New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori

 ,
,  ,
,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Solvents
2.2. Plant Material and Preparation of the Crude Extract
2.3. Fractionation Procedure
2.4. Determination of Total Polyphenols and Total Flavonoids
2.5. UHPLC-HRMS Analysis
2.6. Antimicrobial Disk Diffusion Test
2.7. In-Cell Urease Activity Test
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition of S. campanulata Fractions and Sub-Fractions
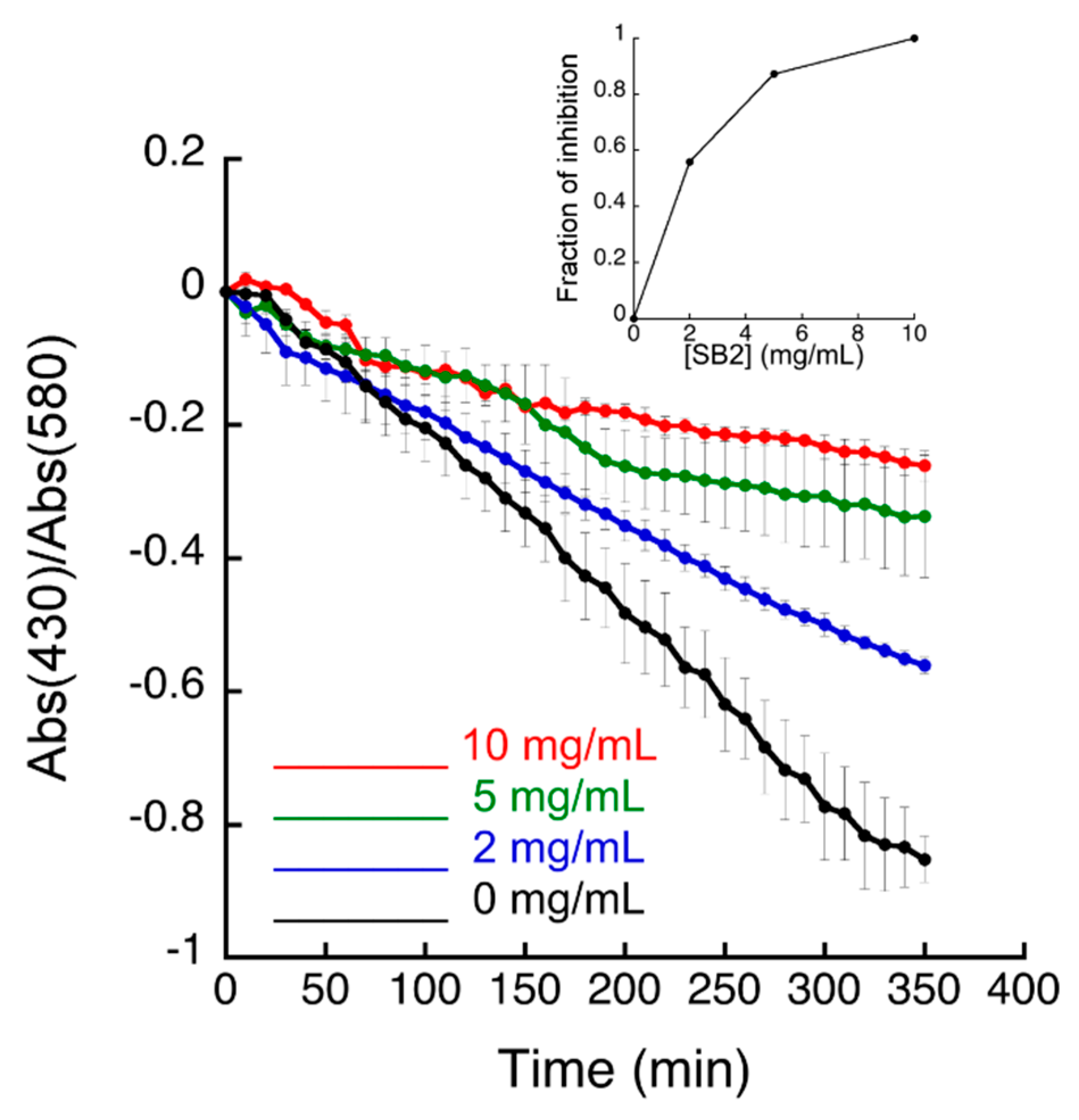
3.2. Inhibitory Activity of S. campanulata Fractions and Sub-Fractions against H. pylori
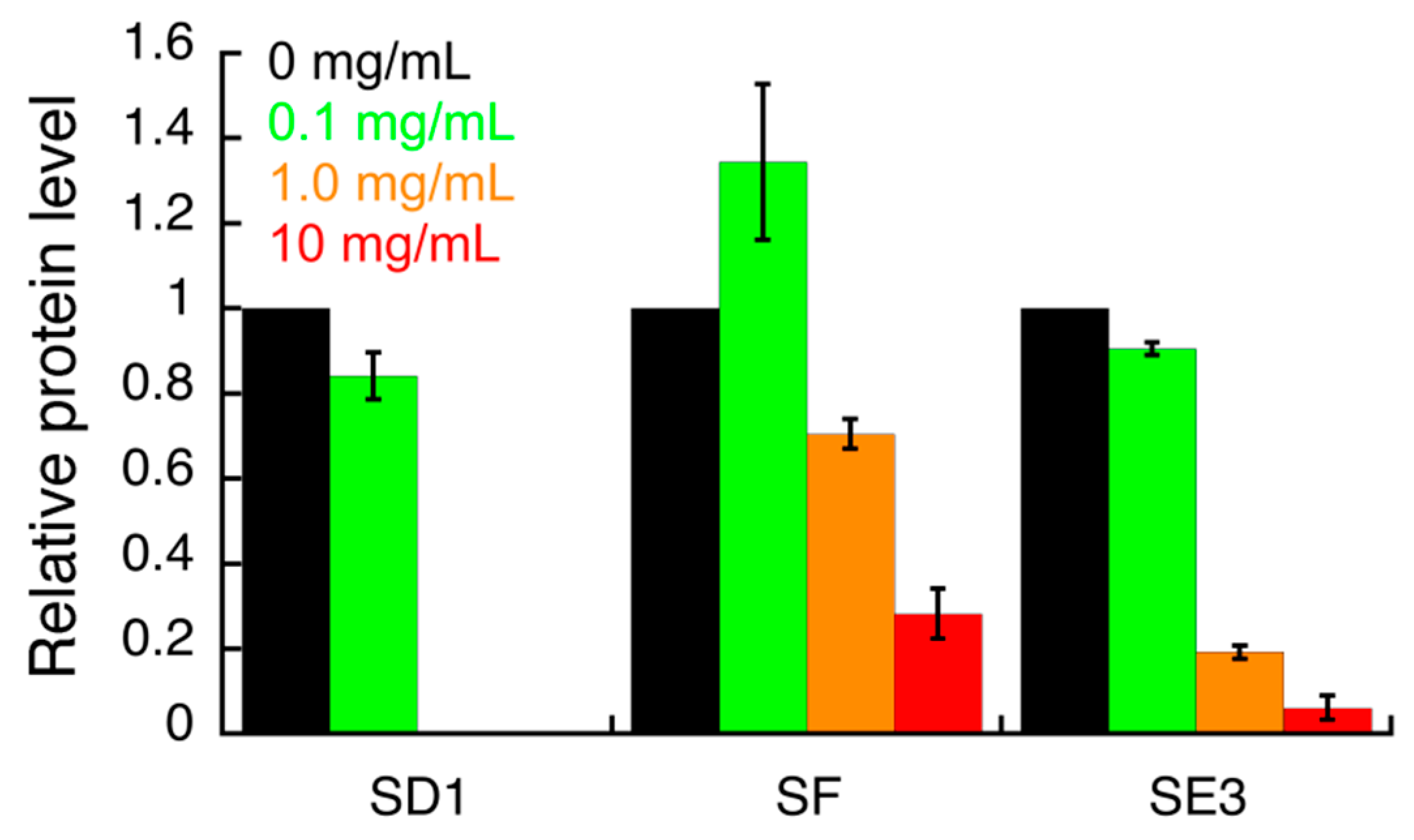
