



Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds

Abstract
:1. Introduction
2. Results
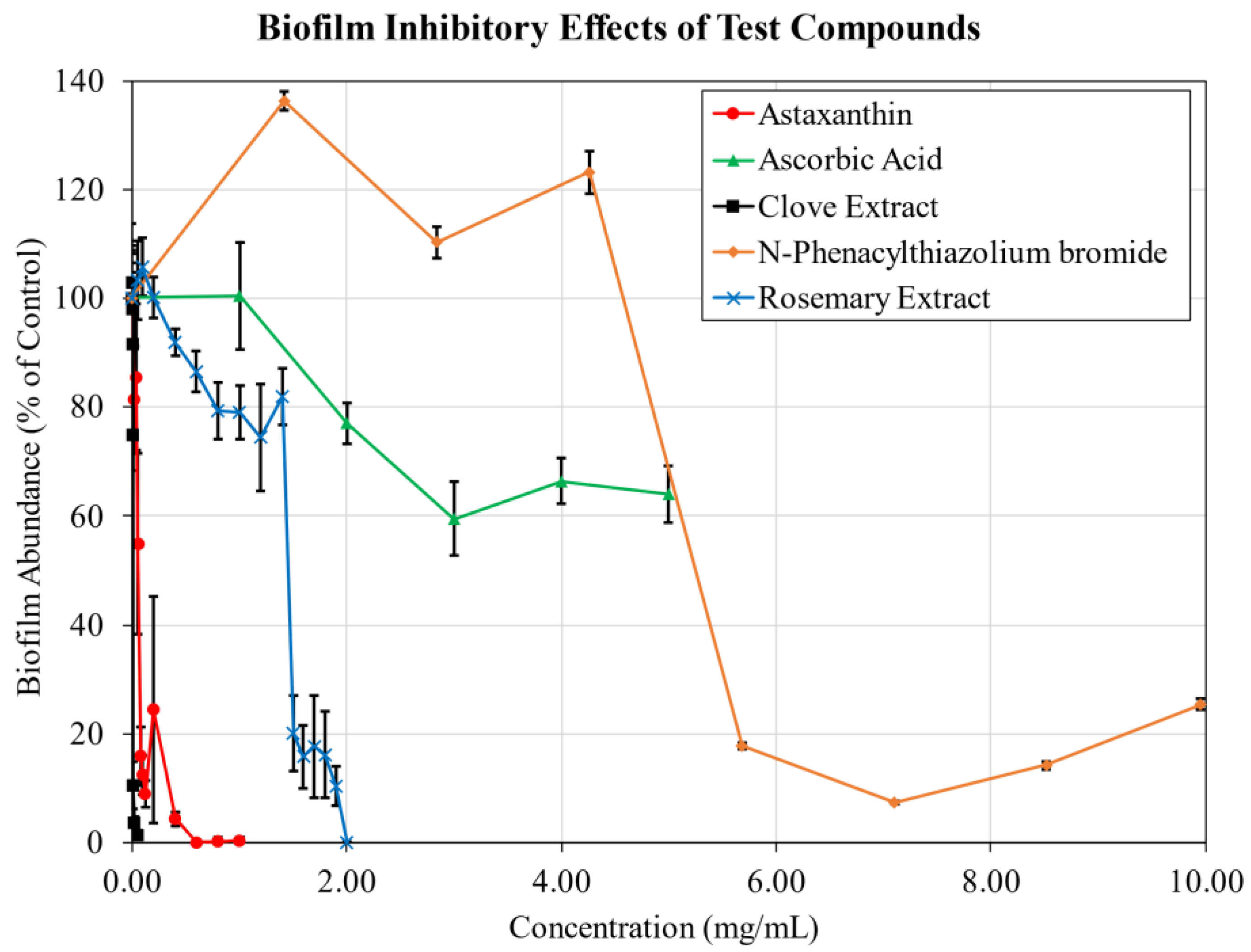
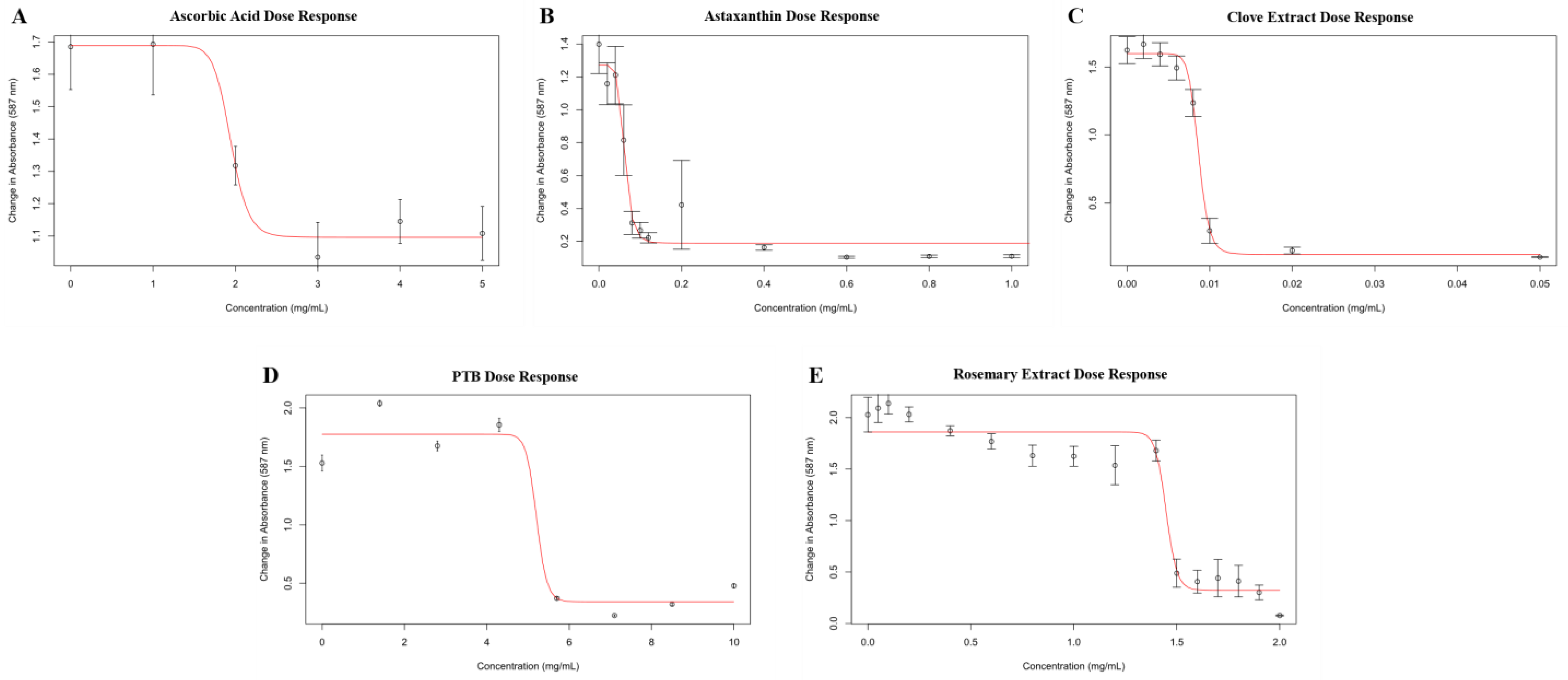
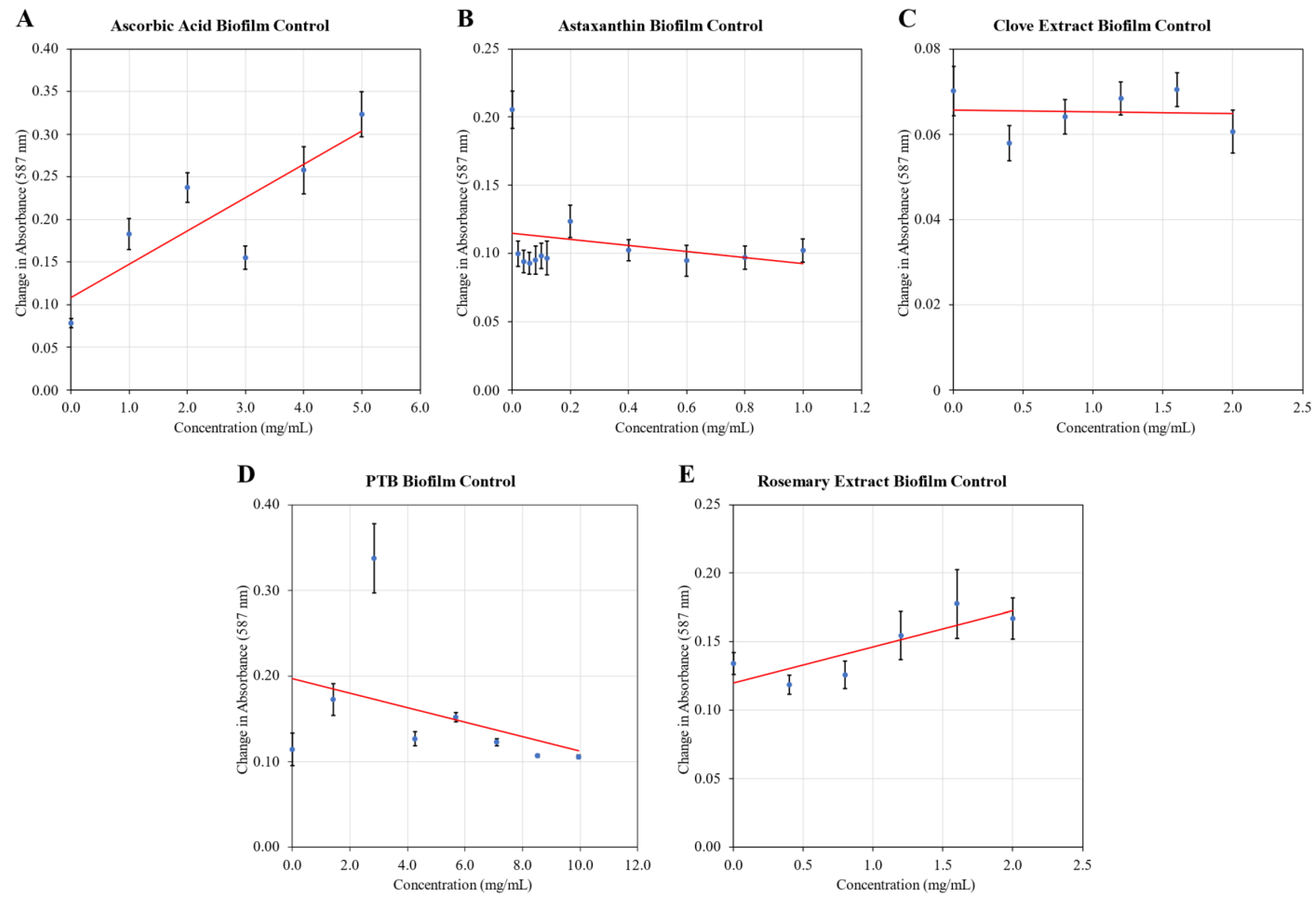
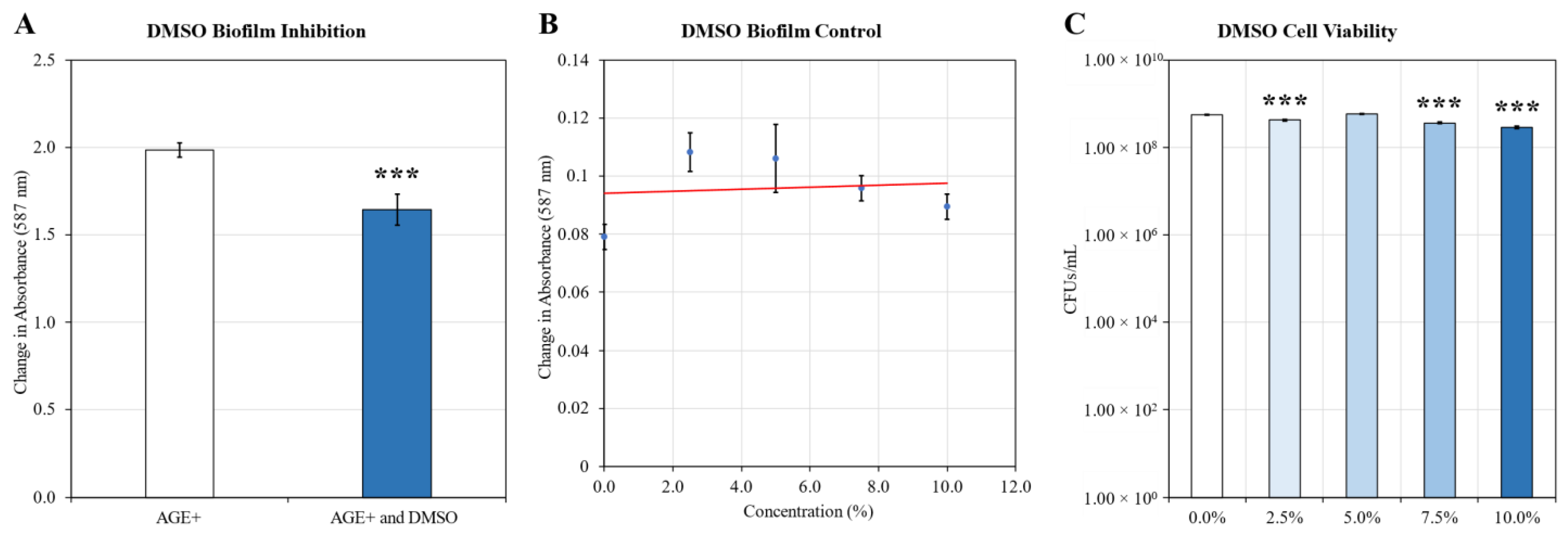
2.1. Biofilm Inhibition
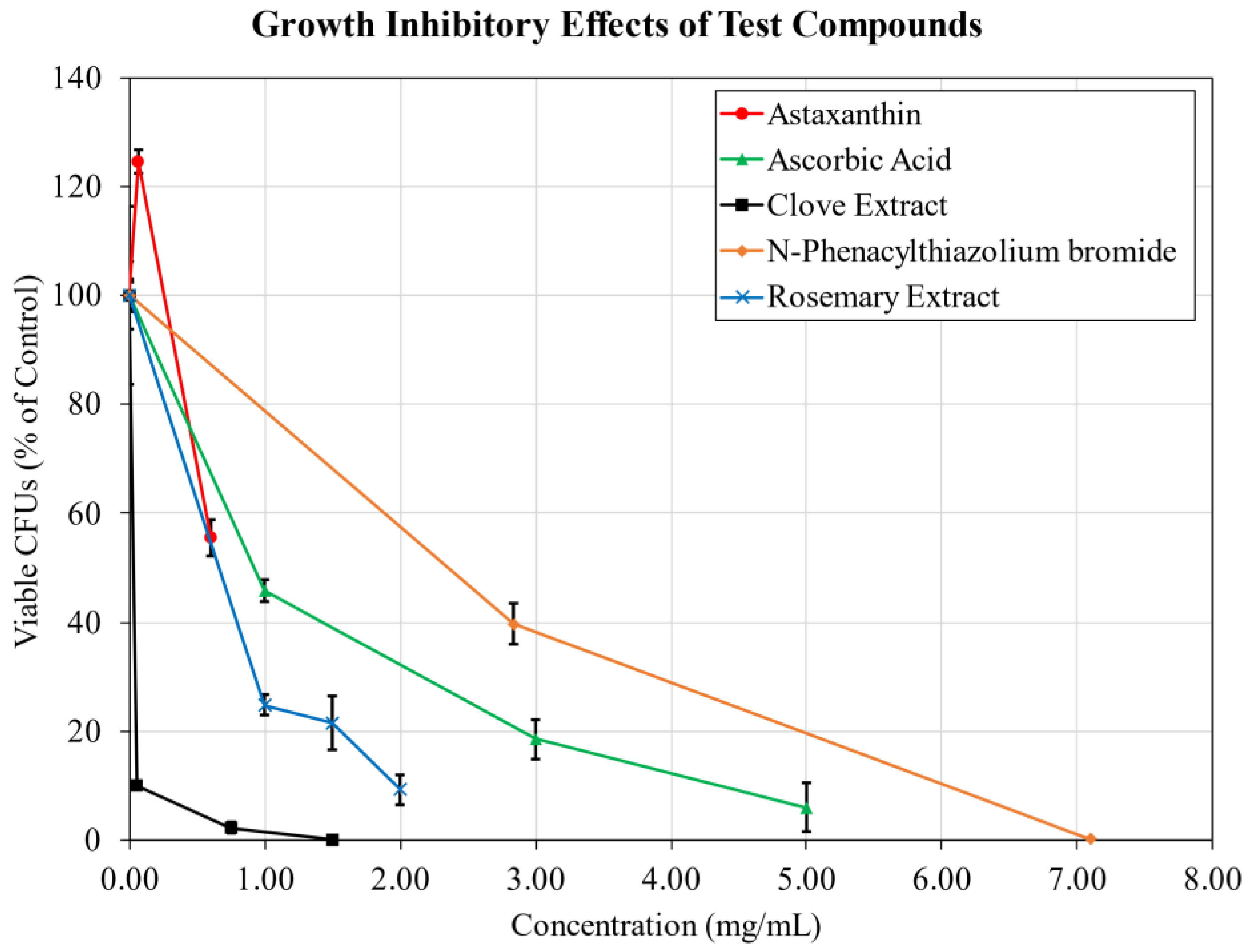
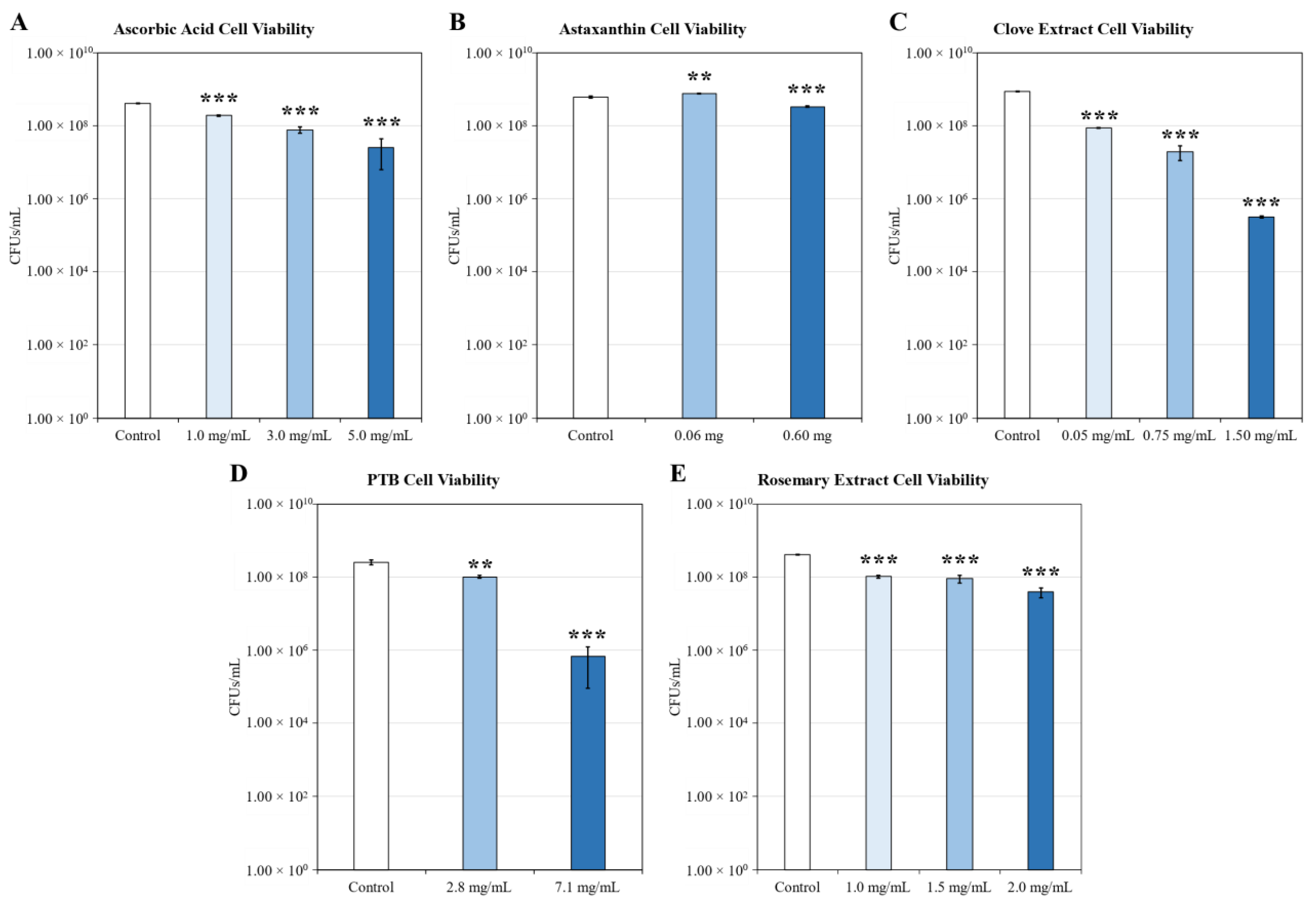
2.2. Growth Inhibition
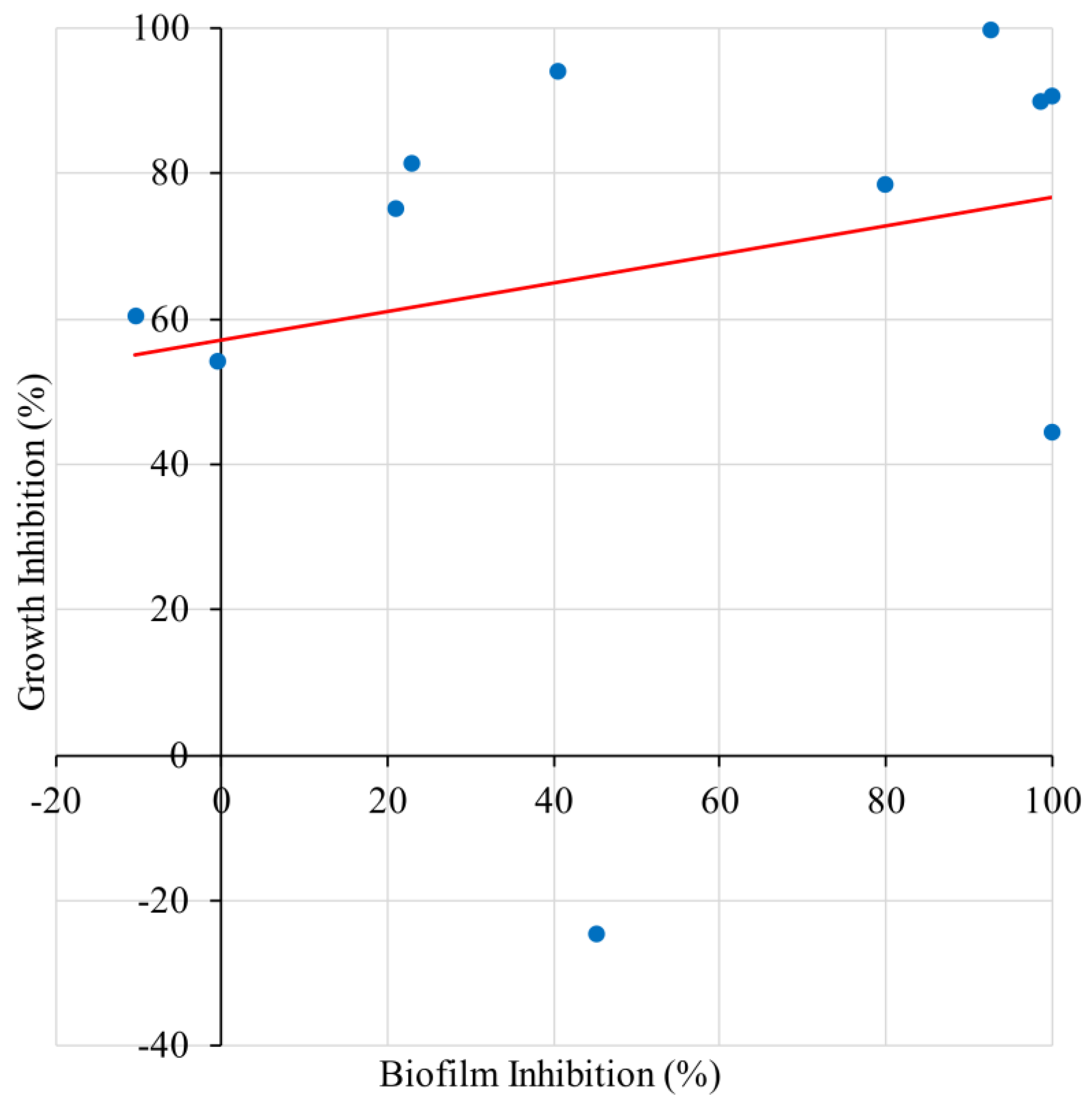
2.3. Correlation of Biofilm Inhibition and Growth Inhibition
3. Discussion
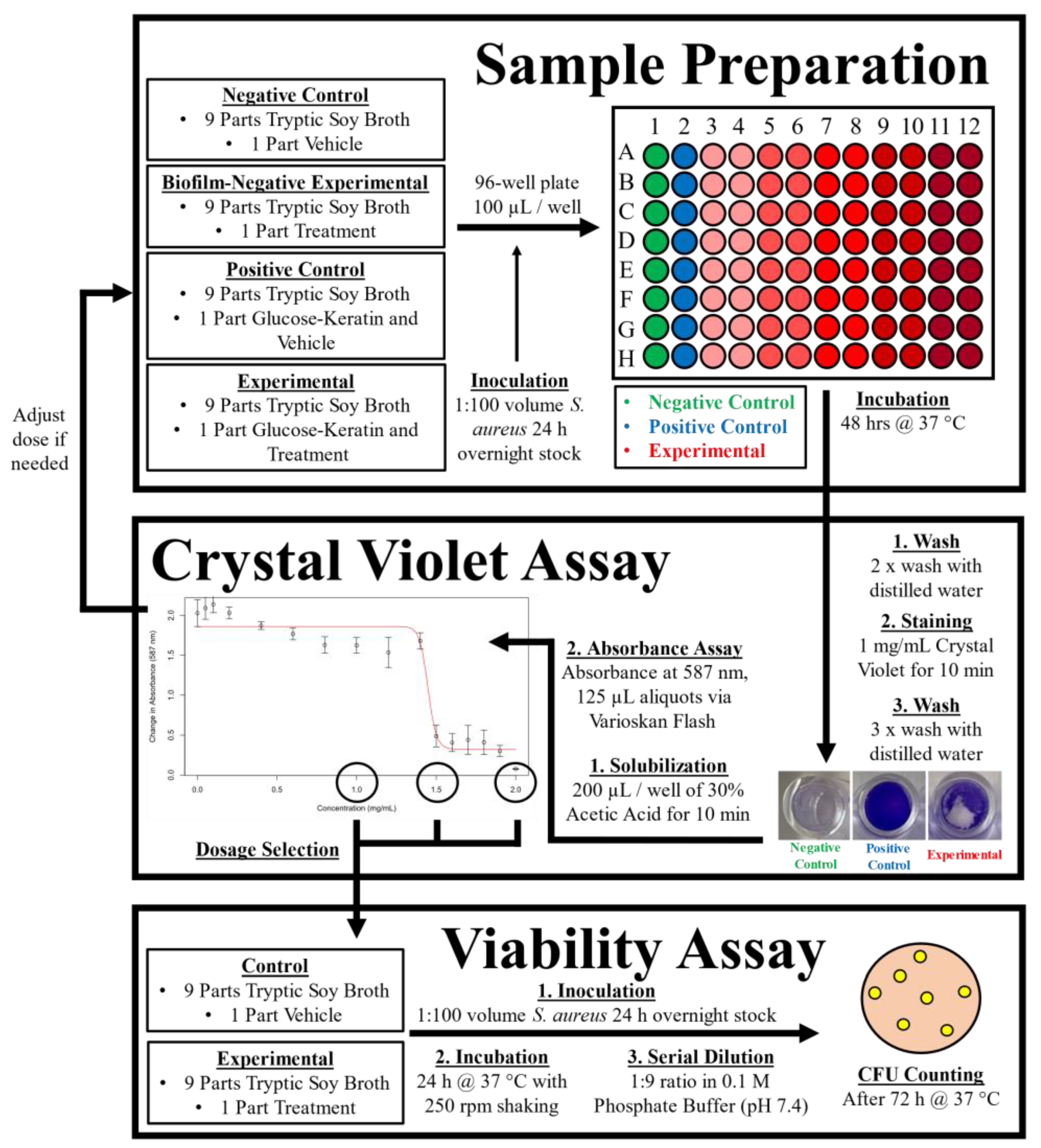
4. Materials and Methods
4.1. Glycation Model
4.2. Bacteria and Growth Conditions
4.3. Biofilm Inhibition Assay
4.4. Cell Viability
4.5. Treatment Compounds
4.6. Logistic Regression Analysis
4.7. Other Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Teissier, T.; Boulanger, É. The receptor for advanced glycation end-products (RAGE) is an important pattern recognition receptor (PRR) for inflammaging. Biogerontology 2019, 20, 279–301. [Google Scholar] [CrossRef] [PubMed]
- Haque, E.; Kamil, M.; Hasan, A.; Irfan, S.; Sheikh, S.; Khatoon, A.; Nazir, A.; Mir, S.S. Advanced glycation end products (AGEs), protein aggregation and their cross talk: New insight in tumorigenesis. Glycobiology 2020, 30, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Genuth, S.; Sun, W.; Cleary, P.; Sell, D.R.; Dahms, W.; Malone, J.; Sivitz, W.; Monnier, V.M. Glycation and Carboxymethyllysine Levels in Skin Collagen Predict the Risk of Future 10-Year Progression of Diabetic Retinopathy and Nephropathy in the Diabetes Control and Complications Trial and Epidemiology of Diabetes Interventions and Complications P. Diabetes 2005, 54, 3103–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rungratanawanich, W.; Qu, Y.; Wang, X.; Essa, M.M.; Song, B.-J. Advanced glycation end products (AGEs) and other adducts in aging-related diseases and alcohol-mediated tissue injury. Exp. Mol. Med. 2021, 53, 168–188. [Google Scholar] [CrossRef]
