
Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal

Abstract
:1. Introduction
2. Results
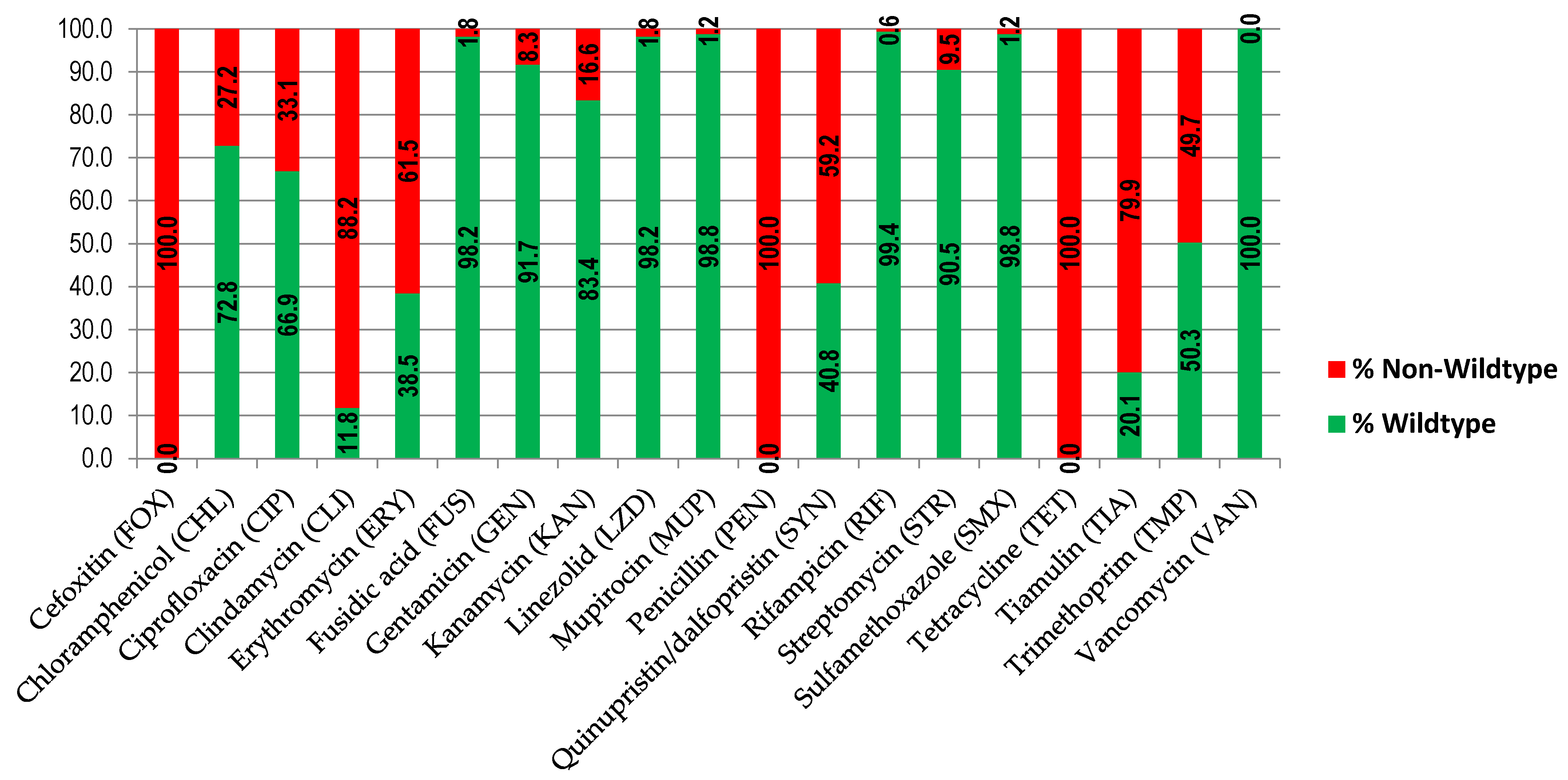
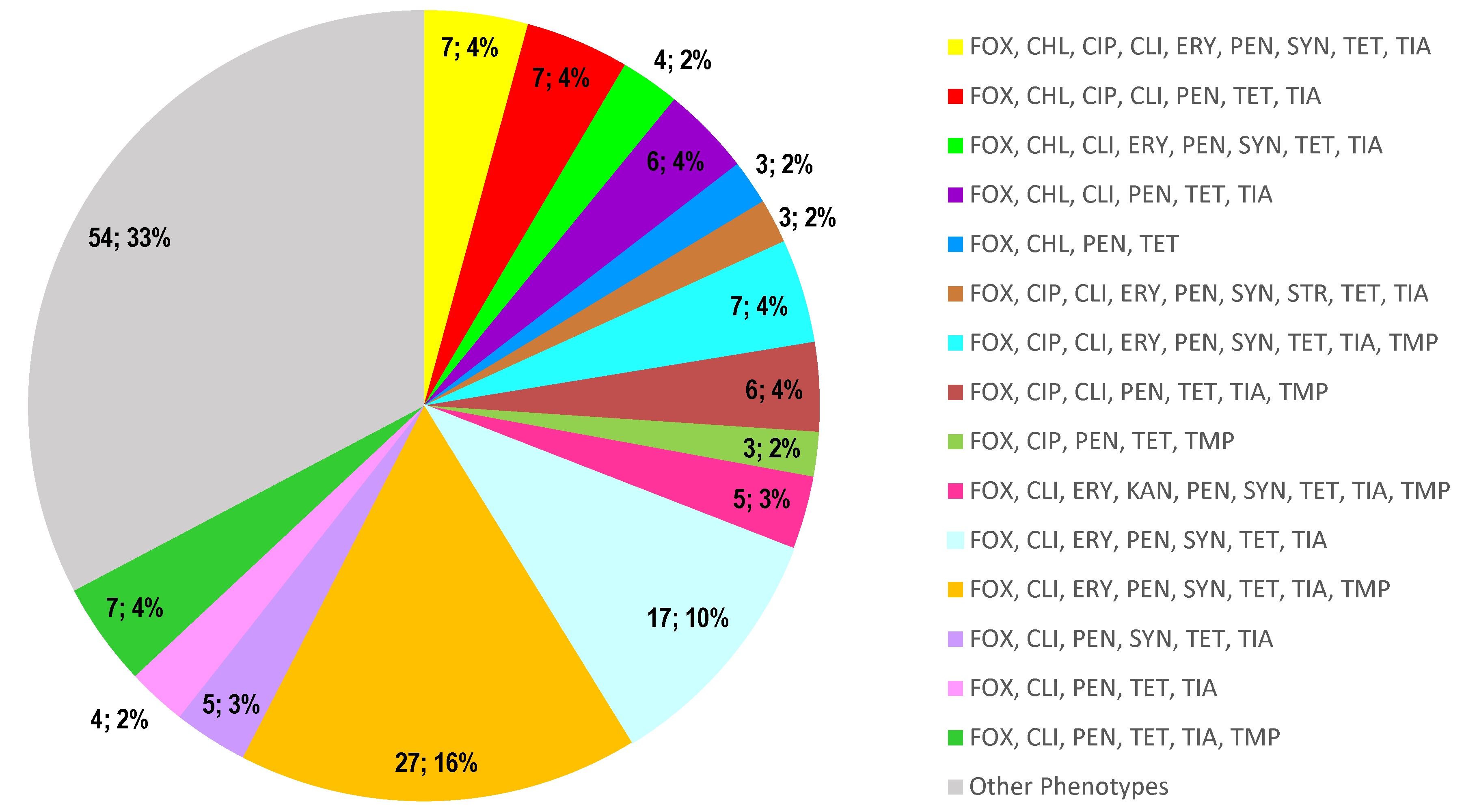
2.1. Bacterial Isolates and Antimicrobial Resistance Patterns
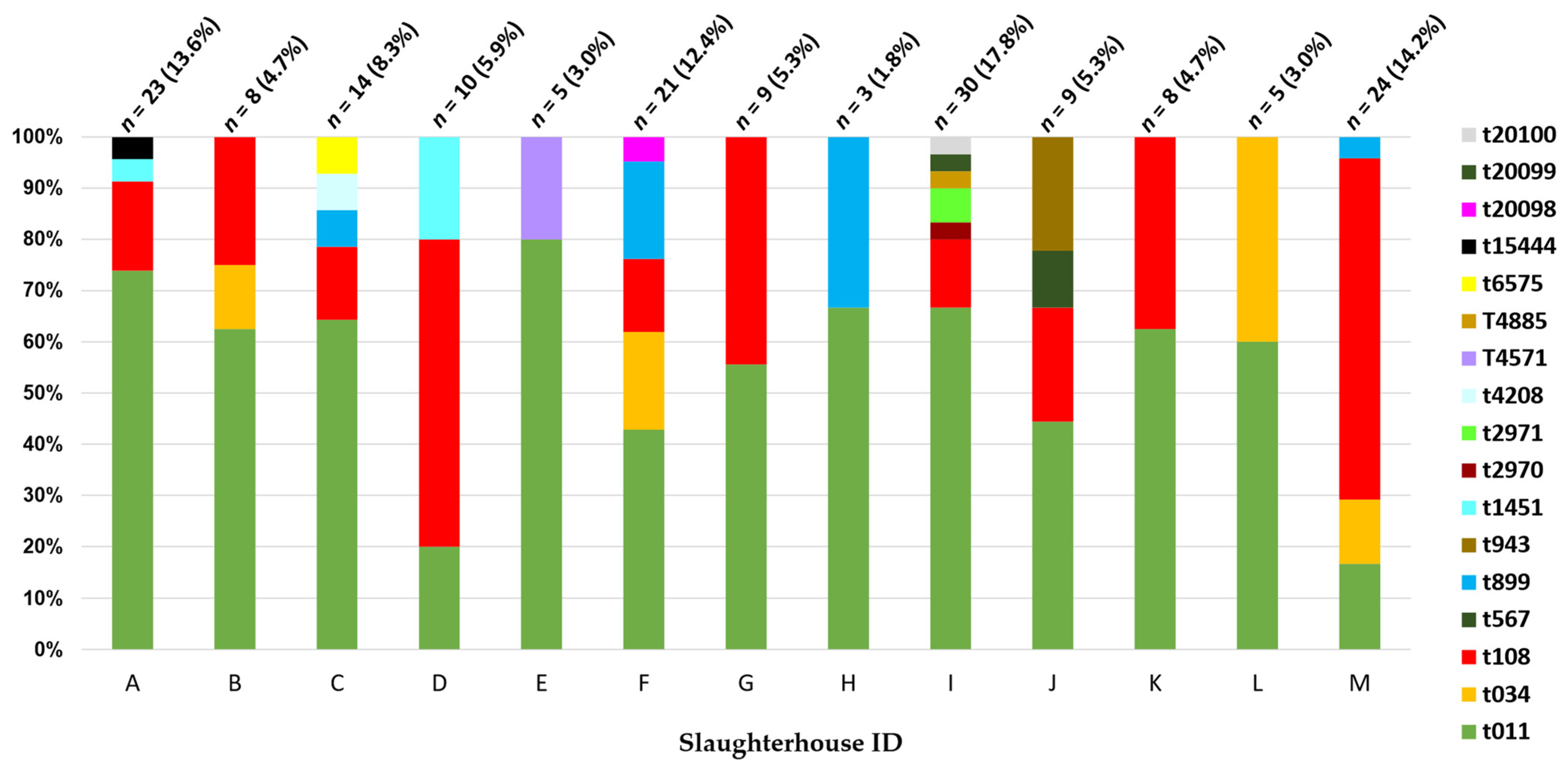
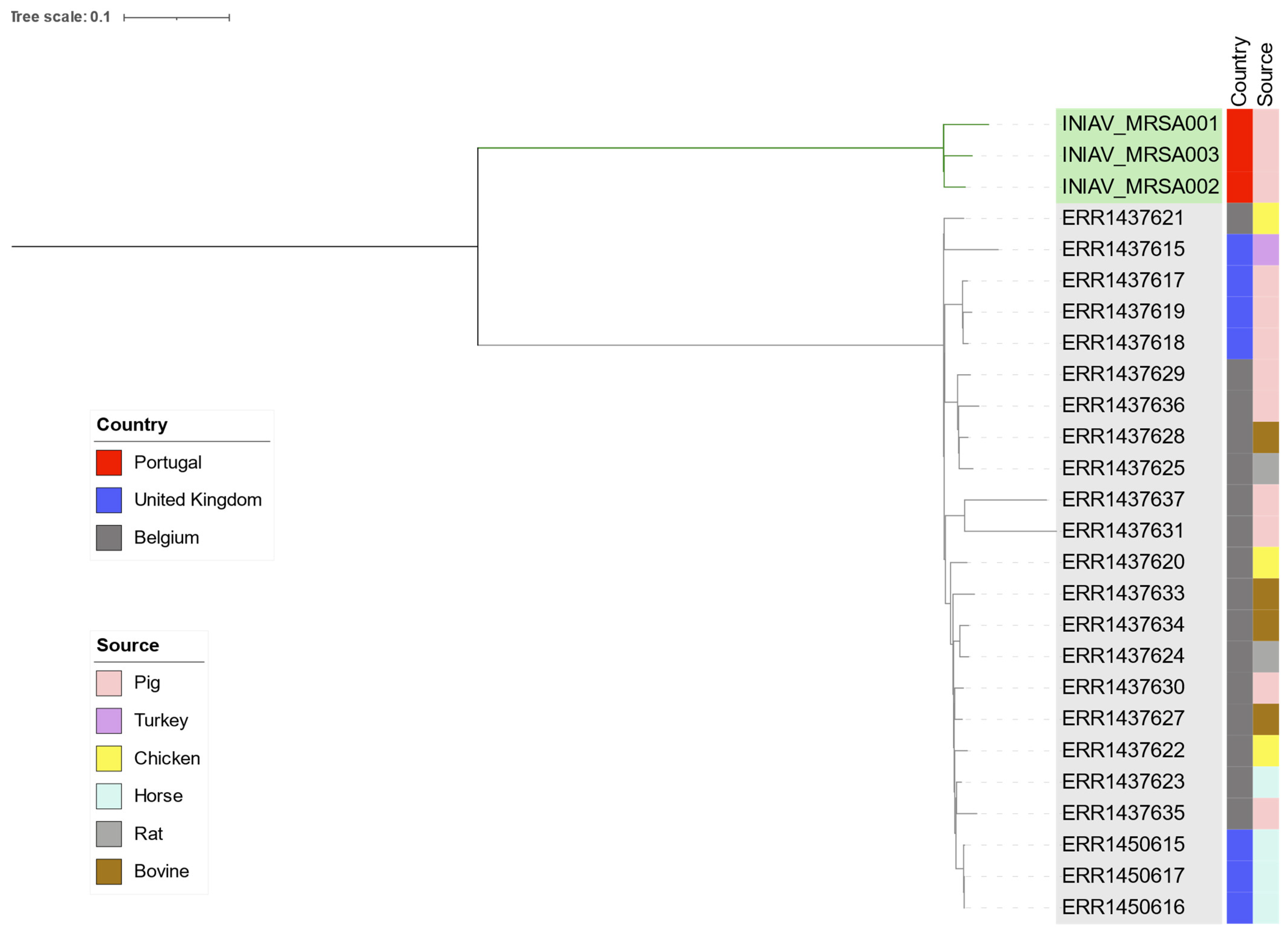
2.2. Molecular Characterization of the MRSA Population
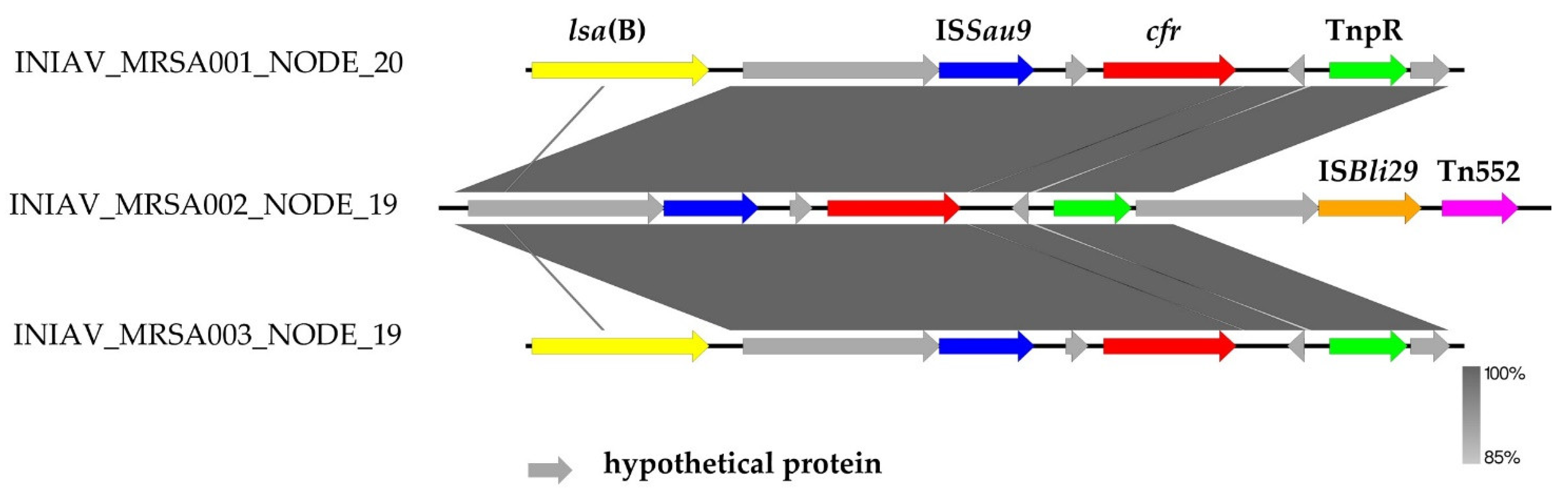
2.3. Whole-Genome Characterization of Linezolid-Resistant MRSA
3. Discussion
4. Materials and Methods
4.1. Sampling and Bacterial Isolation
4.2. Antimicrobial Susceptibility Testing
4.3. Molecular Characterization of the Isolates
4.4. Whole-Genome Sequencing of Linezolid-Resistant Isolates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loncarica, I.; Lepuschitzb, S.; Ruppitschb, W.; Trstana, A.; Andreadisa, T.; Bouchlisa, N.; Marbacha, H.; Schauera, B.; Szostaka, M.; Feßlerc, A.; et al. Increased genetic diversity of methicillin-resistant Staphylococcus aureus (MRSA) isolated from companion animals. Vet. Microbiol. 