
A Novel Ferroptosis-Related Gene Signature Predicts Overall Survival of Breast Cancer Patients


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. TCGA-BRCA Cohort and METABRIC Cohort
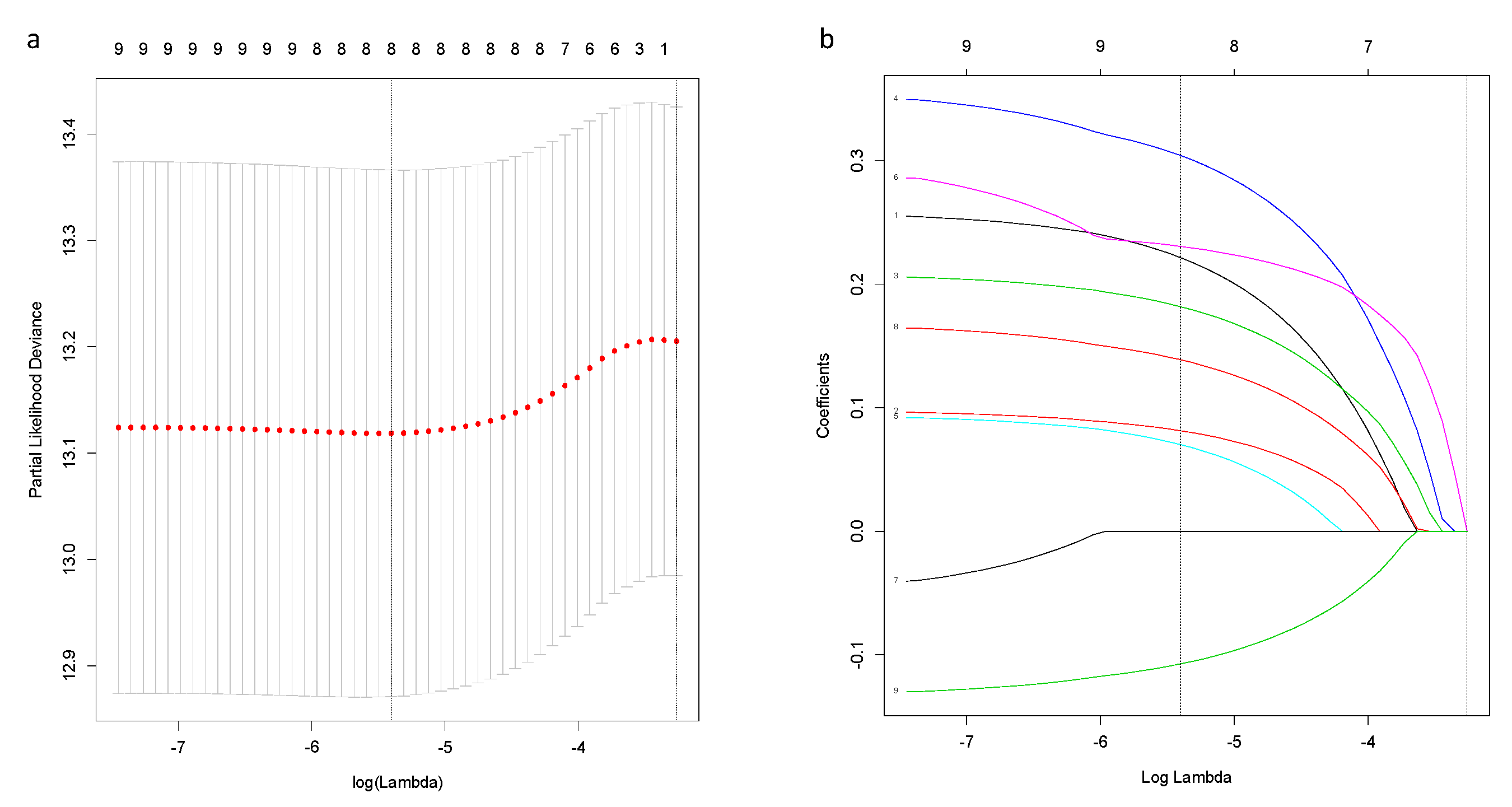
2.2. Construction and Validation of a Novel Prognostic Ferroptosis-Related Gene Signature
2.3. The Tumor Microenvironment Analysis
2.4. Statistical Analysis
3. Results
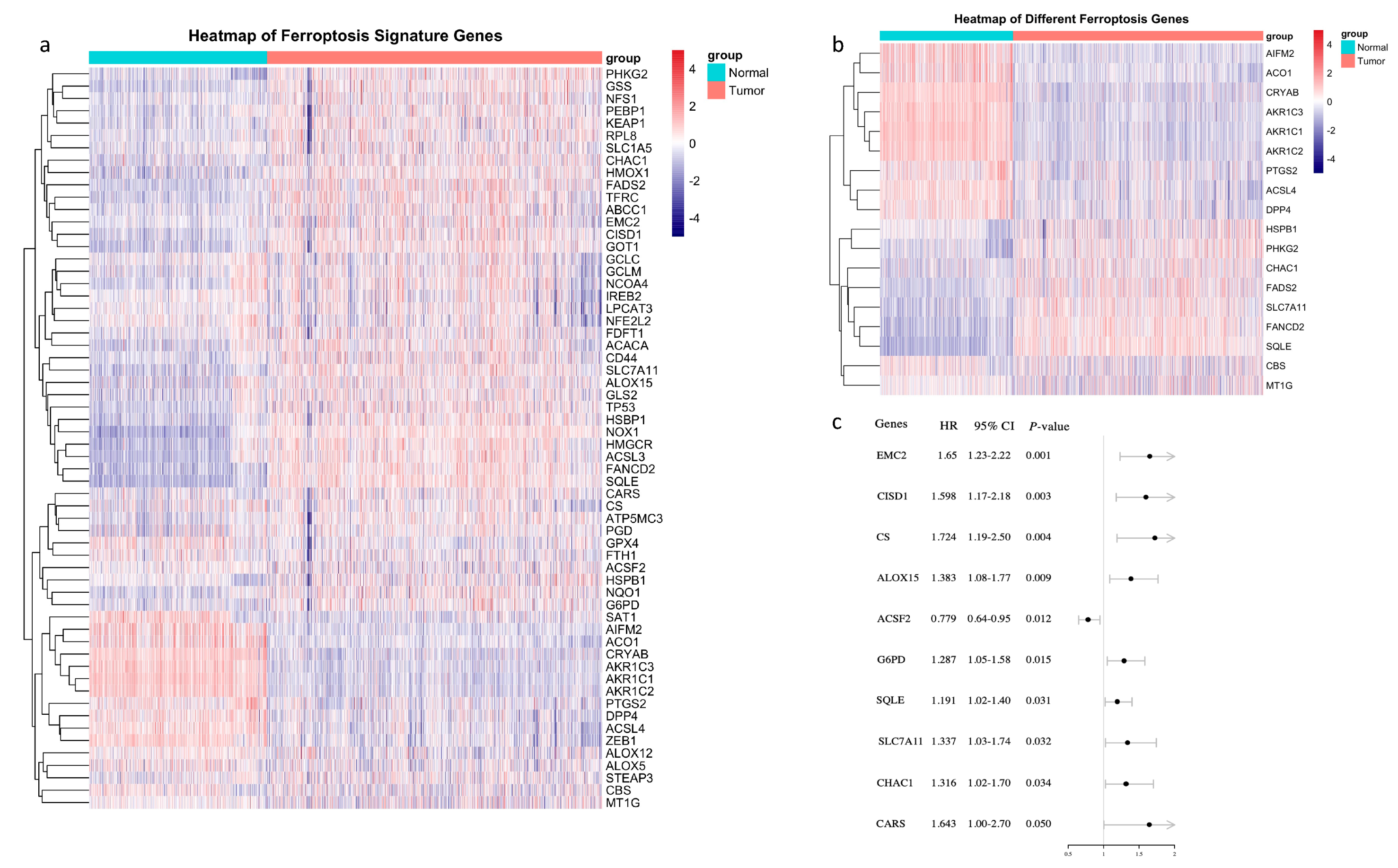
3.1. Identification of Prognostic Ferroptosis-Related Genes in the TCGA-BRCA Cohort
3.2. A Prognostic Model Construction in the TCGA-BRCA Cohort
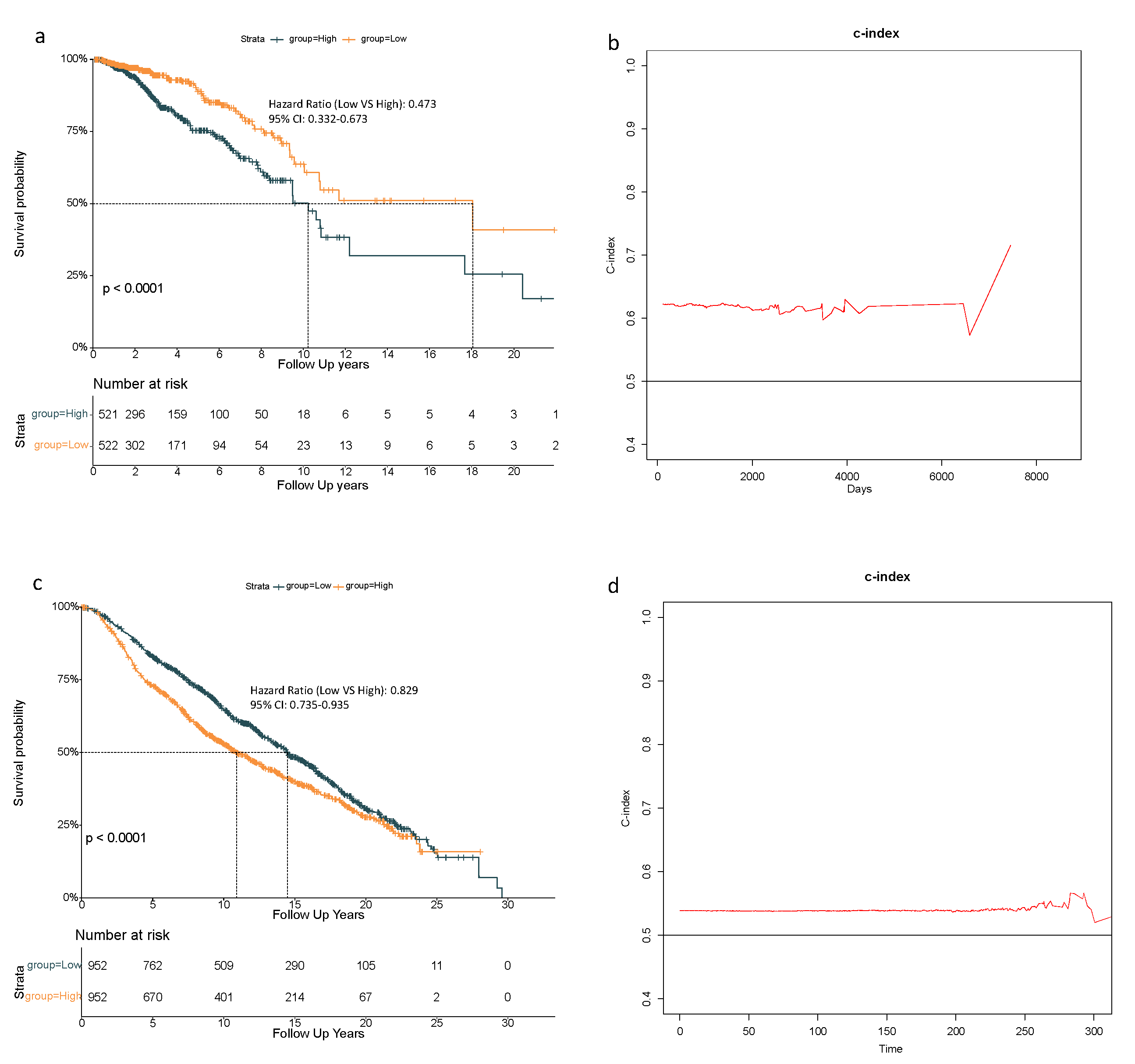
3.3. 8-Gene Signature Validation in the METABRIC Cohort
3.4. Independent Prognostic Value of the 8-Gene Signature
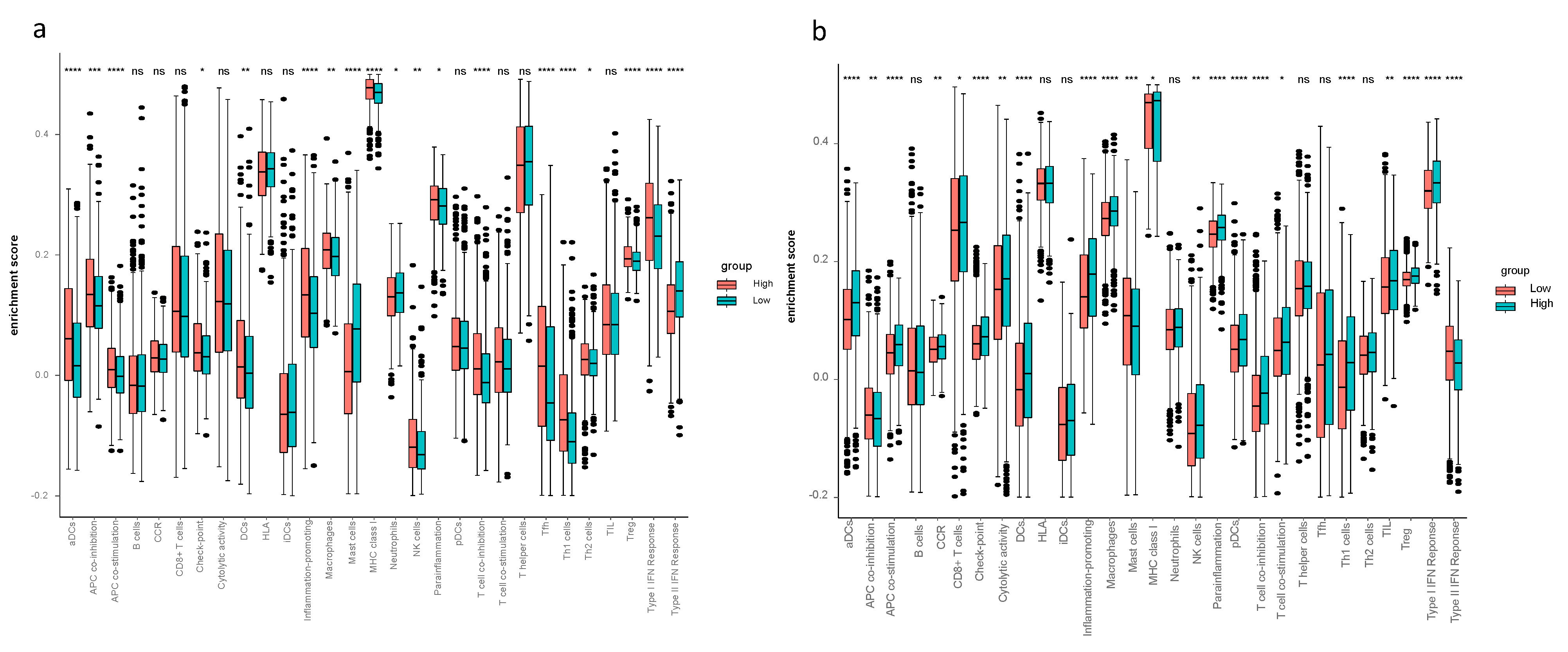
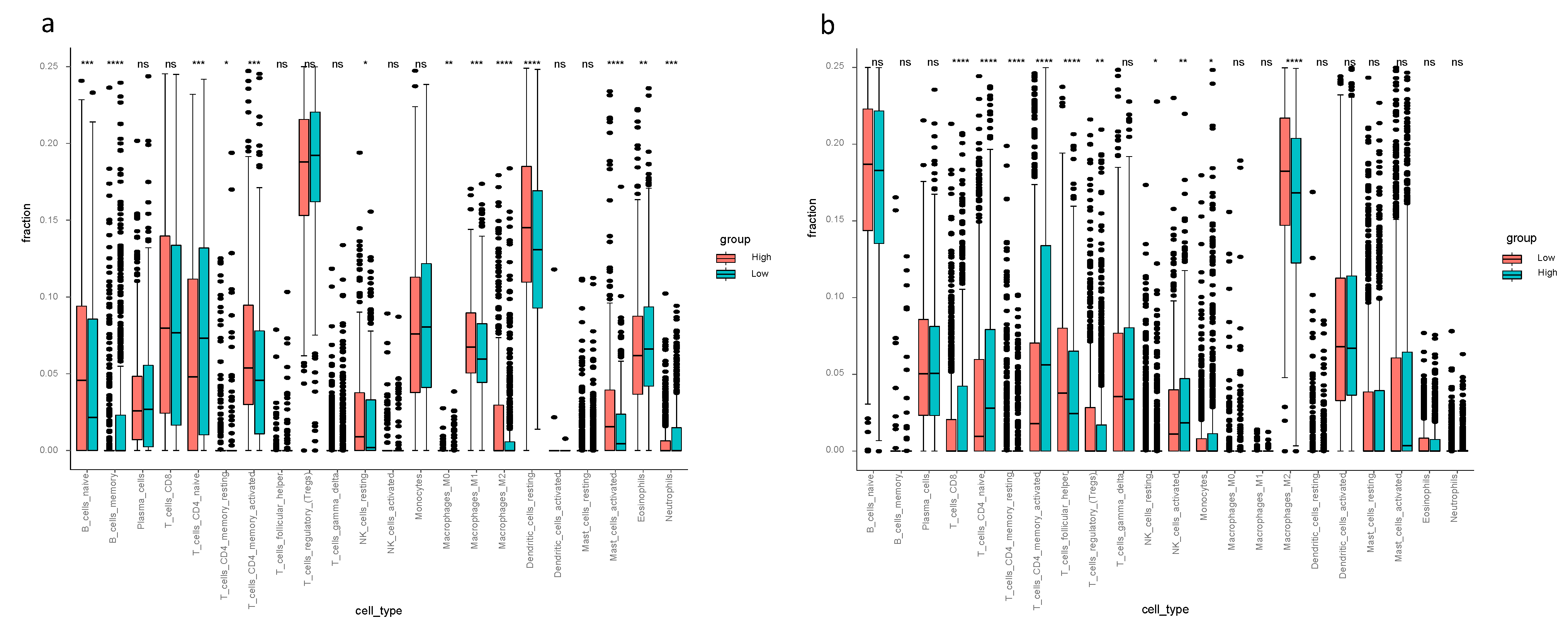
3.5. The Tumor Microenvironment Analysis in the TCGA-BRCA and METABRIC Cohort
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Diaby, V.; Tawk, R.; Sanogo, V.; Xiao, H.; Montero, A.J. A review of systematic reviews of the cost-effectiveness of hormone therapy, chemotherapy, and targeted therapy for breast cancer. Breast Cancer Res. Treat. 2015, 151, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Ferrini, K.; Ghelfi, F.; Mannucci, R.; Titta, L. Lifestyle, nutrition and breast cancer: Facts and presumptions for consideration. Ecancermedicalscience 2015, 9. [Google Scholar] [CrossRef] [Green Version]
- Donepudi, M.S.; Kondapalli, K.; Amos, S.J.; Venkanteshan, P. Breast cancer statistics and markers. J. Cancer Res. Ther. 2014, 10, 506–511. [Google Scholar] [PubMed]