- Gao, Y.; Wake, H.; Morioka, Y.; Liu, K.; Teshigawara, K.; Shibuya, M.; Zhou, J.; Mori, S.; Takahashi, H.; Nishibori, M. Phagocytosis of Advanced Glycation End Products (AGEs) in Macrophages Induces Cell Apoptosis. Oxidative Med. Cell. Longev. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Touré, F.; Zahm, J.-M.; Garnotel, R.; Lambert, E.; Bonnet, N.; Schmidt, A.M.; Vitry, F.; Chanard, J.; Gillery, P.; Rieu, P. Receptor for advanced glycation end-products (RAGE) modulates neutrophil adhesion and migration on glycoxidated extracellular matrix. Biochem. J. 2008, 416, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Ozer, A.; Altuntas, C.Z.; Izgi, K.; Bicer, F.; Hultgren, S.J.; Liu, G.; Daneshgari, F. Advanced glycation end products facilitate bacterial adherence in urinary tract infection in diabetic mice. Pathog. Dis. 2015, 73, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Birukov, A.; Cuadrat, R.; Polemiti, E.; Eichelmann, F.; Schulze, M.B. Advanced glycation end-products, measured as skin autofluorescence, associate with vascular stiffness in diabetic, pre-diabetic and normoglycemic individuals: A cross-sectional study. Cardiovasc. Diabetol. 2021, 20, 1–11. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Zhang, J.-Q.; Li, L.; Guo, M.-M.; He, Y.-F.; Dong, Y.-M.; Meng, H.; Yi, F. Advanced Glycation End Products in the Skin: Molecular Mechanisms, Methods of Measurement, and Inhibitory Pathways. Front. Med. 2022, 9, 837222. [Google Scholar] [CrossRef]
- Tsukahara, H.; Shibata, R.; Ohta, N.; Sato, S.; Hiraoka, M.; Ito, S.; Noiri, E.; Mayumi, M. High levels of urinary pentosidine, an advanced glycation end product, in children with acute exacerbation of atopic dermatitis: Relationship with oxidative stress. Metabolism. 2003, 52, 1601–1605. [Google Scholar] [CrossRef]
- Hong, J.Y.; Kim, M.J.; Hong, J.K.; Noh, H.H.; Park, K.Y.; Lee, M.K.; Seo, S.J. In vivo quantitative analysis of advanced glycation end products in atopic dermatitis—Possible culprit for the comorbidities? Exp. Dermatol. 2020, 29, 1012–1016. [Google Scholar] [CrossRef]
- Papagrigoraki, A.; Del Giglio, M.; Cosma, C.; Maurelli, M.; Girolomoni, G.; Lapolla, A. Advanced glycation end products are increased in the skin and blood of patients with severe psoriasis. Acta Derm. Venereol. 2017, 97, 782–787. [Google Scholar] [CrossRef] [Green Version]