2019, 235, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Ronco, T.; Stegger, M.; Sieber, R.; Fertner, M.; Martin, M.; Farre Toft, N.; Larsen, A.; Pedersen, K. LA-MRSA CC398 in Dairy Cattle and Veal Calf Farms Indicates Spillover From Pig Production. Front. Microbiol. 2019, 10, 2733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição, T.; Lencastre, H.; Aires-de-Sousa, M. Frequent isolation of methicillin resistant Staphylococcus aureus (MRSA) ST398 among healthy pigs in Portugal. PLoS ONE 2017, 12, e0175340. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Belas, A.; Kadlec, K.; Schwarz, S.; Pomba, S. Clonal diversity, virulence patterns and antimicrobial and biocide susceptibility among human, animal and environmental MRSA in Portugal. J. Antimicrob. Chemother. 2015, 70, 2483–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, V.; Vieira-Pinto, M.; Saraiva, C.; Manageiro, V.; Reis, L.; Ferreira, E.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Prevalence and Characteristics of Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) CC398 Isolated from Quails (Coturnix Coturnix Japonica) Slaughtered for Human Consumption. Animals 2021, 11, 2038. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.; Silva, N.; Borges, V.; Gomes, J.; Vieira, L.; Caniça, M.; Torres, C.; Igrejas, G.; Poeta, P. MRSA CC398 recovered from wild boar harboring new SCCmec type IV J3 variant. Sci. Total Environ. 2020, 722, 137845. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A. Staphylococcus aureus. In Molecular Medical Microbiology; Tang, Y.W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Elsevier Ltd.: Dublin, Ireland, 2015; pp. 655–674. ISBN 3522738284. [Google Scholar]
- Quinn, P.J.; Carter, M.E.; Markey, B.; Carter, G.R. Staphylococcus species. In Clinical Veterinary Microbiology; Mosby International Limited: London, UK, 2002; pp. 118–126. ISBN 0723417113. [Google Scholar]
- Voss, A.; Loeffen, F.; Bakker, J.; Klaassen, C.; Wulf, M. Methicillin-resistant Staphylococcus aureus in pig farming. Emerg. Infect. Dis. 2005, 11, 1965–1966. [Google Scholar] [CrossRef]