- Maughan, K.L.; Lutterbie, M.A.; Ham, P.S. Treatment of breast cancer. Am. Fam. Physician 2010, 81, 1339–1346. [Google Scholar] [PubMed]
- Pu, M.; Messer, K.; Davies, S.R.; Vickery, T.L.; Pittman, E.; Parker, B.A.; Ellis, M.J.; Flatt, S.W.; Marinac, C.R.; Nelson, S.H.; et al. Research-based PAM50 signature and long-term breast cancer survival. Breast Cancer Res. Treat. 2020, 179, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Henson, E.S.; Chen, Y.; Gibson, S.B. Ferroptosis is induced following siramesine and lapatinib treatment of breast cancer cells. Cell Death Dis. 2016, 7, e2307. [Google Scholar] [CrossRef] [Green Version]
- Reddel, R.R.; Hedley, D.W.; Sutherland, R.L. Cell cycle effects of iron depletion on T-47D human breast cancer cells. Exp. Cell Res. 1985, 161, 277–284. [Google Scholar] [CrossRef]
- Rao, V.A.; Klein, S.R.; Agama, K.K.; Toyoda, E.; Adachi, N.; Pommier, Y.; Shacter, E.B. The iron chelator Dp44mT causes DNA damage and selective inhibition of topoisomerase IIalpha in breast cancer cells. Cancer Res. 2009, 69, 948–957. [Google Scholar] [CrossRef] [Green Version]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Hangauer, M.J.; Viswanathan, V.S.; Ryan, M.J.; Bole, D.; Eaton, J.K.; Matov, A.; Galeas, J.; Dhruv, H.D.; Berens, M.E.; Schreiber, S.L.; et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 2017, 551, 247–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Zhu, S.; Song, X.; Sun, X.; Fan, Y.; Liu, J.; Zhong, M.; Yuan, H.; Zhang, L.; Billiar, T.R.; et al. The Tumor Suppressor p53 Limits Ferroptosis by Blocking DPP4 Activity. Cell Rep. 2017, 20, 1692–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Takahashi, H.; Rajabi, H.; Alam, M.; Suzuki, Y.; Yin, L.; Tagde, A.; Maeda, T.; Hiraki, M.; Sukhatme, V.P.; et