
Antibiotic Resistance in Campylobacter: A Systematic Review of South American Isolates
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
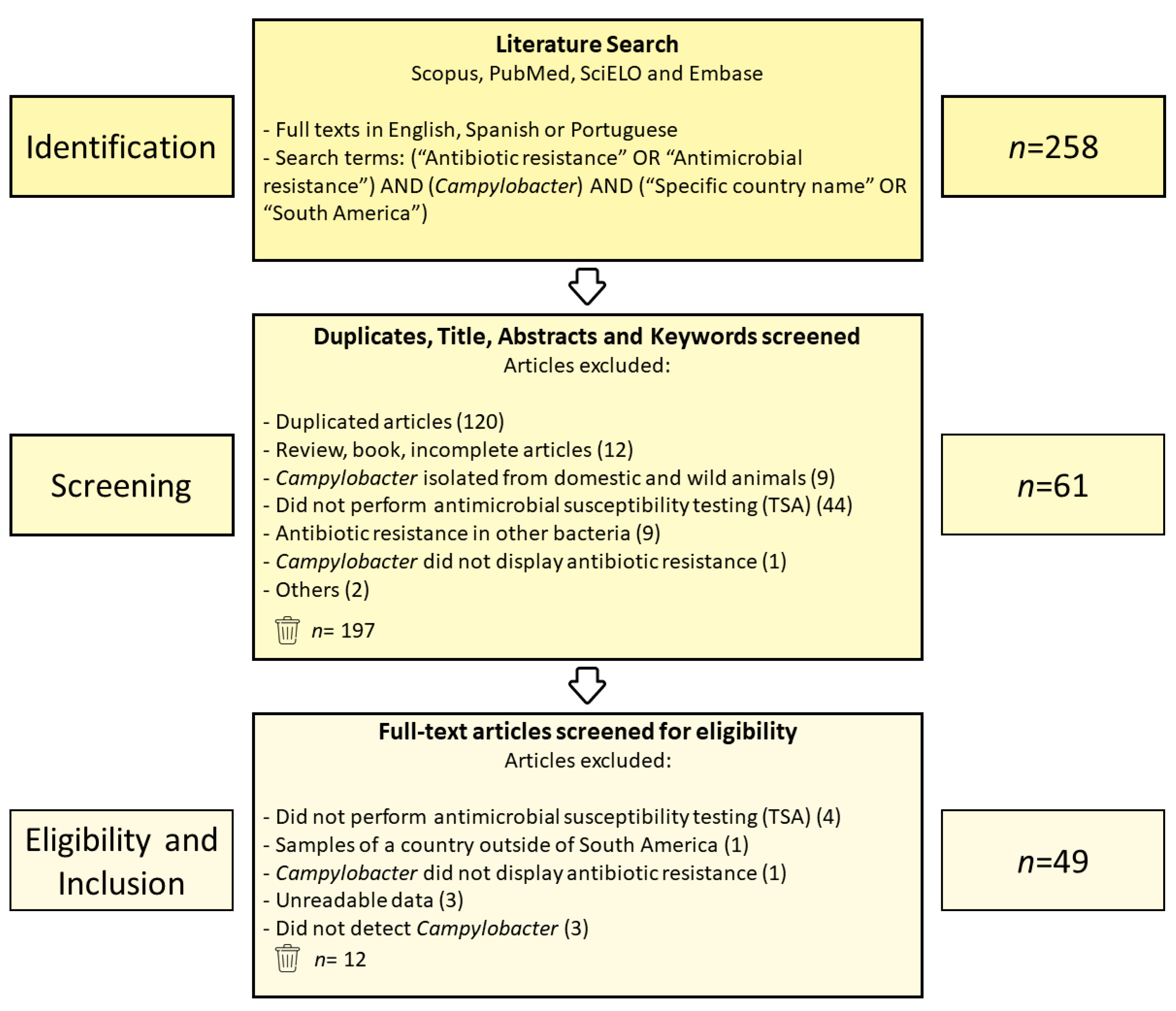
2.1. The Systematic Review Criteria
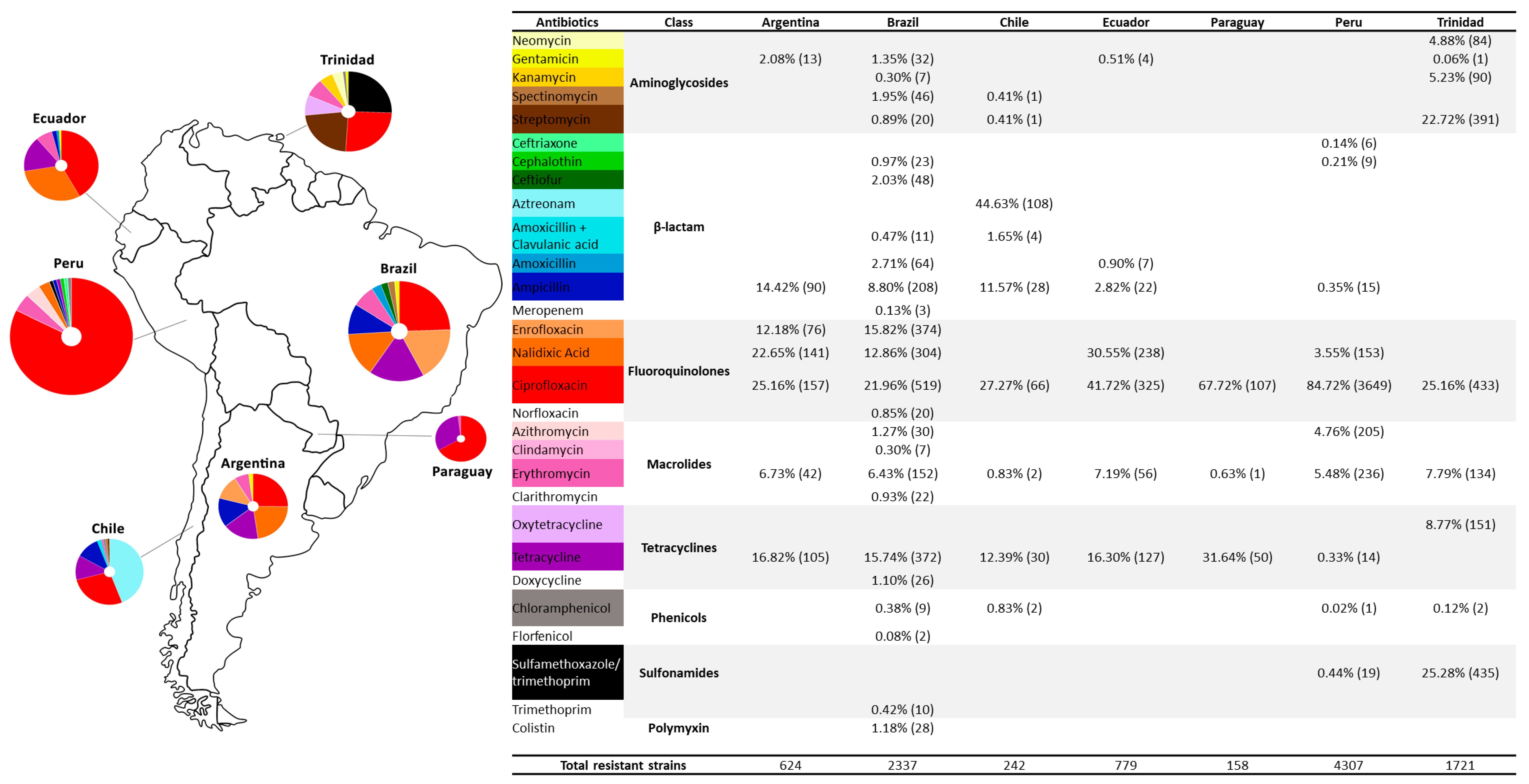
2.2. Frequency of Antibiotic-Resistant Campylobacter by South American Countries
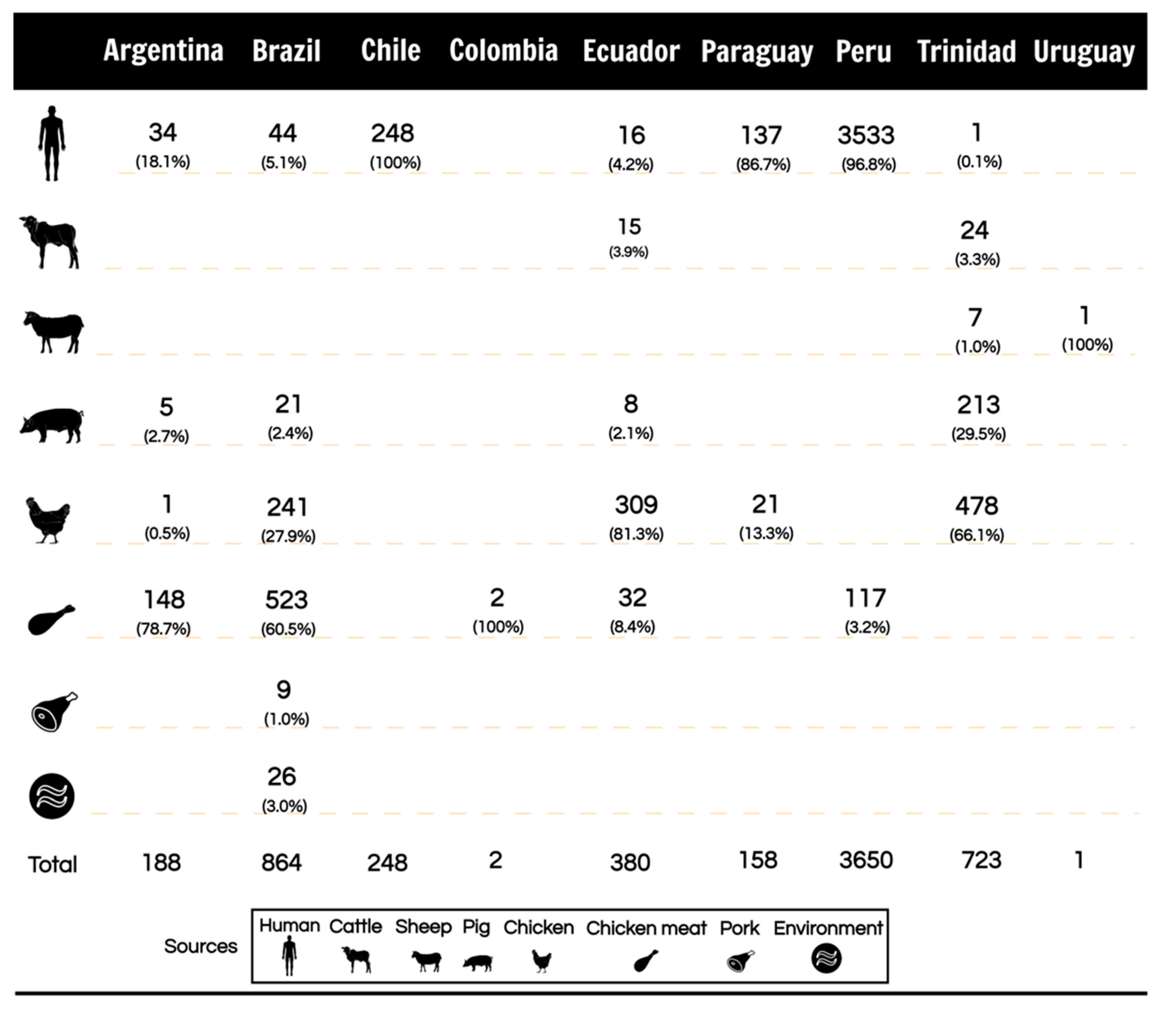
2.3. The Frequency of Campylobacter Species among Isolates Recovered from South American Eligible Studies
3. Discussion
4. Materials and Methods
4.1. Search Criteria
4.1.1. Inclusion Criteria
- Studies must address the detection of antimicrobial resistance in Campylobacter spp.
- Research must be conducted in South American countries.
- Campylobacter isolates must be derived from humans, food-producing animals, or are divergent.
- The study must report the total number of samples analyzed and the number of Campylobacter isolated from them.
- Confirmatory testing for Campylobacter should be addressed with biochemical and/or PCR/sequencing tests.
4.1.2. Exclusion Criteria
- Incomplete books, reviews, and articles.
- Studies written in a language other than English, Spanish, or Portuguese.
- Studies in which Campylobacter was not detected or was detected in sources other than humans, food-producing animals, or food of animal origin.
- Studies that did not perform antibiotic susceptibility testing or showed 100% antimicrobial sensibility.