- Kinross, P.; Petersen, A.; Skov, R.; Van Hauwermeiren, E.; Pantosti, A.; Laurent, F.; Voss, A.; Kluytmans, J.; Struelens, M.J. The European Human LA-MRSA Study Group; et al. Livestock-associated methicillin-resistant Staphylococcus aureus (MRSA) among human MRSA isolates. European Union/European Economic Area countries. Eurosurveillance 2017, 22, 16-00696. [Google Scholar] [CrossRef] [Green Version]
- Pirolo, M.; Visaggio, D.; Gioffrè, A.; Artuso, I.; Gherardi, M.; Pavia, G.; Samele, P.; Ciambrone, L.; Di Natale, R.; Spatari, G.; et al. Unidirectional animal-to-human transmission of methicillin-resistant Staphylococcus aureus ST398 in pig farming; evidence from a surveillance study in southern Italy. Antimicrob. Resist. Infect. Control 2019, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Fessler, A.T.; Hauschild, T.; Schwarz, S. Novel and uncommon antimicrobial resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 745–755. [Google Scholar] [CrossRef]
- Hashemian, S.M.R.; Farhadi, T.; Ganjparvar, M. Linezolid: A review of its properties, function, and use in critical care. Drug Des. Dev. Ther. 2018, 12, 1759–1767. [Google Scholar] [CrossRef]
- Silva, V.; Almeida, F.; Silva, A.; Correia, S.; Carvalho, J.A.; Castro, A.P.; Ferreira, E.; Manageiro, V.; Caniça, M.; Igrejas, G.; et al. First report of linezolid-resistant cfr-positive methicillin-resistant Staphylococcus aureus in humans in Portugal. J. Glob. Antimicrob. Resist. 2019, 17, 323–325. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2018/2019. EFSA J. Wiley Online Libr. 2021, 19, e06490. [Google Scholar] [CrossRef]
- Seixas, R.; Monteiro, V.; Carneiro, C.; Vilela, C.L.; Oliveira, M. First report of a linezolid-resistant MRSA (methicillin resistant Staphylococcus aureus) isolated from a dog with a severe bilateral otitis in Portugal. Rev. Vet. 2011, 22, 81–84. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST). Available online: https://www.eucast.org/mic_distributions_and_ecoffs/ (accessed on 23 February 2022).