al. Functional interactions of the cystine/glutamate antiporter, CD44v and MUC1-C oncoprotein in triple-negative breast cancer cells. Oncotarget 2016, 7, 11756–11769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.-J.; Hu, H.-M.; Li, G.-Z.; Zhang, Y.; Wu, F.; Liu, X.; Wang, K.-Y.; Zhang, C.-B.; Jiang, T. Ferroptosis-Related Gene Signature Predicts Glioma Cell Death and Glioma Patient Progression. Front. Cell Dev. Biol. 2020, 8, 538. [Google Scholar] [CrossRef]
- Kwon, O.-S.; Kwon, E.-J.; Kong, H.-J.; Choi, J.-Y.; Kim, Y.-J.; Lee, E.-W.; Kim, W.; Lee, H.; Cha, H.-J. Systematic identification of a nuclear receptor-enriched predictive signature for erastin-induced ferroptosis. Redox Biol. 2020, 37, 101719. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.-Y.; Wang, D.-S.; Lin, H.-C.; Chen, X.-X.; Yang, H.; Zheng, Y.; Li, Y.-H. A Novel Ferroptosis-related Gene Signature for Overall Survival Prediction in Patients with Hepatocellular Carcinoma. Int. J. Biol. Sci. 2020, 16, 2430–2441. [Google Scholar] [CrossRef] [PubMed]
- Hassannia BV, P.; Vanden Berghe, T. Targeting Ferroptosis to Iron Out Cancer. Cancer Cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Han, C.; Liu, Y.; Dai, R.; Ismail, N.; Su, W.; Li, B. Ferroptosis and Its Potential Role in Human Diseases. Front. Pharmacol. 2020, 11, 239. [Google Scholar] [CrossRef]
- Shi, Z.Z.; Fan, Z.W.; Chen, Y.X.; Xie, X.F.; Jiang, W.; Wang, W.J.; Qiu, Y.T.; Bai, J. Ferroptosis in Carcinoma: Regulatory Mecha-nisms and New Method for Cancer Therapy. Onco Targets Ther. 2019, 12, 11291–11304. [Google Scholar] [CrossRef] [Green Version]