- Kopeć-Pyciarz, K.; Makulska, I.; Zwolińska, D.; Łaczmański, Ł.; Baran, W. Skin autofluorescence, as a measure of age accumulation in individuals suffering from chronic plaque psoriasis. Mediators Inflamm. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Ogonowska, P.; Gilaberte, Y.; Barańska-Rybak, W.; Nakonieczna, J. Colonization With Staphylococcus aureus in Atopic Dermatitis Patients: Attempts to Reveal the Unknown. Front. Microbiol. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Ng, C.Y.; Huang, Y.H.; Chu, C.F.; Wu, T.C.; Liu, S.H. Risks for Staphylococcus aureus colonization in patients with psoriasis: A systematic review and meta-analysis. Br. J. Dermatol. 2017, 177, 967–977. [Google Scholar] [CrossRef]
- Stacey, H.J.; Clements, C.S.; Welburn, S.C.; Jones, J.D. The prevalence of methicillin-resistant Staphylococcus aureus among diabetic patients: A meta-analysis. Acta Diabetol. 2019, 56, 907–921. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Liu, X.; Li, Y.; Luo, L.; Yuan, W.; Chen, B.; Liang, G.; Shen, R.; Li, H.; Huang, S.; et al. Advanced Glycation End Products Enhance Biofilm Formation by Promoting Extracellular DNA Release Through sigB Upregulation in Staphylococcus aureus. Front. Microbiol. 2020, 11, 1479. [Google Scholar] [CrossRef]
- Haasbroek, K.; Yagi, M.; Yonei, Y. Glycated keratin promotes cellular aggregation and biofilm formation in Staphylococcus aureus. Glycative Stress Res. 2021, 8, 175–182. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Thurlow, L.R.; Hanke, M.L.; Fritz, T.; Angle, A.; Aldrich, A.; Williams, H.; Engebretsen, I.L.; Bayles, K.W.; Horswill, A.R. Staphyloccous auerus biofilms prevent marcophage pahgocytosis and attenuate inflammation in vivo. J. Immunol. 2012, 186, 6585–6596. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; Cavallo, I.; Bordignon, V.; Prignano, G.; Sperduti, I.; Gurtner, A.; Trento, E.; Toma, L.; Pimpinelli, F.; Capitanio, B.; et al. Inflammatory cytokines and biofilm production sustain Staphylococcus aureus outgrowth and persistence: A pivotal interplay in the pathogenesis of Atopic Dermatitis. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, T.; Stevens, M.L.; Kyzy, A.B.; Alarcon, R.; He, H.; Kroner, J.W.; Spagna, D.; Grashel, B.; Sidler, E.; Martin, L.J.; et al. Biofilm propensity of Staphylococcus aureus skin isolates is associated with increased atopic dermatitis severity and barrier dysfunction in the MPAACH pediatric cohort. Allergy Eur. J. Allergy Clin. Immunol. 2021, 76, 302–313. [Google Scholar] [CrossRef]
- Kim, C.-S.; Park, S.; Kim, J. The role of glycation in the pathogenesis of aging and its prevention through herbal products and physical exercise. J. Exerc. Nutr. Biochem. 2017, 21, 55–61. [Google Scholar] [CrossRef]
- Yagi, M.; Yonei, Y. Glycative stress and anti-aging: 13. Regulation of glycative stress. 1. Posprandial blood glucose regulation. Glycative Stress Res. 2019, 6, 175–180. [Google Scholar] [CrossRef]
- Song, Q.; Liu, J.; Dong, L.; Wang, X.; Zhang, X. Novel advances in inhibiting advanced glycation end product formation using natural compounds. Biomed. Pharmacother. 2021, 140, 111750. [Google Scholar] [CrossRef]
- Cooper, M.E.; Thallas, V.; Forbes, J.; Scalbert, E.; Sastra, S.; Darby, I.; Soulis, T. The cross-link breaker, N-phenacylthiazolium bromide prevents vascular advanced glycation end-product accumulation. Diabetologia 2000, 43, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Moores, J. Vitamin C: A wound healing perspective. Br. J. Community Nurs. 2013, 18, S6–S11. [Google Scholar] [CrossRef]
- Padayatty, S.J.; Katz, A.; Wang, Y.; Eck, P.; Kwon, O.; Lee, J.-H.; Chen, S.; Corpe, C.; Dutta, A.; Dutta, S.K.; et al. Vitamin C as an Antioxidant: Evaluation of Its Role in Disease Prevention. J. Am. Coll. Nutr. 2003, 22, 18–35. [Google Scholar] [CrossRef]
- Mousavi, S.; Bereswill, S.; Heimesaat, M.M. Immunomodulatory and antimicrobial effects of vitamin C. Eur. J. Microbiol. Immunol. 2019, 9, 73–79. [Google Scholar] [CrossRef]
- Shivaprasad, D.; Taneja, N.K.; Lakra, A.; Sachdev, D. In vitro and in situ abrogation of biofilm formation in E. coli by vitamin C through ROS generation, disruption of quorum sensing and exopolysaccharide production. Food Chem. 2021, 341, 128171. [Google Scholar] [CrossRef]