- Ridom SpaServer Platform. Available online: https://spa.ridom.de/submission.shtml (accessed on 30 July 2021).
- Lopes, E.; Conceição, T.; Poirel, L.; de Lencastre, H.; Aires-de-Sousa, M. Epidemiology and antimicrobial resistance of methicillin-resistant Staphylococcus aureus isolates colonizing pigs with different exposure to antibiotics. PLoS ONE. 2019, 14, e0225497. [Google Scholar] [CrossRef] [Green Version]
- Bouchami, O.; Fraqueza, M.J.; Faria, N.A.; Alves, V.; Lawal, O.U.; de Lencastre, H.; Miragaia, M. Evidence for the Dissemination to Humans of Methicillin-Resistant Staphylococcus aureus ST398 through the Pork Production Chain: A Study in a Portuguese Slaughterhouse. Microorganisms 2020, 8, 1892. [Google Scholar] [CrossRef]
- European Food Safety Authority. Analysis of the baseline survey on the prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in holdings with breeding pigs, in the EU. EJ EFSA J. Wiley Online Libr. 2008, 7, 1376. [Google Scholar] [CrossRef]
- Khaled, R.A.; Suriya, R.; Amani, A.; Asim, D.; Abbas, H.; Sima, T. Molecular typing of MRSA isolates by spa and PFGE. J. King Saud. Univ. Sci. 2019, 31, 999–1004. [Google Scholar] [CrossRef]
- Veterinary Medicines Division. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2018 Trends from 2010 to 2018 Tenth ESVAC Report; EMA/24309/2020; Veterinary Medicines Division: Luxembourg, 2020. [Google Scholar]
- Kehrenberg, C.; Aarestrup, F.M.; Schwarz, S. IS21-558 insertion sequences are involved in the mobility of the multiresistance gene cfr. Antimicrob. Agents Chemother. 2007, 51, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Zhang, W.; Du, X.-D.; Krüger, H.; Feßler, A.T.; Ma, S.; Zhu, Y.; Wu, C.; Shen, J.; Wang, Y. Mobile oxazolidinone resistance genes in Gram-positive and Gram-negative bacteria. Clin. Microbiol. Rev. 2021, 34, e00188-20. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [Green Version]
- Peeters, L.E.J.; Argudín, M.A.; Azadikhah, S.; Butaye, P. Antimicrobial resistance and population structure of Staphylococcus aureus recovered from pigs farms. Vet. Microbiol. 2015, 180, 151–156. [Google Scholar] [CrossRef]
- Kang, H.Y.; Moon, D.C.; Mechesso, A.F.; Choi, J.H.; Kim, S.J.; Song, H.J.; Kim, M.H.; Yoon, S.S.; Lim, S.K. Emergence of cfr-Mediated Linezolid Resistance in Staphylococcus aureus Isolated from Pig Carcasses. Antibiotics 2020, 9, 769. [Google Scholar] [CrossRef]
- Li, S.M.; Zhou, Y.F.; Li, L.; Fang, L.X.; Duan, J.H.; Liu, F.R.; Liang, H.Q.; Wu, Y.T.; Gu, W.Q.; Liao, X.P.; et al. Characterization of the Multi-Drug Resistance Gene cfr in Methicillin-Resistant Staphylococcus aureus (MRSA) Strains Isolated From Animals and Humans in China. Front. Microbiol. 2018, 9, 2925. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Bellés-Bellés, A.; Fernández-Fernández, R.; García, M.; Vilaró, A.; Zarazaga, M.; Torres, C. Linezolid-resistant MRSA-CC398 carrying the cfr gene, and MRSA-CC9 isolates from pigs with signs of infection in Spain. J. Appl. Microbiol. 2021, 131, 615–622. [Google Scholar] [CrossRef]
- Antonelli, A.; D’Andrea, M.M.; Galano, A.; Borchi, B.; Brenciani, A.; Vaggelli, G.; Cavallo, A.; Bartoloni, A.; Giovanetti, E.; Rossolini, G.M. Linezolid-resistant cfr-positive MRSA, Italy. J. Antimicrob. Chemother. 2016, 71, 2349–2351. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ripa, L.; Bellés, A.; García, M.; Torres, C. Detection of a cfr-positive MRSA CC398 strain in a pig farmer in Spain. Enferm. Infecc. Microbiol. Clin. Engl. Ed. 2021, 39, 139–141, (In English and Spanish). [Google Scholar] [CrossRef]
