
Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review

 ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
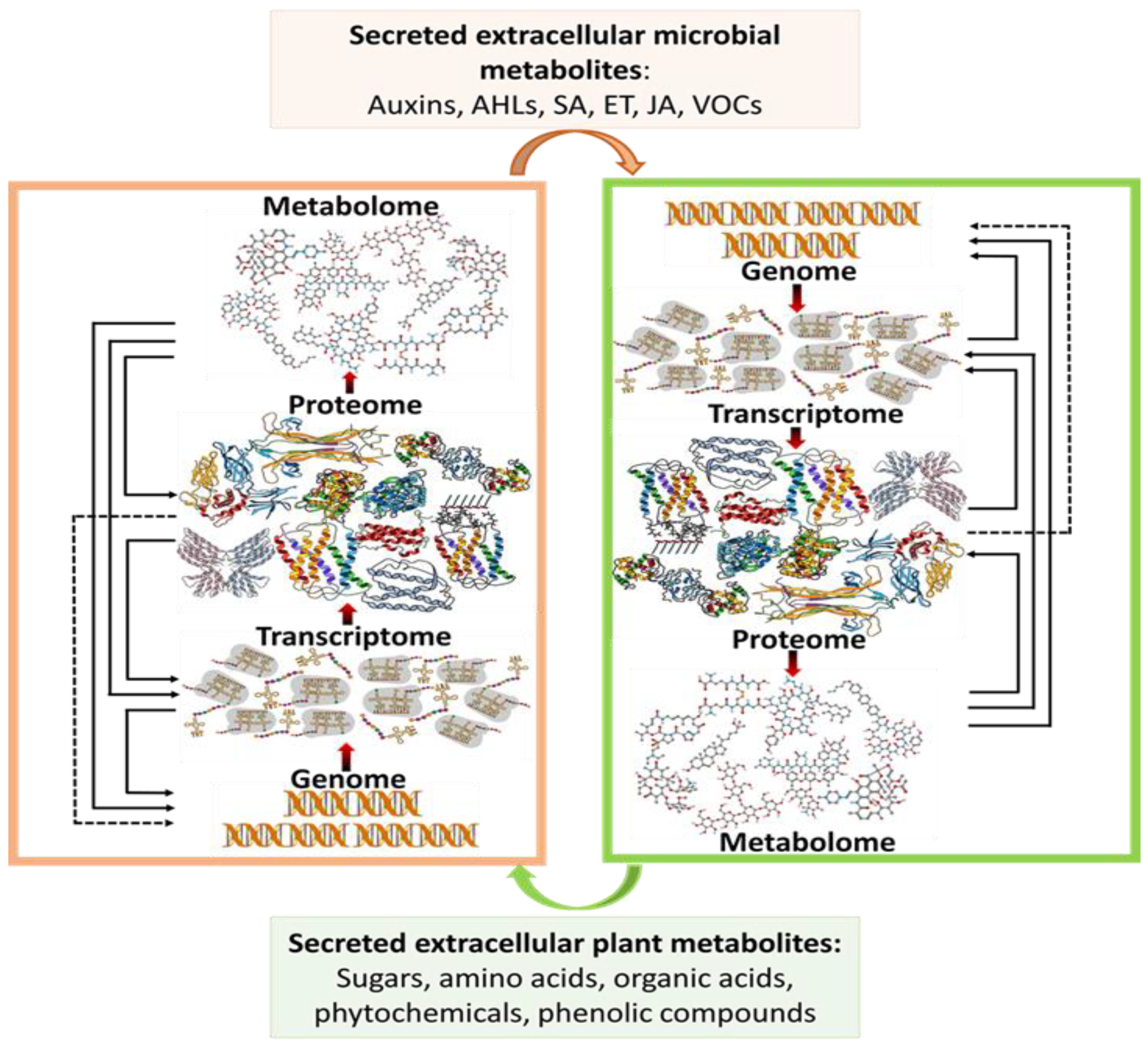