- Yagoda, N.; Von Rechenberg, M.; Zaganjor, E.; Bauer, A.J.; Yang, W.S.; Fridman, D.J.; Wolpaw, A.J.; Smukste, I.; Peltier, J.M.; Boniface, J.J.; et al. RAS–RAF–MEK-dependent oxidative cell death involving voltage-dependent anion channels. Nature 2007, 447, 865–869. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Kong, N.; Li, T.; Wang, S.J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.J.; Patel, D.N.; Welsch, M.; Skouta, R.; Lee, E.D.; Hayano, M.; Thomas, A.G.; Gleason, C.E.; Tatonetti, N.P.; Slusher, B.S.; et al. Pharmacological inhibition of cystine–glutamate exchange induces endoplasmic reticulum stress and ferroptosis. eLife 2014, 3, e02523. [Google Scholar] [CrossRef]
- Bebber, C.M.; Müller, F.; Prieto Clemente, L.; Weber, J.; Von Karstedt, S. Ferroptosis in Cancer Cell Biology. Cancers(Basel) 2020, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Pedro, J.M.B.-S.; Kroemer, G. Ferroptosis in p53-dependent oncosuppression and organismal homeostasis. Cell Death Differ. 2015, 22, 1237–1238. [Google Scholar] [CrossRef]
- Yuan, H.; Li, X.; Zhang, X.; Kang, R.; Tang, D. CISD1 inhibits ferroptosis by protection against mitochondrial lipid peroxidation. Biochem. Biophys. Res. Commun. 2016, 478, 838–844. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Xu, Z.; Zhu, Z.; Chen, A.; Fu, G.; Wang, Y.; Pan, H.; Jin, B. Modulation of G6PD affects bladder cancer via ROS accumulation and the AKT pathway in vitro. Int. J. Oncol. 2018, 53, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.-Q.; Lu, Y.-X.; Wu, Q.-N.; Liu, J.; Zeng, Z.-L.; Mo, H.-Y.; Chen, Y.; Tian, T.; Wang, Y.; Kang, T.