4.2. Focus Questions
4.3. Assessment of the Risk of Bias
4.4. Frequency Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Igwaran, A.; Okoh, A.I. Human Campylobacteriosis: A Public Health Concern of Global Importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Wang, J.; Butaye, P.; Kelly, P.; Li, M.; Yang, F.; Gong, J.; Yassin, A.K.; Guo, W.; et al. Newly Identified Colistin Resistance Genes, Mcr-4 and Mcr-5, from Upper and Lower Alimentary Tract of Pigs and Poultry in China. PLoS ONE 2018, 13, e0193957. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Iraola, G. Pathogenomics of Emerging Campylobacter Species. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef]
- Tang, M.; Zhou, Q.; Zhang, X.; Zhou, S.; Zhang, J.; Tang, X.; Lu, J.; Gao, Y. Antibiotic Resistance Profiles and Molecular Mechanisms of Campylobacter from Chicken and Pig in China. Front. Microbiol. 2020, 11, 592496. [Google Scholar] [CrossRef]
- Man, S.M. The Clinical Importance of Emerging Campylobacter Species. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 669–685. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter Virulence and Survival Factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- Indikova, I.; Humphrey, T.J.; Hilbert, F. Survival with a Helping Hand: Campylobacter and Microbiota. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, H.L.; Dalager-Pedersen, M.; Nielsen, H. Risk of Inflammatory Bowel Disease after Campylobacter jejuni and Campylobacter concisus Infection: A Population-Based Cohort Study. Scand. J. Gastroenterol. 2019, 54, 265–272. [Google Scholar] [CrossRef]
- Leonhard, S.E.; Mandarakas, M.R.; Gondim, F.A.A.; Bateman, K.; Ferreira, M.L.B.; Cornblath, D.R.; van Doorn, P.A.; Dourado, M.E.; Hughes, R.A.C.; Islam, B.; et al. Diagnosis and Management of Guillain–Barré Syndrome in Ten Steps. Nat. Rev. Neurol. 2019, 15, 671–683. [Google Scholar] [CrossRef]
- Rodríguez, Y.; Rojas, M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Monsalve, D.M.; Gershwin, M.E.; Anaya, J.-M. Guillain–Barré Syndrome, Transverse Myelitis and Infectious Diseases. Cell. Mol. Immunol. 2018, 15, 547–562. [Google Scholar] [CrossRef] [Green Version]
- Sukted, N.; Tuitemwong, P.; Tuitemwong, K.; Poonlapdecha, W.; Erickson, L.E. Inactivation of Campylobacter during Immersion Chilling of Chicken Carcasses. J. Food Eng 2017, 202, 25–33. [Google Scholar] [CrossRef]
- COMMISSION REGULATION (EU) 2017/ 1495—of 23 August 2017—Amending Regulation (EC) No 2073/2005 as Regards Campylobacter in Broiler Carcases. Available online: https://www.legislation.gov.uk/eur/2017/1495/data.pdf (accessed on 10 January 2023).
- Adler-Mosca, H.; Lüthy-Hottenstein, J.; Martinetti Lucchini, G.; Burnens, A.; Altwegg, M. Development of Resistance to Quinolones in Five Patients with Campylobacteriosis Treated with Norfloxacin or Ciprofloxacin. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 953–957. [Google Scholar] [CrossRef]
- Bolinger, H.; Kathariou, S. The Current State of Macrolide Resistance in Campylobacter Spp.: Trends and Impacts of Resistance Mechanisms. Appl. Environ. Microbiol. 2017, 83, e00416-17. [Google Scholar] [CrossRef] [Green Version]
- Ata, Z.; Dinc, G.; Yibar, A.; Müştak, H.K.; Sahan, O. Extended Spectrum Beta-Lactamase Activity and Multidrug Resistance of Salmonella Serovars Isolated from Chicken Carcasses from Different Regions of Turkey. Vet. Fakültesi Derg. 2015, 62, 119–123. [Google Scholar] [CrossRef]
- Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; Ouellette, M. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Wieczorek, K.; Osek, J. Antimicrobial Resistance Mechanisms among Campylobacter. BioMed Res. Int. 2013, 2013, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.E.; Barton, M.D.; Blair, I.S.; Corcoran, D.; Dooley, J.S.G.; Fanning, S.; Kempf, I.; Lastovica, A.J.; Lowery, C.J.; Matsuda, M.; et al. The Epidemiology of Antibiotic Resistance in Campylobacter. Microbes Infect. 2006, 8, 1955–1966. [Google Scholar] [CrossRef]
- Vlieghe, E.R.; Jacobs, J.A.; Van Esbroeck, M.; Koole, O.; Van Gompel, A. Trends of Norfloxacin and Erythromycin Resistance of Campylobacter jejuni/Campylobacter coli Isolates Recovered from International Travelers, 1994 to 2006. J. Travel Med. 2008, 15, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Bernal, J.F.; Donado-Godoy, P.; Valencia, M.F.; León, M.; Gómez, Y.; Rodríguez, F.; Agarwala, R.; Landsman, D.; Mariño-Ramírez, L. Whole-Genome Sequences of Two Campylobacter coli Isolates from the Antimicrobial Resistance Monitoring Program in Colombia. Genome Announc. 2016, 4, e00131-16. [Google Scholar] [CrossRef] [Green Version]
- Pantozzi, F.L.; Moredo, F.A.; Vigo, G.B.; Giacoboni, G.I. Resistencia a los antimicrobianos en bacterias indicadoras y zoonóticas aisladas de animales domésticos en Argentina. Rev. Argent. de Microbiol. 2010, 42, 49–52. [Google Scholar]
- Tamborini, A.L.; Casabona, L.M.; Viñas, M.R.; Asato, V.; Hoffer, A.; Farace, M.I.; Lucero, M.C.; Corso, A.; Pichel, M. Campylobacter spp.: Prevalencia y caracterización feno-genotípica de aislamientos de pacientes con diarrea y de sus mascotas en la provincia de La Pampa, Argentina. Rev. Argent. de Microbiol. 2012, 44, 266–271. [Google Scholar]
- Zbrun, M.V.; Olivero, C.; Romero-Scharpen, A.; Rossler, E.; Soto, L.P.; Astesana, D.M.; Blajman, J.E.; Berisvil, A.; Signorini, M.L.; Frizzo, L.S. Antimicrobial Resistance in Thermotolerant Campylobacter Isolated from Different Stages of the Poultry Meat Supply Chain in Argentina. Food Control 2015, 57, 136–141. [Google Scholar] [CrossRef]
- Biasi, R.S.; Freitas de Macedo, R.E.; Scaranello Malaquias, M.A.; Franchin, P.R. Prevalence, Strain Identification and Antimicrobial Resistance of Campylobacter spp. Isolated from Slaughtered Pig Carcasses in Brazil. Food Control 2011, 22, 702–707. [Google Scholar] [CrossRef]