2. Metabolomics as a Prospect in Integrated Systems Biology
3. Rhizosphere Metabolomics: Rhizodeposits as Agents of Rhizosphere Selective Pressure and Current Advances in Rhizosphere Exometabolite Profiling
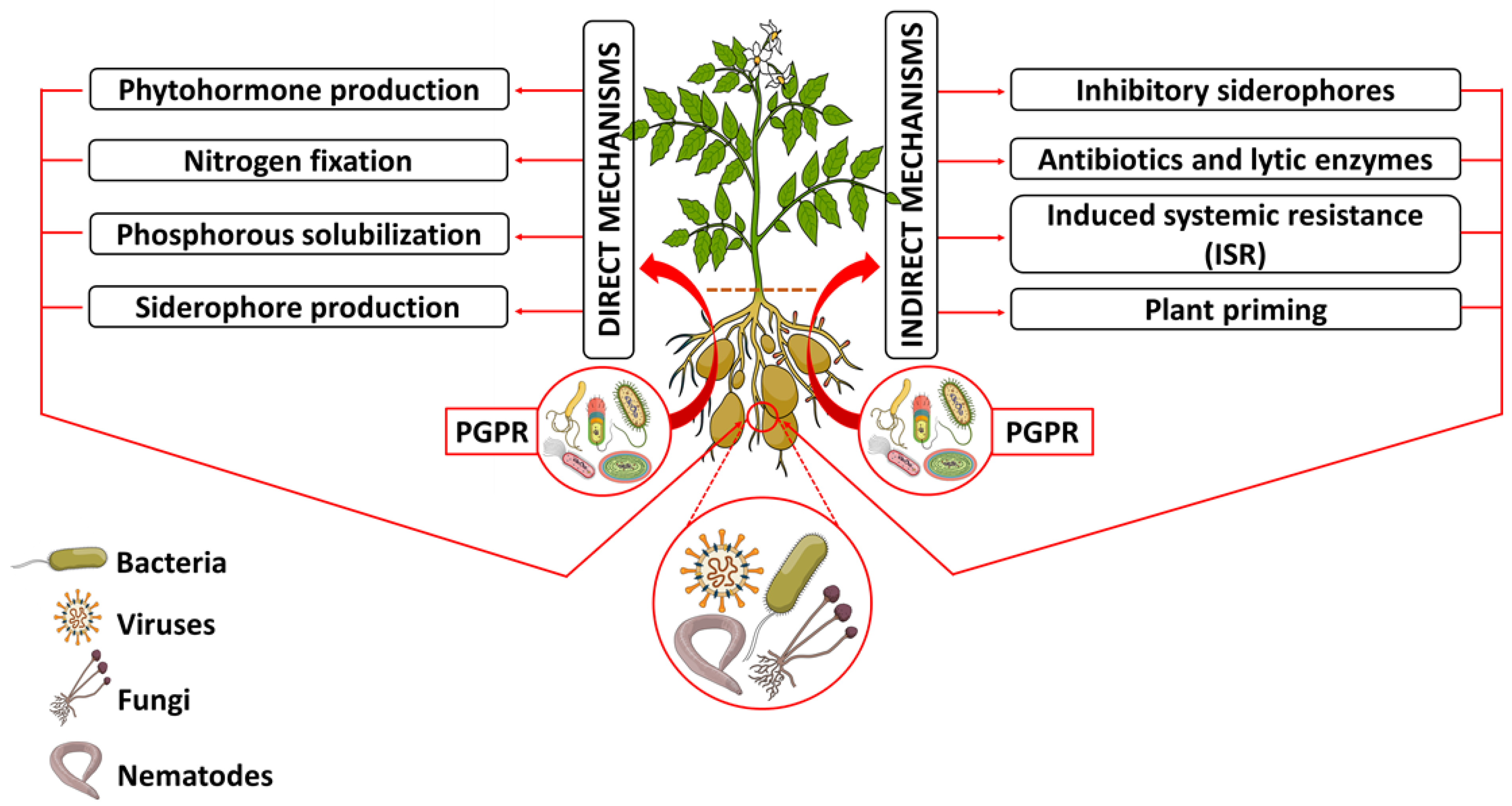
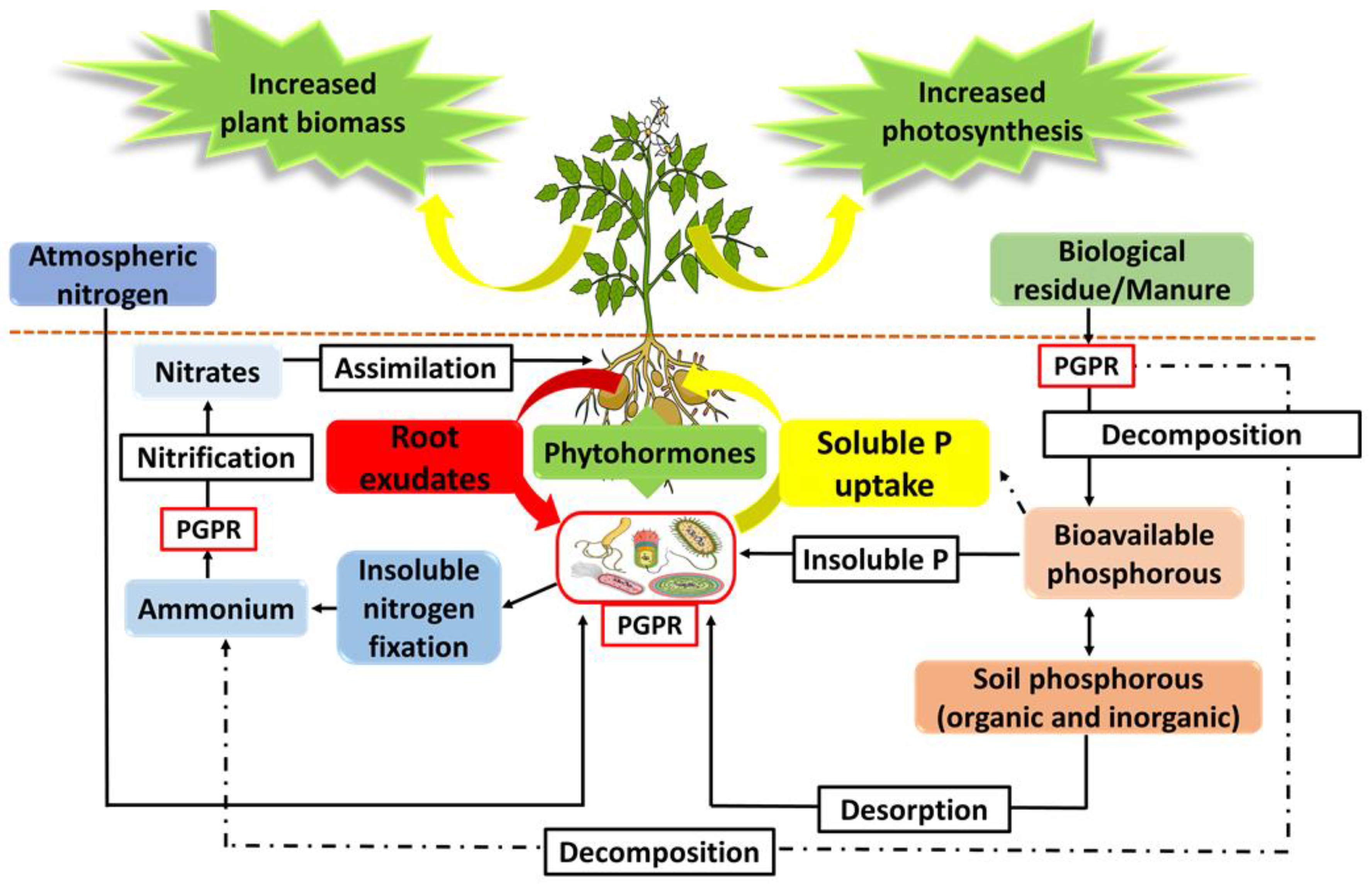
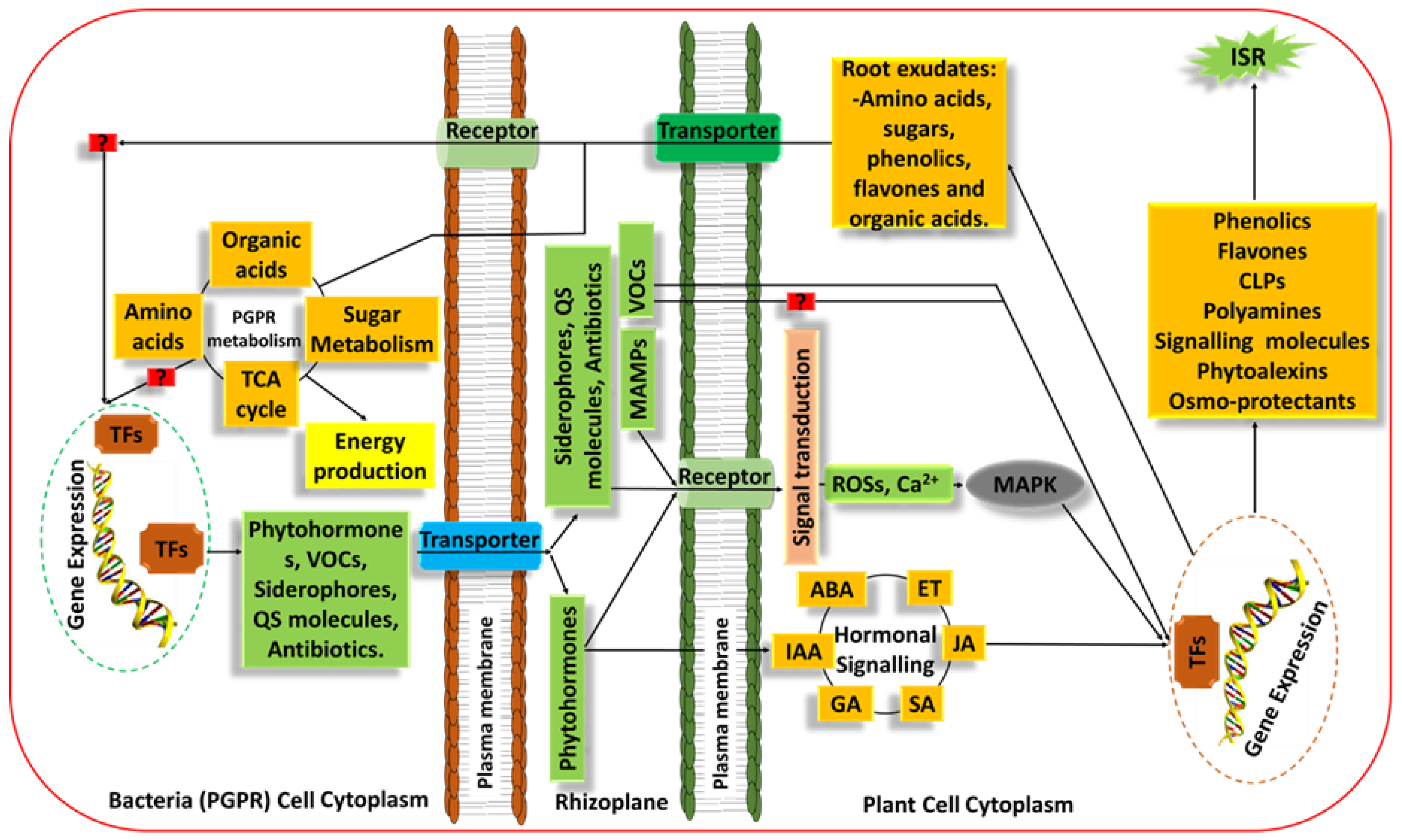
3.1. Rhizosphere Plant–PGPR Communications and PGPR Mechanisms of Action