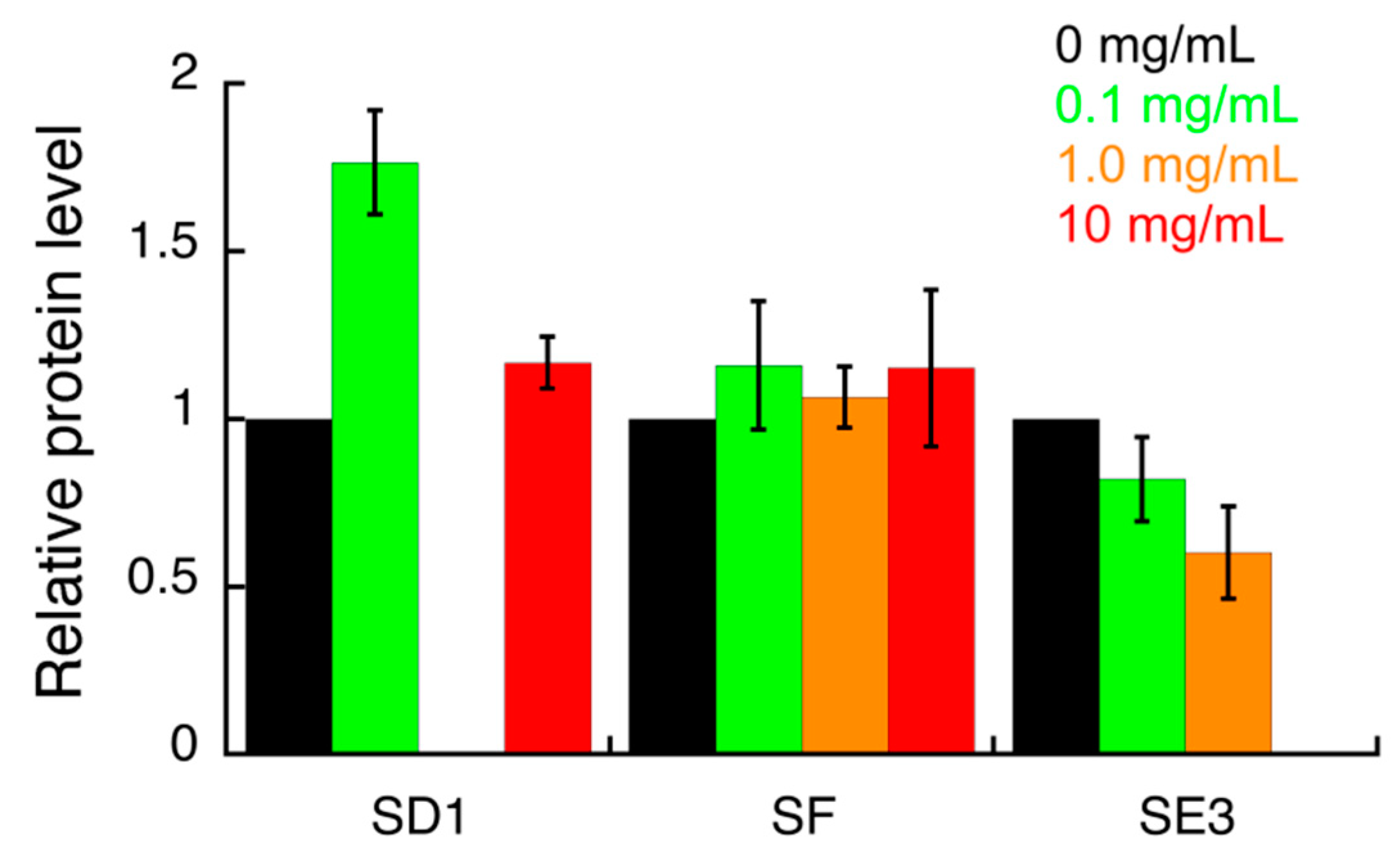
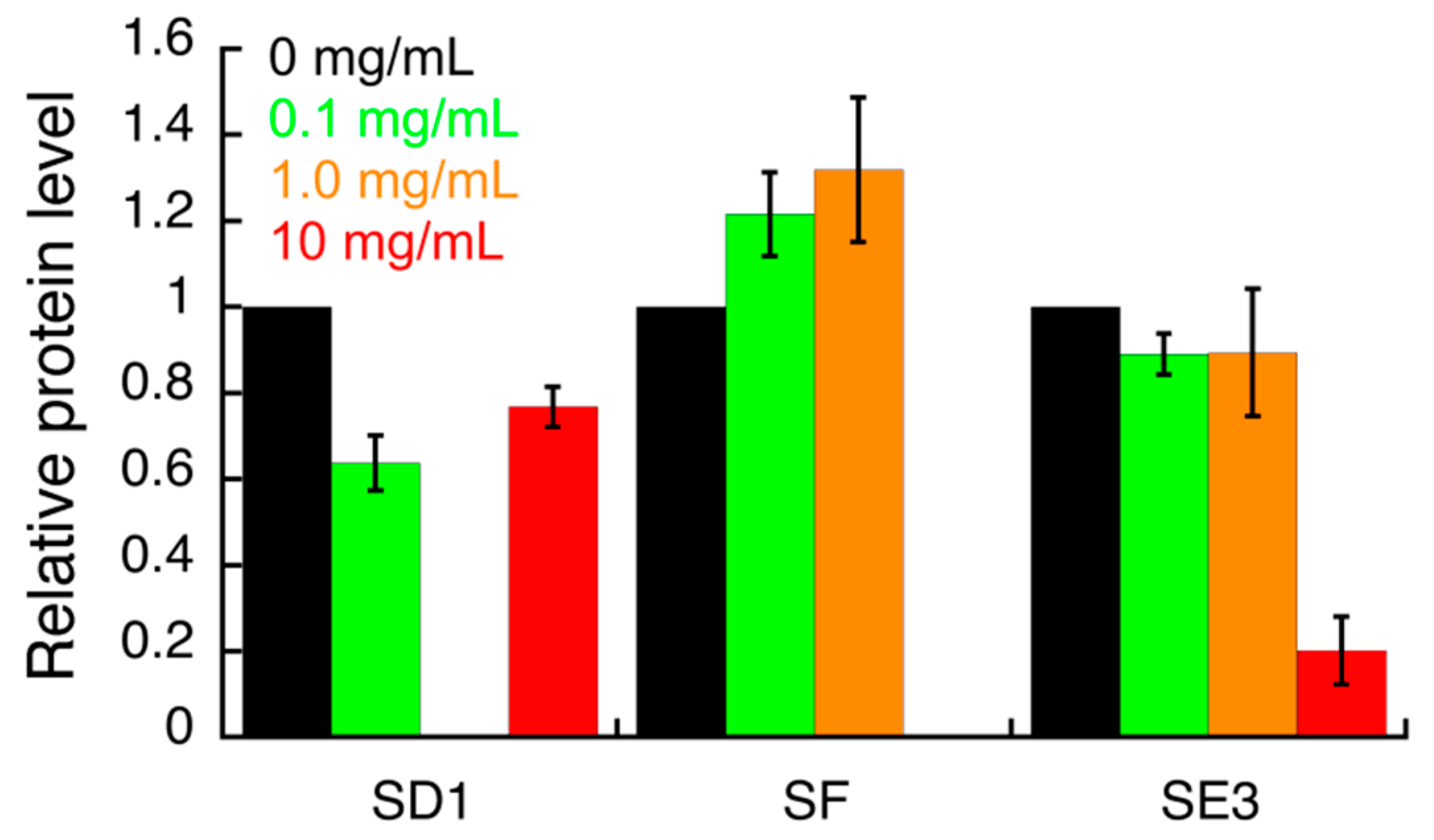
3.3. Effect of S. campanulata Fractions and Sub-Factions on H. pylori Virulence Factors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cover, T.L.; Blaser, M.J. Helicobacter pylori in Health and Disease. Gastroenterology 2009, 136, 1863–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testerman, T.L.; Morris, J. Beyond the stomach: An updated view of Helicobacter pylori pathogenesis, diagnosis, and treatment. World J. Gastroenterol. 2014, 20, 12781–12808. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in Globocan 2012: Globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Safavi, M.; Shams-Ardakani, M.; Foroumadi, A. Medicinal plants in the treatment of Helicobacter pylori infections. Pharm. Biol. 2015, 53, 939–960. [Google Scholar] [CrossRef] [PubMed]
- Al-Eraky, D.M.; Helmy, O.M.; Ragab, Y.M.; Abdul-Khalek, Z.; El-Seidi, E.A.; Ramadan, M.A. Prevalence of CagA and antimicrobial sensitivity of H. pylori isolates of patients with gastric cancer in Egypt. Inf. Agents Cancer 2018, 13, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, U.; Fowora, M.A.; Seriki, A.T.; Loell, E.; Mueller, S.; Ugo-Ijeh, M.; Onyekwere, C.A.; Lesi, O.A.; Otegbayo, J.A.; Akere, A.; et al. Helicobacter pylori strains from a Nigerian cohort show divergent antibiotic resistance rates and a uniform pathogenicity profile. PLoS ONE 2017, 12, e0176454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndip, R.N.; Takang, A.E.M.; Ojongokpoko, J.E.A.; Luma, H.N.; Malongue, A.; Akoachere, J.-F.T.K.; Ndip, L.M.; MacMillan, M.; Weaver, L.T. Helicobacter pylori isolates recovered from gastric biopsies of patients with gastro-duodenal pathologies in Cameroon: Current status of antibiogram. Trop. Med. Int. Health 2008, 13, 848–854. [Google Scholar] [CrossRef]
- WHO Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 6 November 2019).