-B.; et al. Disrupting G6PD-mediated Redox homeostasis enhances chemosensitivity in colorectal cancer. Oncogene 2017, 36, 6282–6292. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Zhang, Y.; Tang, Q.; Jin, L.; Chen, Y. SQLE induces epithelial-to-mesenchymal transition by regulating of miR-133b in esophageal squamous cell carcinoma. Acta Biochim. et Biophys. Sin.(Shanghai) 2017, 49, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Lang, X.; Green, M.D.; Wang, W.; Yu, J.; Choi, J.E.; Jiang, L.; Liao, P.; Zhou, J.; Zhang, Q.; Dow, A.; et al. Radiotherapy and Immunotherapy Promote Tumoral Lipid Oxidation and Ferroptosis via Synergistic Repression of SLC7A. Cancer Discov. 2019, 9, 1673–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.-S.; Wang, S.-F.; Hsu, C.-Y.; Yin, P.-H.; Yeh, T.-S.; Lee, H.-C.; Tseng, L.-M. CHAC1 degradation of glutathione enhances cystine-starvation-induced necroptosis and ferroptosis in human triple negative breast cancer cells via the GCN2-eIF2α-ATF4 pathway. Oncotarget 2017, 8, 114588–114602. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; He, Y.; Luo, N.; Patel, S.J.; Han, Y.; Gao, R.; Modak, M.; Carotta, S.; Haslinger, C.; Kind, D.; et al. Landscape and Dynamics of Single Immune Cells in Hepatocellular Carcinoma. Cell 2019, 179, 829–845.e20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-L.; Zhou, Z.-J.; Hu, Z.-Q.; Huang, X.-W.; Wang, Z.; Chen, E.-B.; Fan, J.; Cao, Y.; Dai, Z.; Zhou, J. Tumor-Associated Neutrophils Recruit Macrophages and T-Regulatory Cells to Promote Progression of Hepatocellular Carcinoma and Resistance to Sorafenib. Gastroenterology 2016, 150, 1646–1658.e17. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Xu, D.; Liu, Z.; Shi, M.; Zhao, P.; Fu, B.; Zhang, Z.; Yang, H.; Zhang, H.; Zhou, C.; et al. Increased Regulatory T Cells Correlate with CD8 T-Cell Impairment and Poor Survival in Hepatocellular Carcinoma Patients. Gastroenterology 2007, 132, 2328–2339. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: New molecules and patterns of gene expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [Green Version]
- Nucera, S.; Biziato, D.; De Palma, M. The interplay between macrophages and angiogenesis in development, tissue injury and regeneration. Int. J. Dev. Biol. 2011, 55, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | TCGA-BRCA | METABRIC |
|---|---|---|
| N | 1043 | 1904 |
| Age (mean (SD)) | 58.33 (13.20) | 61.09 (12.98) |
| Gender (%) | ||
| Male | 12 (1.2) | - |
| Female | 1031 (98.8) | - |
| Race (%) | ||
| Asian | 57 (5.5) | - |
| Black | 178 (17.1) | - |
| White | 727 (69.7) | - |
| NA | 81 (7.8) | - |
| Stage (%) | ||
| I | 178 (17.1) | - |
| II | 589 (56.5) | - |
| III | 234 (22.4) | - |
| IV | 19 (1.8) | - |
| Menopause status (%) | ||
| Pre | 219 (21.0) | 411 (21.6) |
| peri | 72 (6.9) | 0 (0) |
| Post | 664 (63.7) | 1493 (78.4) |
| NA | 88 (8.4) | 0 (0) |
| Chemotherapy (%) | ||
| Yes | - | 396 (20.8) |
| No | - | 1508 (79.2) |
| Pathologic N (%) | ||
| < = N1 | - | 1434 (75.3) |
| > = N2 | - | 470 (24.7) |
| Characteristics | TCGA-BRCA | METABRIC | ||||
|---|---|---|---|---|---|---|
| High-Risk (n = 521) | Low-Risk (n = 522) | p Value | High-Risk (n = 952) | Low-Risk (n = 952) | p Value | |
| Age (mean (SD)) | 58.34 (13.56) | 58.32 (12.84) | 0.983 | 60.66 (13.29) | 61.51 (12.65) | 0.151 |
| Gender (%) | 0.146 | - | ||||
| Male | 9 (1.7) | 3 (0.6) | - | - | ||