- de Moura, H.M.; Silva, P.R.; da Silva, P.H.C.; Souza, N.R.; Racanicci, A.M.C.; Santana, Â.P. Antimicrobial Resistance of Campylobacter jejuni Isolated from Chicken Carcasses in the Federal District, Brazil. J. Food Prot. 2013, 76, 691–693. [Google Scholar] [CrossRef]
- Dias, T.S.; Machado, L.S.; Vignoli, J.A.; Cunha, N.C.; Nascimento, E.R.; Pereira, V.L.A.; Aquino, M.H.C. Phenotypic and Molecular Characterization of Erythromycin Resistance in Campylobacter jejuni and Campylobacter coli Strains Isolated from Swine and Broiler Chickens. Pesq. Vet. Bras. 2020, 40, 598–603. [Google Scholar] [CrossRef]
- Dias, T.S.; Nascimento, R.J.; Machado, L.S.; Abreu, D.L.C.; do Nascimento, E.R.; Pereira, V.L.A.; de Aquino, M.H.C. Comparison of Antimicrobial Resistance in Thermophilic Campylobacter Strains Isolated from Conventional Production and Backyard Poultry Flocks. Br. Poult. Sci. 2021, 62, 188–192. [Google Scholar] [CrossRef]
- Ferro, I.D.; Benetti, T.M.; Oliveira, T.C.R.M.; Abrahão, W.M.; Farah, S.M.S.S.; Luciano, F.B.; Macedo, R.E.F. Evaluation of Antimicrobial Resistance of Campylobacter spp. Isolated from Broiler Carcasses. Br. Poult. Sci. 2015, 56, 66–71. [Google Scholar] [CrossRef]
- Fonseca, B.B.; Rossi, D.A.; Maia, C.A.; Nalevaiko, P.C.; Melo, R.T.; Cuccato, L.P.; Beletti, M.E. Characterization of the Virulence, Growth Temperature and Antibiotic Resistance of the Campylobacter jejuni IAL 2383 Strain Isolated from Humans. Braz. J. Microbiol. 2014, 45, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Frazão, M.R.; Cao, G.; Medeiros, M.I.C.; da Silva Duque, S.; Allard, M.W.; Falcão, J.P. Antimicrobial Resistance Profiles and Phylogenetic Analysis of Campylobacter jejuni Strains Isolated in Brazil by Whole Genome Sequencing. Microb. Drug Resist. 2021, 27, 660–669. [Google Scholar] [CrossRef]
- Gomes, C.N.; Frazão, M.R.; Passaglia, J.; Duque, S.S.; Medeiros, M.I.C.; Falcão, J.P. Molecular Epidemiology and Resistance Profile of Campylobacter jejuni and Campylobacter coli Strains Isolated from Different Sources in Brazil. Microb. Drug Resist. 2020, 26, 1516–1525. [Google Scholar] [CrossRef]
- Hungaro, H.M.; Mendonça, R.C.S.; Rosa, V.O.; Badaró, A.C.L.; Moreira, M.A.S.; Chaves, J.B.P. Low Contamination of Campylobacter spp. on Chicken Carcasses in Minas Gerais State, Brazil: Molecular Characterization and Antimicrobial Resistance. Food Control 2015, 51, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Hong, J.; Bae, W.; Koo, H.C.; Kim, S.H.; Park, Y.H. Prevalence, Antibiograms, and Transferable Tet(O) Plasmid of Campylobacter jejuni and Campylobacter coli Isolated from Raw Chicken, Pork, and Human Clinical Cases in Korea. J. Food Prot. 2010, 73, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Kleinubing, N.R.; Ramires, T.; de Fátima Rauber Würfel, S.; Haubert, L.; Scheik, L.K.; Kremer, F.S.; Lopes, G.V.; da Silva, W.P. Antimicrobial Resistance Genes and Plasmids in Campylobacter Jejuni from Broiler Production Chain in Southern Brazil. LWT 2021, 144, 111202. [Google Scholar] [CrossRef]
- Lopes, G.V.; Landgraf, M.; Destro, M.T. Occurrence of Campylobacter in Raw Chicken and Beef from Retail Outlets in São Paulo, Brazil. J. Food Saf. 2018, 38, e12442. [Google Scholar] [CrossRef]
- Melo, R.T.; Grazziotin, A.L.; Júnior, E.C.V.; Prado, R.R.; Mendonça, E.P.; Monteiro, G.P.; Peres, P.A.B.M.; Rossi, D.A. Evolution of Campylobacter jejuni of Poultry Origin in Brazil. Food Microbiol. 2019, 82, 489–496. [Google Scholar] [CrossRef]
- Nascimento, R.J.; Frasão, B.S.; Dias, T.S.; Nascimento, E.R.; Tavares, L.S.B.; Almeida, V.L.; Aquino, M.H.C. Detection of Efflux Pump CmeABC in Enrofloxacin Resistant Campylobacter spp. Strains Isolated from Broiler Chickens (Gallus Gallus Domesticus) in the State of Rio de Janeiro, Brazil. Pesq. Vet. Bras. 2019, 39, 728–733. [Google Scholar] [CrossRef]
- Paravisi, M.; Laviniki, V.; Bassani, J.; Kunert Filho, H.; Carvalho, D.; Wilsmann, D.; Borges, K.; Furian, T.; Salle, C.; Moraes, H.; et al. Antimicrobial Resistance in Campylobacter jejuni Isolated from Brazilian Poultry Slaughterhouses. Braz. J. Poult. Sci. 2020, 22, eRBCA-2020-1262. [Google Scholar] [CrossRef]
- Ramires, T.; de Oliveira, M.G.; Kleinubing, N.R.; de Fátima Rauber Würfel, S.; Mata, M.M.; Iglesias, M.A.; Lopes, G.V.; Dellagostin, O.A.; da Silva, W.P. Genetic Diversity, Antimicrobial Resistance, and Virulence Genes of Thermophilic Campylobacter Isolated from Broiler Production Chain. Braz. J. Microbiol. 2020, 51, 2021–2032. [Google Scholar] [CrossRef]
- Rossi, D.A.; Dumont, C.F.; de Souza Santos, A.C.; de Lourdes Vaz, M.E.; Prado, R.R.; Monteiro, G.P.; da Silva Melo, C.B.; Stamoulis, V.J.; dos Santos, J.P.; de Melo, R.T. Antibiotic Resistance in the Alternative Lifestyles of Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2021, 11, 535757. [Google Scholar] [CrossRef]
- Sierra-Arguello, Y.M.; Morgan, R.B.; Perdoncini, G.; Lima, L.M.; Gomes, M.J.P.; Nascimento, V.P. do Resistance to β-Lactam and Tetracycline in Campylobacter spp. Isolated from Broiler Slaughterhouses in Southern Brazil. Pesq. Vet. Bras. 2015, 35, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Arguello, Y.M.; Perdoncini, G.; Morgan, R.B.; Salle, C.T.P.; Moraes, H.L.S.; Gomes, M.J.P.; do Nascimento, V.P. Fluoroquinolone and Macrolide Resistance in Campylobacter jejuni Isolated from Broiler Slaughterhouses in Southern Brazil. Avian Pathol. 2016, 45, 66–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz, C.S.L.; Voss-Rech, D.; Lopes, L.S.; Silva, V.S. Applied Research Note: Pulsed-Field Gel Electrophoresis and Antimicrobial Resistance Profiles of Campylobacter jejuni Isolated from Brazilian Broiler Farms. J. Appl. Poult. Res. 2021, 30, 100168. [Google Scholar] [CrossRef]
- de Fátima Rauber Würfel, S.; Jorge, S.; de Oliveira, N.R.; Kremer, F.S.; Sanchez, C.D.; Campos, V.F.; da Silva Pinto, L.; da Silva, W.P.; Dellagostin, O.A. Campylobacter jejuni Isolated from Poultry Meat in Brazil: In Silico Analysis and Genomic Features of Two Strains with Different Phenotypes of Antimicrobial Susceptibility. Mol. Biol. Rep. 2020, 47, 671–681. [Google Scholar] [CrossRef] [PubMed]