- Eydou, Z.; Jad, B.N.; Elsayed, Z.; Ismail, A.; Magaogao, M.; Hossain, A. Investigation on the effect of vitamin C on growth & biofilm-forming potential of Streptococcus mutans isolated from patients with dental caries. BMC Microbiol. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Pandit, S.; Ravikumar, V.; Abdel-Haleem, A.M.; Derouiche, A.; Mokkapati, V.R.S.S.; Sihlbom, C.; Mineta, K.; Gojobori, T.; Gao, X.; Westerlund, F.; et al. Low Concentrations of Vitamin C Reduce the Synthesis of Extracellular Polymers and Destabilize Bacterial Biofilms. Front. Microbiol. 2017, 8, 2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirani, Z.A.; Khan, M.N.; Siddiqui, A.; Khan, F.; Aziz, M.; Naz, S.; Ahmed, A.; Khan, S.I. Ascorbic acid augments colony spreading by reducing biofilm formation of methicillin resistant Staphylococcus aureus. Iran. J. Basic Med. Sci. 2018, 21, 175–180. [Google Scholar] [CrossRef]
- Vinson, J.A.; Howard, T.B. Inhibition of protein glycation and advanced glycation end products by ascorbic acid and other vitamins and nutrients. J. Nutr. Biochem. 1996, 7, 659–663. [Google Scholar] [CrossRef]
- Sztretye, M.; Dienes, B.; Gönczi, M.; Czirják, T.; Csernoch, L.; Dux, L.; Szentesi, P.; Keller-Pintér, A. Astaxanthin: A Potential Mitochondrial-Targeted Antioxidant Treatment in Diseases and with Aging. Oxid. Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Xu, F.H.; Roh, S.-S.; Song, Y.O.; Uebaba, K.; Noh, J.S.; Yokozawa, T. Astaxanthin and Corni Fructus Protect Against Diabetes-Induced Oxidative Stress, Inflammation, and Advanced Glycation End Product in Livers of Streptozotocin-Induced Diabetic Rats. J. Med. Food 2015, 18, 337–344. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, J.; Zeng, X.; Huangfu, J.; Jiang, Y.; Wang, M.; Chen, F. Astaxanthin is responsible for antiglycoxidative properties of microalga Chlorella zofingiensis. Food Chem. 2011, 126, 1629–1635. [Google Scholar] [CrossRef]