| Female | 512(98.3) | 519(99.4) | - | - | ||
| Race (%) | 0.001 | - | ||||
| Black | 94 (18.0) | 84 (16.1) | - | - | ||
| White | 338 (64.9) | 389 (74.5) | - | - | ||
| Asian | 38 (7.3) | 19 (3.6) | - | - | ||
| NA | 51 (9.8) | 30 (5.7) | - | - | ||
| Stage (%) | 0.401 | - | ||||
| I+II | 376(72.2) | 391(74.9) | - | - | ||
| III+IV | 135 (25.9) | 118 (22.6) | - | - | ||
| NA | 10(1.9) | 13(2.5) | - | - | ||
| Menopause status (%) | 0.201 | 0.373 | ||||
| Pre | 117 (22.5) | 102 (19.5) | 214 (22.5) | 197 (20.7) | ||
| post | 331 (63.5) | 333 (63.8) | 738(77.5) | 755(79.3) | ||
| peri | 28 (5.4) | 44 (8.4) | - | - | ||
| NA | 45 (8.6) | 43 (8.2) | - | - | ||
| Chemotherapy (%) | - | <0.001 | ||||
| Yes | - | - | 265 (27.8) | 131 (13.8) | ||
| No | - | - | 687 (72.2) | 821 (86.2) | ||
| Pathologic N (%) | 0.017 | |||||
| < =N1 | 694 (72.9) | 740 (77.7) | ||||
| > =N2 | 258 (27.1) | 212 (22.3) | ||||
| Predictors | TCGA-BRCA METABRIC | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Univariate Analysis | Multivariate Analysis | Univariate Analysis | Multivariate Analysis | |||||||||
| HR | 95% CI | p | HR | 95% CI | p | HR | 95% CI | p | HR | 95% CI | p | |
| Age | 1.032 | 1.02-1.045 | <0.001 | 1.035 | 1.022–1.048 | <0.001 | 1.036 | 1.03–1.041 | <0.001 | 1.054 | 1.046–1.062 | <0.001 |
| Gender (Male vs. Female) | 0.84 | 0.1168–5.993 | 0.859 | - | - | - | - | - | - | - | - | - |
| Race | ||||||||||||
| Black (Ref) | ||||||||||||
| White | 0.826 | 0.552–1.236 | 0.353 | - | - | - | - | - | - | - | - | - |
| Asian | 0.617 | 0.188–2.029 | 0.427 | - | - | - | - | - | - | - | - | - |