- Konieczna, I.; Żarnowiec, P.; Kwinkowski, M.; Kolesińska, B.; Frączyk, J.; Kamiński, Z.; Kaca, W. Bacterial Urease and its Role in Long-Lasting Human Diseases. Curr. Protein Pept. Sci. 2012, 13, 789–806. [Google Scholar] [CrossRef]
- Rutherford, J.C. The Emerging Role of Urease as a General Microbial Virulence Factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [Green Version]
- Hage, N.; Howard, T.; Phillips, C.; Brassington, C.; Overman, R.; Debreczeni, J.; Gellert, P.; Stolnik, S.; Winkler, G.S.; Falcone, F.H. Structural basis of Lewisb antigen binding by the Helicobacter pylori adhesin BabA. Sci. Adv. 2015, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kalali, B.; Mejías-Luque, R.; Javaheri, A.; Gerhard, M. H. pylori Virulence Factors: Influence on Immune System and Pathology. Med. Inflamm 2014, 2014, 426309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escandón, R.A.; del Campo, M.; López-Solis, R.; Obreque-Slier, E.; Toledo, H. Antibacterial effect of kaempferol and (−)-epicatechin on Helicobacter pylori. Eur. Food Res. Technol. 2016, 242, 1495–1502. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Žemlička, M. Plant-derived urease inhibitors as alternative chemotherapeutic agents. Archiv. Pharmazie 2016, 349, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Flavonoid glycosides of Polygonum capitatum protect against inflammation associated with Helicobacter pylori infection. PLoS ONE 2015, 10, e0126584.
- Ghasemian, A.; Fattahi, A.; Mostafavi, S.K.S.; Almarzoqi, A.H.; Memariani, M.; Braiek, O.B.; Yassine, H.M.; Mostafavi, N.S.S.; Ahmed, M.M.; Mirforughi, S.A. Herbal medicine as an auspicious therapeutic approach for the eradication of Helicobacter pylori infection: A concise review. J. Cell. Phys. 2019, 234, 16847–16860. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C. Medicinal plant activity on Helicobacter pylori related diseases. World J. Gastroenterol. 2014, 20, 10368–10382. [Google Scholar] [CrossRef]
- Ilodigwe, E.E.; Akah, P.A.; Nworu, C.S. Anticonvulsant activity of ethanol leaf extract of Spathodea campanulata P. Beauv (Bignoniaceae). J. Med. Food 2010, 13, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Ofori-Kwakye, K.; Kwapong, A.A.; Adu, F. Antimicrobial activity of extracts and topical products of the stem bark of Spathodea campanulata for wound healing. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Teinkela, J.E.M.; Nguedia, J.C.A.; Meyer, F.; Donfack, E.V.; Ndjakou, B.L.; Ngouela, S.; Tsamo, E.; Adiogo, D.; Azebaze, A.G.B.; Wintjens, R. In vitro antimicrobial and anti-proliferative activities of plant extracts from Spathodea campanulata, Ficus bubu, and Carica papaya. Pharm. Biol. 2016, 54, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Ngnameko, C.R.; Njayou, F.N.; Fowora, M.; Nemg, F.B.S.; Moundipa Fewou, P.; Smith, S.I. Inhibitory effect of medicinal plants from Cameroon on the growth and adhesion of Helicobacter pylori. Eur. J. Int. Med. 2019, 30, 100957. [Google Scholar] [CrossRef]
- Amusan, O.O.G.; Adesogan, E.K.; Makinde, J.M. Antimalarial Active Principles of Spathodea campanulata Stem Bark. Phytother. Res. 1996, 10, 692–693. [Google Scholar] [CrossRef]