- Haasbroek, K.; Takabe, W.; Yagi, M.; Yonei, Y. High-fat Diet Induced Dysbiosis & Amelioration by Astaxanthin. Rad Hrvat. Akad. Znan. i Umjet. Med. Znan. 2019, 49, 58–66. [Google Scholar] [CrossRef]
- Kollerup Madsen, B.; Hilscher, M.; Zetner, D.; Rosenberg, J. Adverse reactions of dimethyl sulfoxide in humans: A systematic review. F1000Research 2018, 7, 1746. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, B.; Wang, Y. Clove essential oil confers antioxidant activity and lifespan extension in C. elegans via the DAF-16/FOXO transcription factor. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 242, 108938. [Google Scholar] [CrossRef]
- Han, X.; Parker, T.L. Anti-inflammatory activity of clove ( Eugenia caryophyllata ) essential oil in human dermal fibroblasts. Pharm. Biol. 2017, 55, 1619–1622. [Google Scholar] [CrossRef] [Green Version]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An Overview on the Anti-inflammatory Potential and Antioxidant Profile of Eugenol. Oxid. Med. Cell Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef]
- Nirmala, M.J.; Durai, L.; Gopakumar, V.; Nagarajan, R. Anticancer and antibacterial effects of a clove bud essential oil-based nanoscale emulsion system. Int. J. Nanomed. 2019, 14, 6439–6450. [Google Scholar] [CrossRef] [Green Version]
- Kubatka, P.; Uramova, S.; Kello, M.; Kajo, K.; Kruzliak, P.; Mojzis, J.; Vybohova, D.; Adamkov, M.; Jasek, K.; Lasabova, Z.; et al. Antineoplastic effects of clove buds (Syzygium aromaticum L.) in the model of breast carcinoma. J. Cell. Mol. Med. 2017, 21, 2837–2851. [Google Scholar] [CrossRef]
- Nuñez, L.; D’Aquino, M. Microbicide activity of clove essential oil (Eugenia caryophyllata). Brazilian J. Microbiol. 2012, 43, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of clove (Syzygium aromaticum) oil and its major components to human skin cells. Cell Prolif. 2006, 39, 241–248. [Google Scholar] [CrossRef]
- Aldini, G.; Vistoli, G.; Stefek, M.; Chondrogianni, N.; Grune, T.; Sereikaite, J.; Sadowska-Bartosz, I.; Bartosz, G. Molecular strategies to prevent, inhibit, and degrade advanced glycoxidation and advanced lipoxidation end products. Free Radic. Res. 2013, 47, 93–137. [Google Scholar] [CrossRef] [Green Version]