- Morales, G.; Picazo, J.J.; Baos, E.; Candel, F.J.; Arribi, A.; Peláez, B.; Andrade, R.; de la Torre, M.A.; Fereres, J.; Sánchez-García, M. Resistance to linezolid is mediated by the cfr gene in the first report of an outbreak of linezolid-resistant Staphylococcus aureus. Clin. Infect. Dis. 2010, 50, 821–825. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Chen, L.; Xie, Z.; Zhang, M. Dissemination of cfr-mediated linezolid resistance among Staphylococcus species isolated from a teaching hospital in Beijing, China. J. Int. Med. Res. 2018, 46, 3884–3889. [Google Scholar] [CrossRef] [Green Version]
- Couto, N.; Belas, A.; Rodrigues, C.; Schwarz, S.; Pomba, C. Acquisition of the fexA and cfr genes in Staphylococcus pseudintermedius during florfenicol treatment of canine pyoderma. J. Glob. Antimicrob. Resist. 2016, 7, 126–127. [Google Scholar] [CrossRef]
- Coll, F.; Raven, K.E.; Knight, G.M.; Blane, B.; Harrison, E.M.; Leek, D.; Enoch, D.A.; Brown, N.M.; Parkhill, J.; Peacock, S.J. Definition of a genetic relatedness cutoff to exclude recent transmission of meticillin-resistant Staphylococcus aureus: A genomic epidemiology analysis. Lancet Microbe 2020, 1, e328–e335. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Gião, J.; Leão, C.; Albuquerque, T.; Clemente, L.; Amaro, A. Antimicrobial Susceptibility of Enterococcus Isolates from Cattle and Pigs in Portugal: Linezolid Resistance Genes optrA and poxtA. Antibiotics 2022, 11, 615. [Google Scholar] [CrossRef] [PubMed]
- EURL-AR. European Reference Laboratory for Antimicrobial Resistance. Protocols. Available online: https://www.eurl-ar.eu/CustomerData/Files/Folders/21-protocols/430_mrsa-protocol-final-19-06-2018.pdf (accessed on 23 February 2022).
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecA(LGA251). Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Shopsin, B.; Gomez, M.; Montgomery, S.O.; Smith, D.H.; Waddington, M.; Dodge, D.E.; Bost, D.A.; Riehman, M.; Naidich, S.; Kreiswirth, B.N. Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 1999, 37, 3556–3563. [Google Scholar] [CrossRef] [Green Version]
- European Nucleotide Archive (ENA). Available online: https://www.ebi.ac.uk/ena/submit/sra/#home (accessed on 16 March 2021).
- Andrews, S. FastQC: A quality control tool for high throughput sequence data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 19 January 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling genomes and mini-metagenomes from highly chimeric reads. In Proceedings of the Research in Computational Molecular Biology: 17th Annual International Conference (RECOMB), Beijing, China, 7–10 April 2013; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 158–170. Available online: http://link.springer.com/chapter/10.1007/978-3-642-37195-0_13 (accessed on 15 June 2021).
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.F.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.R.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Larsen, M.; Lund, O.; Voldby Villa, L.; Møller Aarestrup, F.; Hasman, H. PlasmidFinder and pMLST: In silico detection and typing of plasmids. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontéen, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total Genome Sequenced Bacteria. J. Clin. Micobiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Micobiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing whole-genome sequencing with Sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- International Working Group on The Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC). Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 56, 4961–4967. [Google Scholar]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr and major difference in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Nunez-Garcia, J.; Kearns, A.M.; Doumith, M.; Butaye, P.R.; Argudín, M.A.; Lahuerta-Marin, A.; Pichon, B.; AbuOun, M.; Rogers, J.; et al. Livestock-Associated Methicillin Resistant Staphylococcus aureus (LA-MRSA) Clonal Complex (CC) 398 Isolated from UK Animals belong to European Lineages. Front. Microbiol. 2016, 7, 1741. [Google Scholar] [CrossRef] [Green Version]