- Boniface, P.K.; Singh, M.; Verma, S.; Shukla, A.; Khan, F.; Srivastava, S.K.; Pal, A. RP-HPLC-DAD method for the identification of two potential antioxidant agents namely verminoside and 1-O-(E)-caffeoyl-β-gentiobiose from Spathodea campanulata leaves. Nat. Prod. Res. 2015, 29, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Heim, S.C.; Guarnier, F.A.; Ferreira, D.T.; Braz-Filho, R.; Cecchini, R.; Cecchini, A.L. Antioxidant activity of Spathodea campanulata (Bignoneaceae) extracts. Rev. Bras. Plantas Med. 2012, 14, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Lifongo, L.L.; Simoben, C.V.; Ntie-Kang, F.; Babiaka, S.B.; Judson, P.N. A bioactivity versus ethnobotanical survey of medicinal plants from Nigeria, west Africa. Nat. Prod. Bioprospect. 2014, 4, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazif, N.M. Phytochemical and antioxidant activity of Spathodea campanulata P. beauvois. Growing in Egypt. Nat. Prod. Sci. 2007, 13, 11–16. [Google Scholar]
- Ngouela, S.; Tsamo, E.; Sondengam, B.L.; Connolly, J.D. Spathodol, a New Polyhydroxysterol from the Leaves of Spathodea campanulata. J. Nat. Prod. 1991, 54, 873–876. [Google Scholar] [CrossRef]
- Ngoungoure, V.L.N.; Mfotie Njoya, E.; Ngamli Fewou, S.; Ella, A.F.; McGaw, L.J.; Moundipa, P.F. Acetylcholinesterase inhibitory, anti-inflammatory and antioxidant properties of some Cameroonian medicinal plants used to treat some neurological disorders. Investig. Med. Chem. Pharmacol. 2019, 2, 33–46. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Pianaro, A.; Pinto, J.; Trevisan Ferreira, D.; Ishikawa, N.; Braz-Filho, R. Iridoid glucoside and antifungal phenolic compounds from Spathodea campanulata roots. Semin. Cienc. Agrar. 2007, 28, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Tarsia, C.; Danielli, A.; Florini, F.; Cinelli, P.; Ciurli, S.; Zambelli, B. Targeting Helicobacter pylori urease activity and maturation: In-cell high-throughput approach for drug discovery. Biochim. Biophsica Acta (BBA) Gen. Subj. 2018, 1862, 2245–2253. [Google Scholar] [CrossRef]
- Godstime, O.C.; García, A.; Augustina, J.O.; Christopher, E. Mechanisms of Antimicrobial Actions of Phytochemicals against Enteric Pathogens—A Review. J. Pharm. Chem. Biol. Sci. 2014, 2, 77–85. [Google Scholar]
- Yeon, M.J.; Lee, M.H.; Kim, D.H.; Yang, J.Y.; Woo, H.J.; Kwon, H.J.; Moon, C.; Kim, S.-H.; Kim, J.-B. Anti-inflammatory effects of Kaempferol on Helicobacter pylori-induced inflammation. Biosci. Biotechnol. Biochem. 2019, 83, 166–173. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Salillas, S.; Velázquez-Campoy, A.; Espinosa Angarica, V.; Fillat, M.F.; Sancho, J.; Lanas, Á. Identifying potential novel drugs against Helicobacter pylori by targeting the essential response regulator HsrA. Sci. Rep. 2019, 9, 11294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehues, M.; Stark, T.; Keller, D.; Hofmann, T.; Hensel, A. Antiadhesion as a functional concept for prevention of pathogens: N-Phenylpropenoyl-L-amino acid amides as inhibitors of the Helicobacter pylori BabA outer membrane protein. Mol. Nutr. Food Res. 2011, 55, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Racha, S.; Wongrattanakamon, P.; Raiwa, A.; Jiranusornkul, S. Discovery of novel potent small natural molecules able to enhance attenuation of the pathobiology of gastric cancer-associated Helicobacter pylori by molecular modeling. Int. J. Pept. Res. Ther. 2019, 25, 881–896. [Google Scholar] [CrossRef]