- Hollenbach, S.; Thampi, P.; Viswanathan, T.; Abraham, E.C. Cleavage of in vitro and in vivo formed lens protein cross-links by a novel cross-link breaker. Mol. Cell. Biochem. 2003, 243, 73–80. [Google Scholar] [CrossRef]
- Chang, P.-C.; Tsai, S.-C.; Chong, L.Y.; Kao, M.-J. N-Phenacylthiazolium Bromide Inhibits the Advanced Glycation End Product (AGE)–AGE Receptor Axis to Modulate Experimental Periodontitis in Rats. J. Periodontol. 2014, 85, e268–e276. [Google Scholar] [CrossRef]
- Bradke, B.S.; Vashishth, D. N-Phenacylthiazolium Bromide Reduces Bone Fragility Induced by Nonenzymatic Glycation. PLoS ONE 2014, 9, e103199. [Google Scholar] [CrossRef]
- Nieto, G.; Ros, G.; Castillo, J. Antioxidant and antimicrobial properties of rosemary (Rosmarinus officinalis, L.): A review. Medicines 2018, 5, 98. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Han, J.; Zheng, X.; Ai, B.; Yang, Y.; Xiao, D.; Zheng, L.; Sheng, Z. Rosemary leaf extract inhibits glycation, breast cancer proliferation, and diabetes risks. Appl. Sci. 2020, 10, 2249. [Google Scholar] [CrossRef] [Green Version]
- Jean, D.; Pouligon, M.; Dalle, C. Evaluation in vitro of AGE-crosslinks breaking ability of rosmarinic acid. Glycative Stress Res. 2015, 2, 204–207. [Google Scholar]
- Hori, M.; Yagi, M.; Nomoto, K.; Ichijo, R.; Shimode, A.; Kitano, T.Y. Experimental models for advanced glycation end product formation using albumin, collagen, elastin, keratin and proteoglycan. Anti-Aging Med. 2012, 9, 125–134. [Google Scholar]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and Analyzing Static Biofilms. In Current Protocols in Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005. [Google Scholar]
- Shay, R.; Wiegand, A.A.; Trail, F. Biofilm Formation and Structure in the Filamentous Fungus Fusarium graminearum, a Plant Pathogen. Microbiol. Spectr. 2022, 10, e0017122. [Google Scholar] [CrossRef]
- Albaghdadi, S.Z.; Altaher, J.B.; Drobiova, H.; Bhardwaj, R.G.; Karched, M. In vitro Characterization of Biofilm Formation in Prevotella Species. Front. Oral Health 2021, 2, 724194. [Google Scholar] [CrossRef]