| Stage (III/IV vs. I/II) | 2.665 | 1.905–3.730 | <0.001 | 2.84 | 2.024–3.982 | <0.001 | - | - | - | - | - | - |
| Menopause status | ||||||||||||
| Pre (Ref) | ||||||||||||
| Post | 1.279 | 0.842–1.943 | 0.248 | - | - | - | 1.685 | 1.431–1.983 | <0.001 | 0.633 | 0.501–0.799 | <0.001 |
| Peri | 0.906 | 0.457–1.794 | 0.776 | - | - | - | - | - | - | - | - | - |
| Group (Low vs. High) | 0.515 | 0.369–0.720 | <0.001 | 0.473 | 0.332–0.673 | <0.001 | 0.780 | 0.693–0.878 | <0.001 | 0.829 | 0.735–0.935 | <0.01 |
| Chemotherapy (Yes vs. No) | - | - | - | - | - | - | 1.228 | 1.057–1.427 | 0.007 | 1.741 | 1.450–2.091 | <0.001 |
| Pathologic N (> = N2 vs. < = N1) | - | - | - | - | - | - | 1.952 | 1.714–2.222 | <0.001 | 1.705 | 1.487–1.955 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Li, L.; Xue, C.; Huang, R.; Hu, A.; An, X.; Shi, Y. A Novel Ferroptosis-Related Gene Signature Predicts Overall Survival of Breast Cancer Patients. Biology 2021, 10, 151. https://doi.org/10.3390/biology10020151
Li H, Li L, Xue C, Huang R, Hu A, An X, Shi Y. A Novel Ferroptosis-Related Gene Signature Predicts Overall Survival of Breast Cancer Patients. Biology. 2021; 10(2):151. https://doi.org/10.3390/biology10020151
Chicago/Turabian StyleLi, Haifeng, Lu Li, Cong Xue, Riqing Huang, Anqi Hu, Xin An, and Yanxia Shi. 2021. "A Novel Ferroptosis-Related Gene Signature Predicts Overall Survival of Breast Cancer Patients" Biology 10, no. 2: 151. https://doi.org/10.3390/biology10020151
APA StyleLi, H., Li, L., Xue, C., Huang, R., Hu, A., An, X., & Shi, Y. (2021). A Novel Ferroptosis-Related Gene Signature Predicts Overall Survival of Breast Cancer Patients. Biology, 10(2), 151. https://doi.org/10.3390/biology10020151