- de Fátima Rauber Würfel, S.; da Fontoura Prates, D.; Kleinubing, N.R.; Vecchia, J.D.; Vaniel, C.; Haubert, L.; Dellagostin, O.A.; da Silva, W.P. Comprehensive Characterization Reveals Antimicrobial-Resistant and Potentially Virulent Campylobacter Isolates from Poultry Meat Products in Southern Brazil. LWT 2021, 149, 111831. [Google Scholar] [CrossRef]
- Bravo, V.; Katz, A.; Porte, L.; Weitzel, T.; Varela, C.; Gonzalez-Escalona, N.; Blondel, C.J. Genomic Analysis of the Diversity, Antimicrobial Resistance and Virulence Potential of Clinical Campylobacter jejuni and Campylobacter coli Strains from Chile. PLoS Negl. Trop. Dis. 2021, 15, e0009207. [Google Scholar] [CrossRef]
- Collado, L.; Muñoz, N.; Porte, L.; Ochoa, S.; Varela, C.; Muñoz, I. Genetic Diversity and Clonal Characteristics of Ciprofloxacin-Resistant Campylobacter jejuni Isolated from Chilean Patients with Gastroenteritis. Infect. Genet. Evol. 2018, 58, 290–293. [Google Scholar] [CrossRef]
- Fernández, H.; Mansilla, M.; González, V. Antimicrobial Susceptibility of Campylobacter jejuni subsp. Jejuni Assessed by E-Test and Double Dilution Agar Method in Southern Chile. Mem. Inst. Oswaldo Cruz 2000, 95, 247–249. [Google Scholar] [CrossRef] [Green Version]
- García, C.P.; Valenzuela, S.N.; Rodríguez, L.M.V.; León, C.E.; Fernández, J.H. Susceptibilidad antimicrobiana de Campylobacter jejuni aislado de coprocultivos en Santiago de Chile. Rev. Chil. Infectol. 2009, 26. [Google Scholar] [CrossRef] [Green Version]
- Levican, A.; Flores, O.; Sanchez, S.; Bascuñan, M.G.; Lopez, R.; Ojeda, K.; Hernandez, E.; Salah, P. Faecal Shedding of Campylobacteria among Domestic and Wild Animals from an Urban Coastal Área. Austral J. Vet. Sci. 2019, 51, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, S.; Simaluiza, R.J.; Toledo, Z.; Fernández, H. Frequency and Antimicrobial Behaviour of Thermophilic Campylobacter Species Isolated from Ecuadorian Backyard Chickens. Arch. Med. Vet. 2016, 48, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Simaluiza, R.J.; Toledo, Z.; Ochoa, S.; Fernandez, H. The Prevalence and Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli in Chicken Livers Used for Human Consumption in Ecuador. J. Anim. Vet. Adv. 2015, 14, 6–9. [Google Scholar]
- Simaluiza, R.; Toledo, Z.; Fernández, H. Prevalencia y caracterización del perfil de susceptibilidad antimicrobiana de Campylobacter jejuni y Campylobacter coli en niños con diarrea de la ciudad de Loja, Ecuador. Rev. Chil. Infectol. 2018, 35, 213–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, Z.; Simaluiza, R.J.; Fernández, H. Occurrence and Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli Isolated from Domestic Animals from Southern Ecuador. Cienc. Rural 2018, 48. [Google Scholar] [CrossRef]
- Vinueza-Burgos, C.; Wautier, M.; Martiny, D.; Cisneros, M.; Van Damme, I.; De Zutter, L. Prevalence, Antimicrobial Resistance and Genetic Diversity of Campylobacter coli and Campylobacter jejuni in Ecuadorian Broilers at Slaughter Age. Poult. Sci. 2017, 96, 2366–2374. [Google Scholar] [CrossRef]
- Cardozo, L.; Castro, L.; Zarate, N.; Torres, C.; Stavis, S. Presence of Campylobacter spp. and Antimicrobial Resistance to Ciprofloxacin and Erythromycin in One Laying Hens Production Stablishment of THE Central Department, Paraguay. Compend. Cienc. Vet. 2017, 7, 7–11. [Google Scholar] [CrossRef]
- Anampa, D.; Benites, C.; Lázaro, C.; Espinoza, J.; Angulo, P.; Díaz, D.; Manchego, A.; Rojas, M. Detección del gen ermB asociado a la resistencia a macrólidos en cepas de Campylobacter aisladas de pollos comercializados en Lima, Perú. Rev. Panam. de Salud Pública 2020, 44, 1. [Google Scholar] [CrossRef]
- Espinoza, N.; Rojas, J.; Pollett, S.; Meza, R.; Patiño, L.; Leiva, M.; Camiña, M.; Bernal, M.; Reynolds, N.D.; Maves, R.; et al. Validation of the T86I Mutation in the GyrA Gene as a Highly Reliable Real Time PCR Target to Detect Fluoroquinolone-Resistant Campylobacter jejuni. BMC Infect. Dis. 2020, 20, 518. [Google Scholar] [CrossRef]
- Jennings, M.C.; Tilley, D.H.; Ballard, S.-B.; Villanueva, M.; Costa, F.M.; Lopez, M.; Steinberg, H.E.; Giannina Luna, C.; Meza, R.; Silva, M.E.; et al. Case–Case Analysis Using 7 Years of Travelers’ Diarrhea Surveillance Data: Preventive and Travel Medicine Applications in Cusco, Peru. Am. J. Trop. Med. Hyg. 2017, 16–0633. [Google Scholar] [CrossRef] [Green Version]
- Pollett, S.; Rocha, C.; Zerpa, R.; Patiño, L.; Valencia, A.; Camiña, M.; Guevara, J.; Lopez, M.; Chuquiray, N.; Salazar-Lindo, E.; et al. Campylobacterantimicrobial Resistance in Peru: A Ten-Year Observational Study. BMC Infect. Dis. 2012, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Moya-Salazar, J.; Terán-Vásquez, A.; Salazar-Hernández, R. Alta Resistencia Antimicrobiana a Fluoroquinolonas por Campylobacter en Pacientes Pediátricos de un Hospital Peruano. Rev. Peru Med. Exp. Salud Public 2018, 35, 155–156 . [Google Scholar]
- Taitt, C.R.; Leski, T.A.; Prouty, M.G.; Ford, G.W.; Heang, V.; House, B.L.; Levin, S.Y.; Curry, J.A.; Mansour, A.; Mohammady, H.E.; et al. Tracking Antimicrobial Resistance Determinants in Diarrheal Pathogens: A Cross-Institutional Pilot Study. Int. J. Mol. Sci. 2020, 21, 5928. [Google Scholar] [CrossRef] [PubMed]
- Adesiyun, A.A.; Kaminjolo, J.S.; Loregnard, R.; Kitson-Piggott, W. Campylobacter Infections in Calves, Piglets, Lambs and Kids in Trinidad. Br. Vet. J. 1992, 148, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Devi, A.; Mahony, T.J.; Wilkinson, J.M.; Vanniasinkam, T. Antimicrobial Susceptibility of Clinical Isolates of Campylobacter jejuni from New South Wales, Australia. J. Glob. Antimicrob. Resist. 2019, 16, 76–80. [Google Scholar] [CrossRef]