- Ciccarelli, F.D.; Doerks, T.; von Mering, C.; Creevey, C.J.; Snel, B.; Bork, P. Toward automatic reconstruction of a highly resolved tree of life. Science 2006, 311, 1283–1287. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | Clinical MIC Breakpoints [17] (mg/L) | Epidemiological MIC Breakpoints [17] (mg/L) (T) | MIC50 (mg/L) (n = 169) | MIC90 (mg/L) (n = 169) | % DS (N) |
|---|---|---|---|---|---|
| Cefoxitin (FOX) | 4 | NA | >16 | >16 | 100.0 (169) |
| Chloramphenicol (CHL) | 8 | NA | 16 | 64 | 27.2 (46) |
| Ciprofloxacin (CIP) | 1 | 1 | 0.5 | >8 | 33.1 (56) |
| Clindamycin (CLI) | 0.25 | 0.25 | >4 | >4 | 88.2 (149) |
| Erythromycin (ERY) | 2 | 1 | >8 | >8 | 61.5 (104) |
| Fusidic acid (FUS) | 1 | 0.5 | ≤0.5 | ≤0.5 | 1.8 (3) |
| Gentamicin (GEN) | 2 | (2) | ≤1 | ≤1 | 8.3 (14) |
| Kanamycin (KAN) | 8 | NA | ≤4 | 32 | 16.6 (28) |
| Linezolid (LZD) | 4 | 4 | 2 | 4 | 1.8 (3) |
| Mupirocin (MUP) | NA | (1) | ≤0.5 | ≤0.5 | 1.2 (2) |
| Penicillin (PEN) | 0.125 | NA | >2 | >2 | 100.0 (169) |
| Quinupristin/dalfopristin (SYN) | 2 | NA | 2 | 4 | 59.2 (100) |
| Rifampicin (RIF) | 0.06 | (0.03) | ≤0.016 | ≤0.016 | 0.6 (1) |
| Streptomycin (STR) | NA | NA | 8 | 16 | 9.5 (16) |
| Sulfamethoxazole (SMX) | NA | NA | ≤64 | ≤64 | 1.2 (2) |
| Tetracycline (TET) | 2 | 1 | >16 | >16 | 100.0 (169) |
| Tiamulin (TIA) | NA | NA | >4 | >4 | 79.9 (135) |
| Trimethoprim (TMP) | NA | (2) | ≤2 | >32 | 49.7 (84) |
| Vancomycin (VAN) | 2 | 2 | ≤1 | ≤1 | 0.0 (0) |
| Feature | INIAV_MRSA001 | INIAV_MRSA002 | INIAV_MRSA003 |
|---|---|---|---|
| Sampling date | October 22 | November 7 | November 20 |
| Farm region | AML (Peninsula de Setubal) | Alentejo | Alentejo Litoral |
| Slaughterhouse ID (region) | A (AML, Península de Setúbal) | K (North region, Ave) | K (North region, Ave) |
| Phenotype | FOX, CHL, CLI, ERY, LZD, PEN, SYN, TET, TIA, TMP | FOX, CHL, CLI, ERY, LZD, PEN, SYN, TET, TIA, TMP | FOX, CHL, CLI, LZD, PEN, SYN, TET, TIA, TMP |
| Antibiotic resistance genes | aadD, blaZ, mecA, lsa(B), cfr, fexA, tet(M), tet(L), tet(K) | blaZ, mecA, vga(A)LC, cfr, fexA, tet(M), tet(K), dfrG | blaZ. mecA, erm(B), Isa(B), cfr, vga(A)LC, fexA, tet(M), tet(L), tet(K), dfrK |
| Biocide resistance genes | norA, lmrS, mepA, sepA | norA, lmrS, mepA, sepA | norA, lmrS, mepA, sepA |
| SCCmec elements | SCCmec_type_Vc(5C2&5) | SCCmec_type_Vc(5C2&5) | SCCmec_type_Vc(5C2&5) |
| Virulence genes | aur; hlgA; hlgB; hlgC | aur; hlgA; hlgB; hlgC | aur; hlgA; hlgB; hlgC |
| Plasmid replicons | rep21, rep22, rep7a, repUS43 | rep7a, rep7b, repUS43, repUS5 | rep16, repUS5, rep22, rep7a, rep7b, repUS43 |
| Pathogenicity (%) | 97.9 | 97.7 | 97.7 |
| Spa-type | t011 | t011 | t011 |
| MLST | ST398 | ST398 | ST398 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leão, C.; Clemente, L.; Cara d’Anjo, M.; Albuquerque, T.; Amaro, A. Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal. Antibiotics 2022, 11, 1439. https://doi.org/10.3390/antibiotics11101439
Leão C, Clemente L, Cara d’Anjo M, Albuquerque T, Amaro A. Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal. Antibiotics. 2022; 11(10):1439. https://doi.org/10.3390/antibiotics11101439
Chicago/Turabian StyleLeão, Célia, Lurdes Clemente, Maria Cara d’Anjo, Teresa Albuquerque, and Ana Amaro. 2022. "Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal" Antibiotics 11, no. 10: 1439. https://doi.org/10.3390/antibiotics11101439
APA StyleLeão, C., Clemente, L., Cara d’Anjo, M., Albuquerque, T., & Amaro, A. (2022). Emergence of Cfr-Mediated Linezolid Resistance among Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) from Healthy Pigs in Portugal. Antibiotics, 11(10), 1439. https://doi.org/10.3390/antibiotics11101439