- Suantawee, T.; Wesarachanon, K.; Anantsuphasak, K.; Daenphetploy, T.; Thien-Ngern, S.; Thilavech, T.; Pasukamonset, P.; Ngamukote, S.; Adisakwattana, S. Protein glycation inhibitory activity and antioxidant capacity of clove extract. J. Food Sci. Technol. 2014, 52, 3843–3850. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing R Foundation for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | ED50 (mg/mL) | Max Inhibition (% Control) |
|---|---|---|
| Ascorbic Acid | 1.9 | 40.5 |
| Astaxanthin | 0.060 | 100.0 |
| Clove Extract | 0.0087 | 98.6 |
| n-Phenacylthiazolium Bromide | 5.3 | 92.6 |
| Rosemary Extract (AGE Breaker) | 1.5 | 100.0 |
| Material | Manufacturer | Antimicrobial Activity | AGE Crosslink Breaking | Glycation Inhibition | Anti-Oxidant Activity |
|---|---|---|---|---|---|
| Ascorbic Acid | Wako Chemical (Osaka, Japan) | + [31,32] | − | + [25,34] | + [28] |
| Astaxanthin | Sigma-Aldrich (St. Louis, MO, USA) | − | − | + [36,37] | + [35] |
| Clove Extract | House Foods (Osaka, Japan) | + [43,45] | − | + [58] | + [40] |
| n-Phenacylthiazolium Bromide | Fluorochem (Hadfield, UK) | − | + [26,48] | − | − |
| Rosemary Extract (AGE Breaker) | A2P Sciences (Lyon, France) | + [51] | + [53] | + [52] | + [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haasbroek, K.; Yagi, M.; Yonei, Y. Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds. Antibiotics 2022, 11, 1412. https://doi.org/10.3390/antibiotics11101412
Haasbroek K, Yagi M, Yonei Y. Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds. Antibiotics. 2022; 11(10):1412. https://doi.org/10.3390/antibiotics11101412
Chicago/Turabian StyleHaasbroek, Kyle, Masayuki Yagi, and Yoshikazu Yonei. 2022. "Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds" Antibiotics 11, no. 10: 1412. https://doi.org/10.3390/antibiotics11101412
APA StyleHaasbroek, K., Yagi, M., & Yonei, Y. (2022). Staphylococcus aureus Biofilm Inhibiting Activity of Advanced Glycation Endproduct Crosslink Breaking and Glycation Inhibiting Compounds. Antibiotics, 11(10), 1412. https://doi.org/10.3390/antibiotics11101412