- Schiaffino, F.; Colston, J.M.; Paredes-Olortegui, M.; François, R.; Pisanic, N.; Burga, R.; Peñataro-Yori, P.; Kosek, M.N. Antibiotic Resistance of Campylobacter Species in a Pediatric Cohort Study. Antimicrob. Agents Chemother. 2019, 63, e01911-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signorini, M.L.; Rossler, E.; Díaz David, D.C.; Olivero, C.R.; Romero-Scharpen, A.; Soto, L.P.; Astesana, D.M.; Berisvil, A.P.; Zimmermann, J.A.; Fusari, M.L.; et al. Antimicrobial Resistance of Thermotolerant Campylobacter Species Isolated from Humans, Food-Producing Animals, and Products of Animal Origin: A Worldwide Meta-Analysis. Microb. Drug Resist. 2018, 24, 1174–1190. [Google Scholar] [CrossRef] [PubMed]
- Sproston, E.L.; Wimalarathna, H.M.L.; Sheppard, S.K. Trends in Fluoroquinolone Resistance in Campylobacter. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef]
- Changkwanyeun, R.; Usui, M.; Kongsoi, S.; Yokoyama, K.; Kim, H.; Suthienkul, O.; Changkaew, K.; Nakajima, C.; Tamura, Y.; Suzuki, Y. Characterization of Campylobacter jejuni DNA Gyrase as the Target of Quinolones. J. Infect. Chemother. 2015, 21, 604–609. [Google Scholar] [CrossRef]
- Han, J.; Wang, Y.; Sahin, O.; Shen, Z.; Guo, B.; Shen, J.; Zhang, Q. A Fluoroquinolone Resistance Associated Mutation in GyrA Affects DNA Supercoiling in Campylobacter jejuni. Front. Cell. Inf. Microbio. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Aksomaitiene, J.; Ramonaite, S.; Olsen, J.E.; Malakauskas, M. Prevalence of Genetic Determinants and Phenotypic Resistance to Ciprofloxacin in Campylobacter jejuni from Lithuania. Front. Microbiol. 2018, 9, 203. [Google Scholar] [CrossRef]
- Engberg, J. Quinolone and Macrolide Resistance in Campylobacter jejuni and C. coli: Resistance Mechanisms and Trends in Human Isolates. Emerg. Infect. Dis. 2001, 7, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connell, S.R.; Tracz, D.M.; Nierhaus, K.H.; Taylor, D.E. Ribosomal Protection Proteins and Their Mechanism of Tetracycline Resistance. Antimicrob. Agents Chemother. 2003, 47, 3675–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Salvatore, D.; Sakhria, S.; Catelli, E.; Lupini, C.; Abbassi, M.S.; Bessoussa, G.; Ben Yahia, S.; Ben Chehida, N. High Frequency and Diversity of Tetracycline Resistance Genes in the Microbiota of Broiler Chickens in Tunisia. Animals 2021, 11, 377. [Google Scholar] [CrossRef]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6, 6.4.14. [Google Scholar] [CrossRef] [Green Version]
- Vilela, F.P.; Gomes, C.N.; Passaglia, J.; Rodrigues, D.P.; Costa, R.G.; Tiba Casas, M.R.; Fernandes, S.A.; Falcão, J.P.; Campioni, F. Genotypic Resistance to Quinolone and Tetracycline in Salmonella Dublin Strains Isolated from Humans and Animals in Brazil. Microb. Drug Resist. 2019, 25, 143–151. [Google Scholar] [CrossRef]
- García-Fernández, A.; Dionisi, A.M.; Arena, S.; Iglesias-Torrens, Y.; Carattoli, A.; Luzzi, I. Human Campylobacteriosis in Italy: Emergence of Multi-Drug Resistance to Ciprofloxacin, Tetracycline, and Erythromycin. Front. Microbiol. 2018, 9, 1906. [Google Scholar] [CrossRef] [Green Version]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Kielsznia, A.; Korzekwa, K.; Tobiasz, A.; Korzeniowska-Kowal, A.; Wieliczko, A. High Prevalence of Resistance to Fluoroquinolones and Tetracycline Campylobacter Spp. Isolated from Poultry in Poland. Microb. Drug Resist. 2018, 24, 314–322. [Google Scholar] [CrossRef]
- Sougakoff, W.; Papadopoulou, B.; Nordmann, P.; Courvalin, P. Nucleotide Sequence and Distribution of Gene TetO Encoding Tetracycline Resistance in Campylobacter coli. FEMS Microbiol. Lett. 1987, 44, 153–159. [Google Scholar] [CrossRef]
- Sagara, H.; Mochizuki, A.; Okamura, N.; Nakaya, R. Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli with Special Reference to Plasmid Profiles of Japanese Clinical Isolates. Antimicrob. Agents Chemother. 1987, 31, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Tenover, F.C.; Bronsdon, M.A.; Gordon, K.P.; Plorde, J.J. Isolation of Plasmids Encoding Tetracycline Resistance from Campylobacter Jejuni Strains Isolated from Simians. Antimicrob. Agents Chemother. 1983, 23, 320–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hormeño, L.; Campos, M.J.; Vadillo, S.; Quesada, A. Occurrence of Tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter. Microorganisms 2020, 8, 1710. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in Food and Feedingstuffs: from Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. J. Anal. Methods Chem. 2017, 2017, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Iovine, N.M. Resistance Mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.I.; Kaye, K.M. Beta-Lactam Antibiotics: Newer Formulations and Newer Agents. Infect. Dis. Clin. North Am. 2004, 18, 603–619. [Google Scholar] [CrossRef]
- Casagrande Proietti, P.; Guelfi, G.; Bellucci, S.; De Luca, S.; Di Gregorio, S.; Pieramati, C.; Franciosini, M.P. Beta-Lactam Resistance in Campylobacter coli and Campylobacter jejuni Chicken Isolates and the Association between BlaOXA-61 Gene Expression and the Action of β-Lactamase Inhibitors. Vet. Microbiol. 2020, 241, 108553. [Google Scholar] [CrossRef]
- Hadiyan, M.; Momtaz, H.; Shakerian, A. Prevalence, Antimicrobial Resistance, Virulence Gene Profile and Molecular Typing of Campylobacter Species Isolated from Poultry Meat Samples. Vet. Med. Sci. 2022, 8, 2482–2493. [Google Scholar] [CrossRef]
- Alfredson, D.A.; Korolik, V. Isolation and Expression of a Novel Molecular Class D β-Lactamase, OXA-61, from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2515–2518. [Google Scholar] [CrossRef] [Green Version]
