin V acylase from Streptomyces mobaraensis. Biosci. Biotechnol. Biochem. 2007, 71, 1582–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Càmara, M.; Daykin, M.; Chhabra, S.R. 6.12 Detection, purification, and synthesis of N-acylhomoserine lactone quorum sensing signal molecules. Methods Microbiol. 1998, 27, 319–330. [Google Scholar] [CrossRef]
- McInnis, C.E.; Blackwell, H.E. Non-native N-aroyl L-homoserine lactones are potent modulators of the quorum sensing receptor RpaR in Rhodopseudomonas palustris. Chembiochem. Eur. J. Chem. Biol. 2014, 15, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Syrpas, M.; Ruysbergh, E.; Stevens, C.V.; De Kimpe, N.; Mangelinckx, S. Synthesis and biological evaluation of novel N-α-haloacylated homoserine lactones as quorum sensing modulators. Beilstein J. Org. Chem. 2014, 10, 2539–2549. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Martinez, J.; Nieto-Alvarez, E.; Ramirez-Chavez, E.; Molina-Torres, J. Enzymatic Method for N-Acyl Homoserine Lactones Synthesis Using Immobilized Candida antarctica Lipase. Catal. Lett. 2018, 148, 62–67. [Google Scholar] [CrossRef]
- Burk, M.J.; Van Dien, S. Biotechnology for Chemical Production: Challenges and Opportunities. Trends Biotechnol. 2016, 34, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, A.R. Biocatalysis and Pharmaceuticals: A Smart Tool for Sustainable Development. Catalysts 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmid. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor: New York, NY, USA, 2001. [Google Scholar]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [Green Version]
- Quirós, L.M.; Aguirrezabalaga, I.; Olano, C.; Méndez, C.; Salas, J.A. Two glycosyltransferases and a glycosidase are involved in oleandomycin modification during its biosynthesis by Streptomyces antibioticus. Mol. Microbiol. 1998, 28, 1177–1185. [Google Scholar] [CrossRef]
- Chiba, K.; Hoshino, Y.; Ishino, K.; Kogure, T.; Mikami, Y.; Uehara, Y.; Ishikawa, J. Construction of a pair of practical Nocardia-Escherichia coli shuttle vectors. Jpn. J. Infect. Dis. 2007, 60, 45–47. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, H.; Inokoshi, J.; Takada, Y.; Tanaka, H.; Omura, S. A deacylation enzyme for aculeacin A, a neutral lipopeptide antibiotic, from Actinoplanes utahensis: Purification and characterization. J. Biochem. 1989, 105, 606–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–252. [Google Scholar] [CrossRef]
- Speicher, K.D.; Gorman, N.; Speicher, D.W. N-terminal sequence analysis of proteins and peptides. Curr. Protoc. Protein Sci. 2009, 57, 11.10.11–11.10.31. [Google Scholar] [CrossRef]
- Böhm, G.; Muhr, R.; Jaenicke, R. Quantitative analysis of protein far-UV circular dicroism spectra by neural networks. Protein Eng. 1992, 5, 191–195. [Google Scholar] [CrossRef]
- Ludtke, S.J.; Baldwin, P.R.; Chiu, W. EMAN: Semiautomated Software for High-Resolution Single-Particle Reconstructions. J. Struct. Biol. 1999, 128, 82–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasingham, K.; Warburton, D.; Dunnill, P.; Lilly, M.D. The isolation and kinetics of penicillin amidase from Escherichia coli. Biochim. Biophys. Acta 1972, 276, 250–256. [Google Scholar] [CrossRef]






| Substrate | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) |
|---|---|---|---|
| Penicillin V | 0.472 ± 0.05 | 106.34 ± 2.84 | 239.3 |
| Penicillin dihidroF | 0.199 ± 0.02 | 9.15 ± 0.28 | 48.8 |
| Penicillin K | 0.675 ± 0.19 | 2.33 ± 0.23 | 3.66 |
| Penicillin G | 1.21 ± 0.30 | 2.71 ± 0.18 | 2.39 |
| C4-HSL | ND | ND | ND |
| C6-HSL | 2.25 ± 0.50 | 6.07 ± 0.69 | 2.87 |
| C8-HSL | 0.860 ± 0.15 | 3.52 ± 0.29 | 4.35 |
| C10-HSL | 0.084 ± 0.021 | 2.59 ± 0.23 | 32.7 |
| C12-HSL | 0.008 ± 0.002 | 0.75 ± 0.045 | 99.5 |
| 3-oxo-C6-HSL | ND | ND | ND |
| 3-oxo-C8-HSL | 1.04 ± 0.42 | 0.19 ± 0.028 | 0.20 |
| 3-oxo-C10-HSL | 0.238 ± 0.086 | 0.50 ± 0.097 | 2.22 |
| 3-oxo-C12-HSL | 0.092 ± 0.021 | 0.53 ± 0.039 | 6.11 |
| Product | AuAHLA (IU/mL) | DMSO (%) | Vs (mM/h) | Vh (mM/h) | S/H | Ymax (%) | Time for Ymax (h) |
|---|---|---|---|---|---|---|---|
| Penicillin V | 0.125 | 3 | 0.85 | 0.40 | 2.13 | 30.8 | 24 |
| 0.25 | 3 | 1.57 | 0.53 | 2.99 | 50.5 | 24 | |
| 1 | 3 | 3.81 | 0.55 | 6.94 | 72.6 | 6 | |
| 1 | 32 | 3.70 | 0.37 | 10.1 | 76.8 | 8 | |
| POHL | 1 | 23 | 1.35 | 0.23 | 5.87 | 46.4 | 24 |
| 1 | 50 | 1.11 | 0.08 | 13.1 | 69.9 | 24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano-Aguirre, L.; Velasco-Bucheli, R.; García-Álvarez, B.; Saborido, A.; Arroyo, M.; de la Mata, I. Novel Bifunctional Acylase from Actinoplanes utahensis: A Versatile Enzyme to Synthesize Antimicrobial Compounds and Use in Quorum Quenching Processes. Antibiotics 2021, 10, 922. https://doi.org/10.3390/antibiotics10080922
Serrano-Aguirre L, Velasco-Bucheli R, García-Álvarez B, Saborido A, Arroyo M, de la Mata I. Novel Bifunctional Acylase from Actinoplanes utahensis: A Versatile Enzyme to Synthesize Antimicrobial Compounds and Use in Quorum Quenching Processes. Antibiotics. 2021; 10(8):922. https://doi.org/10.3390/antibiotics10080922
Chicago/Turabian StyleSerrano-Aguirre, Lara, Rodrigo Velasco-Bucheli, Begoña García-Álvarez, Ana Saborido, Miguel Arroyo, and Isabel de la Mata. 2021. "Novel Bifunctional Acylase from Actinoplanes utahensis: A Versatile Enzyme to Synthesize Antimicrobial Compounds and Use in Quorum Quenching Processes" Antibiotics 10, no. 8: 922. https://doi.org/10.3390/antibiotics10080922
APA StyleSerrano-Aguirre, L., Velasco-Bucheli, R., García-Álvarez, B., Saborido, A., Arroyo, M., & de la Mata, I. (2021). Novel Bifunctional Acylase from Actinoplanes utahensis: A Versatile Enzyme to Synthesize Antimicrobial Compounds and Use in Quorum Quenching Processes. Antibiotics, 10(8), 922. https://doi.org/10.3390/antibiotics10080922