- Kataoka, M.; Hirata, K.; Kunikata, T.; Ushio, S.; Iwaki, K.; Ohashi, K.; Ikeda, M.; Kurimoto, M. Antibacterial action of tryptanthrin and kaempferol, isolated from the indigo plant (Polygonum tinctorium Lour.), against Helicobacter pylori-infected Mongolian gerbils. J. Gastroenterol. 2001, 36, 5–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Polyphenols (mg GAE/g) a | ||||||
| Crude extract | A | B | C | D | E | F |
| 51.4 b | 46.4 ± 0.5 | 43.0 ± 0.1 | 53.0 b | 48.3 b | 68.8 ± 0.3 | 56.3 ± 1.1 |
| Total Flavonoids (mg QE/g) a | ||||||
| Crude extract | A | B | C | D | E | F |
| 5.2 ± 0.2 | 2.8 ± 0.1 | 1.8 b | 1.9 b | 2.0 b | 7.1 b | 1.9 ± 0.1 |
| Compound | Structure | Fraction/Sub-Fraction | |||||||
|---|---|---|---|---|---|---|---|---|---|
| C | E | SA1 | SB1 | SC2 | SD2 | SE1 | SE3 | ||
| 4-Hydroxy Benzoic Acid |  | ||||||||
| Methyl-4-Hydroxybenzoate |  | ||||||||
| 5,7-Dihydroxy-4-Metilcoumarin |  | ||||||||
| Quercetin |  | ||||||||
| Kaempferol |  | ||||||||
| Kaempferol 3-O-Glucoside |  | ||||||||
| Spathodol |  | ||||||||
| Spathodic Acid |  | ||||||||
| Tomentosolic Acid |  | ||||||||
| Ursolic Acid |  | ||||||||
| Corosolic Acid |  | ||||||||
| Sample. | Concentration | |||
|---|---|---|---|---|
| 0.1 mg/mL | 1.0 mg/mL | 10.0 mg/mL | MAQ (mg/mL) | |
| Crude extract | ND | 8.6 ± 0.8 | 11.6 ± 0.7 | 1.0 |
| B | 11.3 ± 1.2 | 11.7 ± 2.1 | 12.1 ± 1.0 | 0.1 |
| C | 10.0 | 12.0 | 16.3 ± 2.5 | 0.1 |
| E | 10.0 | 11.7 ± 0.6 | 15.8 ± 0.3 | 0.1 |
| SA1 | 9.8 ± 0.4 | 10.0 ± 1.0 | 10.7 ± 1.2 | 0.1 |
| SB1 | 9.3 ± 0.6 | 9.7 ± 0.6 | 10.7 ± 0.6 | 0.1 |
| SB2 | ND | 8.7 ± 2.3 | 11.3 ± 2.5 | 1.0 |
| SC2 | ND | 8.0 | 10.0 | 1.0 |
| SD1 | ND | ND | 10.3 ± 0.6 | 10.0 |
| SE3 | ND | ND | 13.0 ± 1.0 | 10.0 |
| SF | ND | 9.3 ± 1.2 | 10.7 ± 0.6 | 1.0 |
| Kanamycin (2.5 mg/mL) | 35.7 ± 0.6 | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngnameko, C.R.; Marchetti, L.; Zambelli, B.; Quotadamo, A.; Roncarati, D.; Bertelli, D.; Njayou, F.N.; Smith, S.I.; Moundipa, P.F.; Costi, M.P.; et al. New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori. Antibiotics 2020, 9, 258. https://doi.org/10.3390/antibiotics9050258
Ngnameko CR, Marchetti L, Zambelli B, Quotadamo A, Roncarati D, Bertelli D, Njayou FN, Smith SI, Moundipa PF, Costi MP, et al. New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori. Antibiotics. 2020; 9(5):258. https://doi.org/10.3390/antibiotics9050258
Chicago/Turabian StyleNgnameko, Corinne Raïssa, Lucia Marchetti, Barbara Zambelli, Antonio Quotadamo, Davide Roncarati, Davide Bertelli, Frederic Nico Njayou, Stella I. Smith, Paul F. Moundipa, Maria Paola Costi, and et al. 2020. "New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori" Antibiotics 9, no. 5: 258. https://doi.org/10.3390/antibiotics9050258
APA StyleNgnameko, C. R., Marchetti, L., Zambelli, B., Quotadamo, A., Roncarati, D., Bertelli, D., Njayou, F. N., Smith, S. I., Moundipa, P. F., Costi, M. P., & Pellati, F. (2020). New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori. Antibiotics, 9(5), 258. https://doi.org/10.3390/antibiotics9050258