- Lachance, N.; Gaudreau, C.; Lamothe, F.; Larivière, L.A. Role of the Beta-Lactamase of Campylobacter Jejuni in Resistance to Beta-Lactam Agents. Antimicrob. Agents Chemother. 1991, 35, 813–818. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Brown, S.; Gillespie, B.; Lin, J. A Single Nucleotide in the Promoter Region Modulates the Expression of the -Lactamase OXA-61 in Campylobacter jejuni. J. Antimicrob. Chemother. 2014, 69, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Sykes, R.B.; Bonner, D.P. Aztreonam: The First Monobactam. Am. J. Med. 1985, 78, 9. [Google Scholar] [CrossRef] [PubMed]
- Hellinger, W.C.; Brewer, N.S. Carbapenems and Monobactams: Imipenem, Meropenem, and Aztreonam. Mayo Clin. Proc. 1999, 74, 420–434. [Google Scholar] [CrossRef]
- Thomas, G.D. Pilot Study for the Development of a New Campylobacter Selective Medium at 37 °C Using Aztreonam. J. Clin. Pathol. 2005, 58, 413–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo, S.; Adesiyun, A.; Asgarali, Z.; Swanston, W. Antimicrobial Resistance of Campylobacter spp. Isolated from Broilers in Small Poultry Processing Operations in Trinidad. Food Control 2007, 18, 321–325. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Engberg, J. Antimicrobial Resistance of Thermophilic Campylobacter. Vet. Res. 2001, 32, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Lehtopolku, M.; Nakari, U.-M.; Kotilainen, P.; Huovinen, P.; Siitonen, A.; Hakanen, A.J. Antimicrobial Susceptibilities of Multidrug-Resistant Campylobacter jejuni and C. coli Strains: In Vitro Activities of 20 Antimicrobial Agents. Antimicrob. Agents Chemother. 2010, 54, 1232–1236. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Mukherjee, S.; Chen, Y.; Li, C.; Young, S.; Warren, M.; Abbott, J.; Friedman, S.; Kabera, C.; Karlsson, M.; et al. Novel Gentamicin Resistance Genes in Campylobacter Isolated from Humans and Retail Meats in the USA. J. Antimicrob. Chemother. 2015, 70, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Mukherjee, S.; Hoffmann, M.; Kotewicz, M.L.; Young, S.; Abbott, J.; Luo, Y.; Davidson, M.K.; Allard, M.; McDermott, P.; et al. Whole-Genome Sequencing of Gentamicin-Resistant Campylobacter coli Isolated from U.S. Retail Meats Reveals Novel Plasmid-Mediated Aminoglycoside Resistance Genes. Antimicrob. Agents Chemother. 2013, 57, 5398–5405. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.D.; Sanchez, S.; Zimmer, M.; Idris, U.; Berrang, M.E.; McDermott, P.F. Class 1 Integron-Associated Tobramycin-Gentamicin Resistance in Campylobacter jejuni Isolated from the Broiler Chicken House Environment. Antimicrob. Agents Chemother. 2002, 46, 3660–3664. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Wang, Y.; Zhang, Q.; Chen, X.; Shen, Z.; Deng, F.; Wu, C.; Shen, J. Identification of a Novel Genomic Island Conferring Resistance to Multiple Aminoglycoside Antibiotics in Campylobacter coli. Antimicrob. Agents Chemother. 2012, 56, 5332–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, M.; Frase, H.; Antunes, N.T.; Vakulenko, S.B. Novel Aminoglycoside 2″-Phosphotransferase Identified in a Gram-Negative Pathogen. Antimicrob. Agents Chemother. 2013, 57, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Adawy, H.; Hotzel, H.; Düpre, S.; Tomaso, H.; Neubauer, H.; Hafez, H.M. Determination of Antimicrobial Sensitivities of Campylobacter jejuni Isolated from Commercial Turkey Farms in Germany. Avian Dis. 2012, 56, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, M.; Salata, C.; Martini, M.; Montesissa, C.; Piccirillo, A. Antimicrobial Resistance of Campylobacter jejuni and Campylobacter coli from Poultry in Italy. Microb. Drug Resist. 2014, 20, 181–188. [Google Scholar] [CrossRef]
- Nayak, R.; Stewart, T.; Nawaz, M.; Cerniglia, C. In Vitro Antimicrobial Susceptibility, Genetic Diversity and Prevalence of UDP-Glucose 4-Epimerase (GalE) Gene in Campylobacter coli and Campylobacter jejuni from Turkey Production Facilities. Food Microbiol. 2006, 23, 379–392. [Google Scholar] [CrossRef]
- Karmali, M.A.; De Grandis, S.; Fleming, P.C. Antimicrobial Susceptibility of Campylobacter jejuni with Special Reference to Resistance Patterns of Canadian Isolates. Antimicrob. Agents Chemother. 1981, 19, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Gibreel, A.; Sköld, O. High-Level Resistance to Trimethoprim in Clinical Isolates of Campylobacter jejuni by Acquisition of Foreign Genes (Dfr1 and Dfr9) Expressing Drug-Insensitive Dihydrofolate Reductases. Antimicrob. Agents Chemother. 1998, 42, 3059–3064. [Google Scholar] [CrossRef] [Green Version]
- Dorsch, M.A.; Casaux, M.L.; Calleros, L.; Aráoz, V.; Caffarena, R.D.; Monesiglio, C.; Barcellos, M.; da Silva Silveira, C.; Perdomo, Y.; Banchero, G.; et al. Placentitis and Abortion Caused by a Multidrug Resistant Strain of Campylobacter fetus Subspecies Fetus in a Sheep in Uruguay. Rev. Argent. de Microbiol. 2022, 54, 25–30. [Google Scholar] [CrossRef]
- Sahin, O.; Yaeger, M.; Wu, Z.; Zhang, Q. Campylobacter—Associated Diseases in Animals. Annu. Rev. Anim. Biosci. 2017, 5, 21–42. [Google Scholar] [CrossRef]
- Wagenaar, J.A.; van Bergen, M.A.P.; Blaser, M.J.; Tauxe, R.V.; Newell, D.G.; van Putten, J.P.M. Campylobacter Fetus Infections in Humans: Exposure and Disease. Clin. Infect. Dis. 2014, 58, 1579–1586. [Google Scholar] [CrossRef] [Green Version]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.N.; Passaglia, J.; Vilela, F.P.; Pereira da Silva, F.M.H.S.; Duque, S.S.; Falcão, J.P. High Survival Rates of Campylobacter coli under Different Stress Conditions Suggest That More Rigorous Food Control Measures Might Be Needed in Brazil. Food Microbiol. 2018, 73, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Meredith, H.; Valdramidis, V.; Rotabakk, B.T.; Sivertsvik, M.; McDowell, D.; Bolton, D.J. Effect of Different Modified Atmospheric Packaging (MAP) Gaseous Combinations on Campylobacter and the Shelf-Life of Chilled Poultry Fillets. Food Microbiol. 2014, 44, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, N.; McKenna, A.; Richmond, A.; Ricke, S.C.; Callaway, T.; Stratakos, A.C.; Gundogdu, O.; Corcionivoschi, N. A Review of the Effect of Management Practices on Campylobacter Prevalence in Poultry Farms. Front. Microbiol. 2018, 9, 2002. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.C.; Targino, B.N.; Mendonça, R.S.; Sant’Ana, A.S.; Hungaro, H.M. Campylobacter: An Overview of Cases, Occurrence in Food, Contamination Sources, and Antimicrobial Resistance in Brazil. Food Rev. Int. 2018, 34, 364–389. [Google Scholar] [CrossRef]
- Frirdich, E.; Biboy, J.; Pryjma, M.; Lee, J.; Huynh, S.; Parker, C.T.; Girardin, S.E.; Vollmer, W.; Gaynor, E.C. The Campylobacter jejuni Helical to Coccoid Transition Involves Changes to Peptidoglycan and the Ability to Elicit an Immune Response. Mol. Microbiol. 2019, 112, 280–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmonts, M.-H.; Dufour-Gesbert, F.; Avrain, L.; Kempf, I. Antimicrobial Resistance in Campylobacter Strains Isolated from French Broilers before and after Antimicrobial Growth Promoter Bans. J. Antimicrob. Chemother. 2004, 54, 1025–1030. [Google Scholar] [CrossRef] [Green Version]
- Gomes, B.C.; de Melo Franco, B.D.G.; De Martinis, E.C.P. Microbiological Food Safety Issues in Brazil: Bacterial Pathogens. Foodborne Pathog. Dis. 2013, 10, 197–205. [Google Scholar] [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbio. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Cody, A.J.; Bray, J.E.; Jolley, K.A.; McCarthy, N.D.; Maiden, M.C.J. Core Genome Multilocus Sequence Typing Scheme for Stable, Comparative Analyses of Campylobacter jejuni and C. coli Human Disease Isolates. J. Clin. Microbiol. 2017, 55, 2086–2097. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.G.A.; Costantino, R.; Bianco, A.; Pileggi, C.; Pavia, M. Prevalence and Pattern of Antibiotic Resistance of Campylobacter spp. in Poultry Meat in Southern Italy. Food Control 2013, 32, 715–718. [Google Scholar] [CrossRef]
- Suzuki, H.; Yamamoto, S. Campylobacter Contamination in Retail Poultry Meats and By-Products in the World: A Literature Survey. J. Vet. Med. Sci. 2009, 71, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Country | Articles (n) | Sample Analyzed | Campylobacter Isolated (n) | Campylobacter Species (n) | Frequency (%) | Reference |
|---|---|---|---|---|---|---|
| Argentina | 3 | 50 | 11 | C. jejuni (8) | C. jejuni (75.6%) | [25] |
| C. coli (3) | C. coli (22.5%) | |||||
| 327 | 50 | C. jejuni (48) | [22] | |||
| C. coli (2) | ||||||
| 555 | 152 | C. jejuni (105) | [23] | |||
| C. coli (43) | ||||||
| Brazil | 22 | 259 | 9 | C. jejuni (5) | C. jejuni (82.9%) | [24] |
| C. coli (3) | C. coli (13.0%) | |||||
| 92 | 16 | C. jejuni (16) | C. lari (0.1%) | [25] | ||
| 50 | 34 | C. coli (14) | [26] | |||
| 70 | 70 | C. jejuni (69) | [27] | |||
| 24 | 24 | C. jejuni (22) | [28] | |||
| C. coli (1) | ||||||
| C. lari (1) | ||||||
| 1 | 1 | C. jejuni (1) | [29] | |||
| 67 | 67 | C. jejuni (67) | [30] | |||
| 42 | 42 | C. jejuni (14) | [31] | |||
| C. coli (25) | ||||||
| 95 | 20 | C. jejuni (18) | [32] | |||
| C. coli (2) | ||||||
| 78 | 46 | C. jejuni (39) | [33] | |||
| C. coli (7) | ||||||
| 173 | 28 | C. jejuni (28) | [34] | |||
| 120 | 18 | C. jejuni (5) | [35] | |||
| C. coli (13) | ||||||
| 1070 | 99 | C. jejuni (99) | [36] | |||
| 159 | 159 | C. jejuni (81) | [37] | |||
| C. coli (78) | ||||||
| 54 | 54 | C. jejuni (54) | [38] | |||
| 116 | 116 | C. jejuni (116) | [39] | |||
| 442 | 35 | C. jejuni (35) | [40] | |||
| 141 | 141 | C. jejuni (140) | [41] | |||
| C. coli (1) | ||||||
| 50 | 50 | C. jejuni (50) | [42] | |||
| 515 | 80 | C. jejuni (80) | [43] | |||
| 2 | 2 | C. jejuni (2) | [44] | |||
| 48 | 32 | C. jejuni (32) | [45] | |||
| Chile | 5 | 81 | 81 | C. jejuni (69) | C. jejuni (95.9%) | [46] |
| C. coli (12) | C. coli (4.1%) | |||||
| 50 | 50 | C. jejuni (50) | [47] | |||
| 108 | 108 | C. jejuni (108) | [48] | |||
| 73 | 73 | C. jejuni (73) | [49] | |||
| 350 | 28 | C. jejuni (26) | [50] | |||
| C. coli (2) | ||||||
| Colombia | 1 | 2 | 2 | C. coli (2) | C. coli (100%) | [20] |
| Ecuador | 5 | 120 | 50 | C. jejuni (39) | C. jejuni (77.1%) | [51] |
| C. coli (11) | C. coli (22.9%) | |||||
| 51 | 32 | C. jejuni (22) | [52] | |||
| C. coli (10) | ||||||
| 253 | 16 | C. jejuni (13) | [53] | |||
| C. coli (3) | ||||||
| 250 | 64 | C. jejuni (49) | [54] | |||
| C. coli (15) | ||||||
| 379 | 218 | C. jejuni (170) | [55] | |||
| C. coli (48) | ||||||
| Paraguay | 1 | 150 | 22 | C. jejuni (21) | C. jejuni (95.4%) | [56] |
| Peru | 6 | 120 | 117 | C. coli (117) | C. jejuni (80.4%) | [57] |
| 189 | 189 | C. jejuni (189) | C. coli (14.8%) | [58] | ||
| 230 | 19 | C. jejuni (16) | [59] | |||
| C. coli (3) | ||||||
| 4652 | 4652 | C. jejuni (3856) | [60] | |||
| C. coli (554) | ||||||
| 150 | 106 | C. jejuni (30) | [61] | |||
| C. coli (76) | ||||||
| 7 | 7 | C. jejuni (4) | [62] | |||
| C. coli (3) | ||||||
| Trinidad | 1 | 689 | 315 | C. jejuni (89) | C. jejuni (28.2%) | [63] |
| C. coli (226) | C. coli (71.7%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portes, A.B.; Panzenhagen, P.; Pereira dos Santos, A.M.; Junior, C.A.C. Antibiotic Resistance in Campylobacter: A Systematic Review of South American Isolates. Antibiotics 2023, 12, 548. https://doi.org/10.3390/antibiotics12030548
Portes AB, Panzenhagen P, Pereira dos Santos AM, Junior CAC. Antibiotic Resistance in Campylobacter: A Systematic Review of South American Isolates. Antibiotics. 2023; 12(3):548. https://doi.org/10.3390/antibiotics12030548
Chicago/Turabian StylePortes, Ana Beatriz, Pedro Panzenhagen, Anamaria Mota Pereira dos Santos, and Carlos Adam Conte Junior. 2023. "Antibiotic Resistance in Campylobacter: A Systematic Review of South American Isolates" Antibiotics 12, no. 3: 548. https://doi.org/10.3390/antibiotics12030548
APA StylePortes, A. B., Panzenhagen, P., Pereira dos Santos, A. M., & Junior, C. A. C. (2023). Antibiotic Resistance in Campylobacter: A Systematic Review of South American Isolates. Antibiotics, 12(3), 548. https://doi.org/10.3390/antibiotics12030548