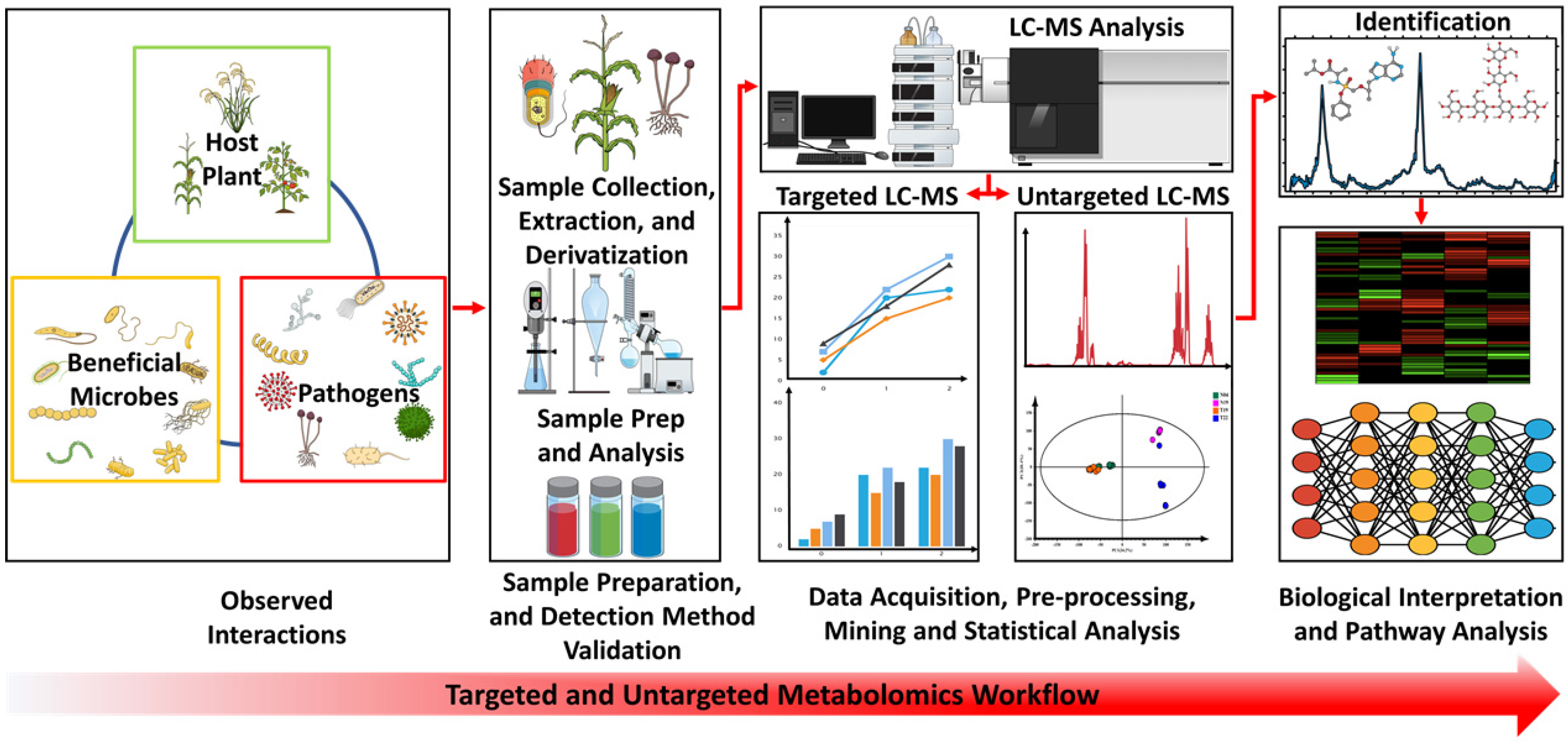
3.2. Rhizosphere Metabolomics: Current Methods and Applications
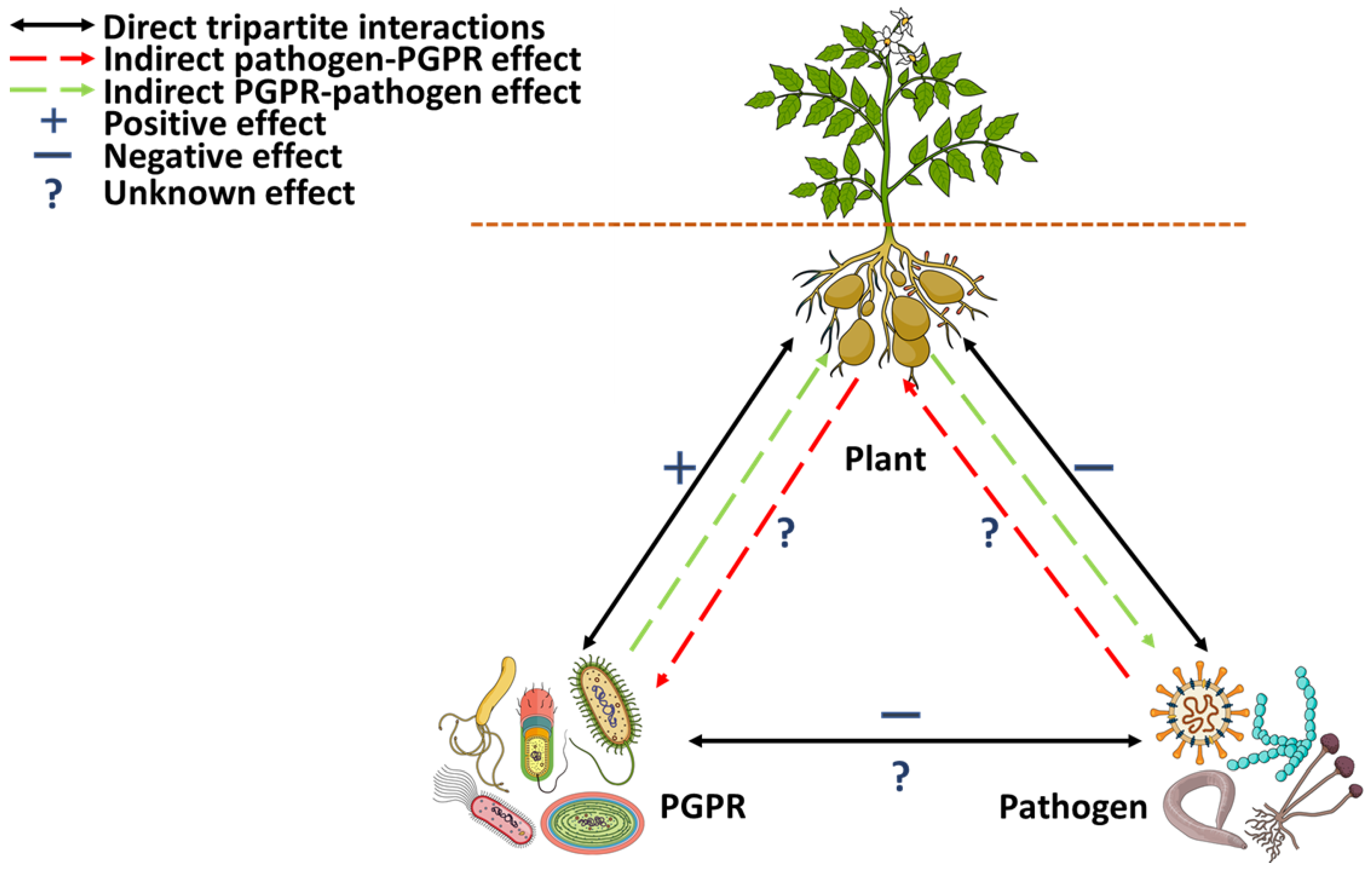
4. Metabolomics in Understanding Microbe-Induced Plant Metabolite Perturbations and the Potential for Deciphering Plant–Microbe Tripartite Interactions: Challenges and Prospects
4.1. Metabolomics in the Elucidation of PGPR-Induced Plant Metabolite Perturbations
4.2. Applications of Metabolomics in Understanding Tripartite Plant–Microbe Interactions: Current Advances and Challenges
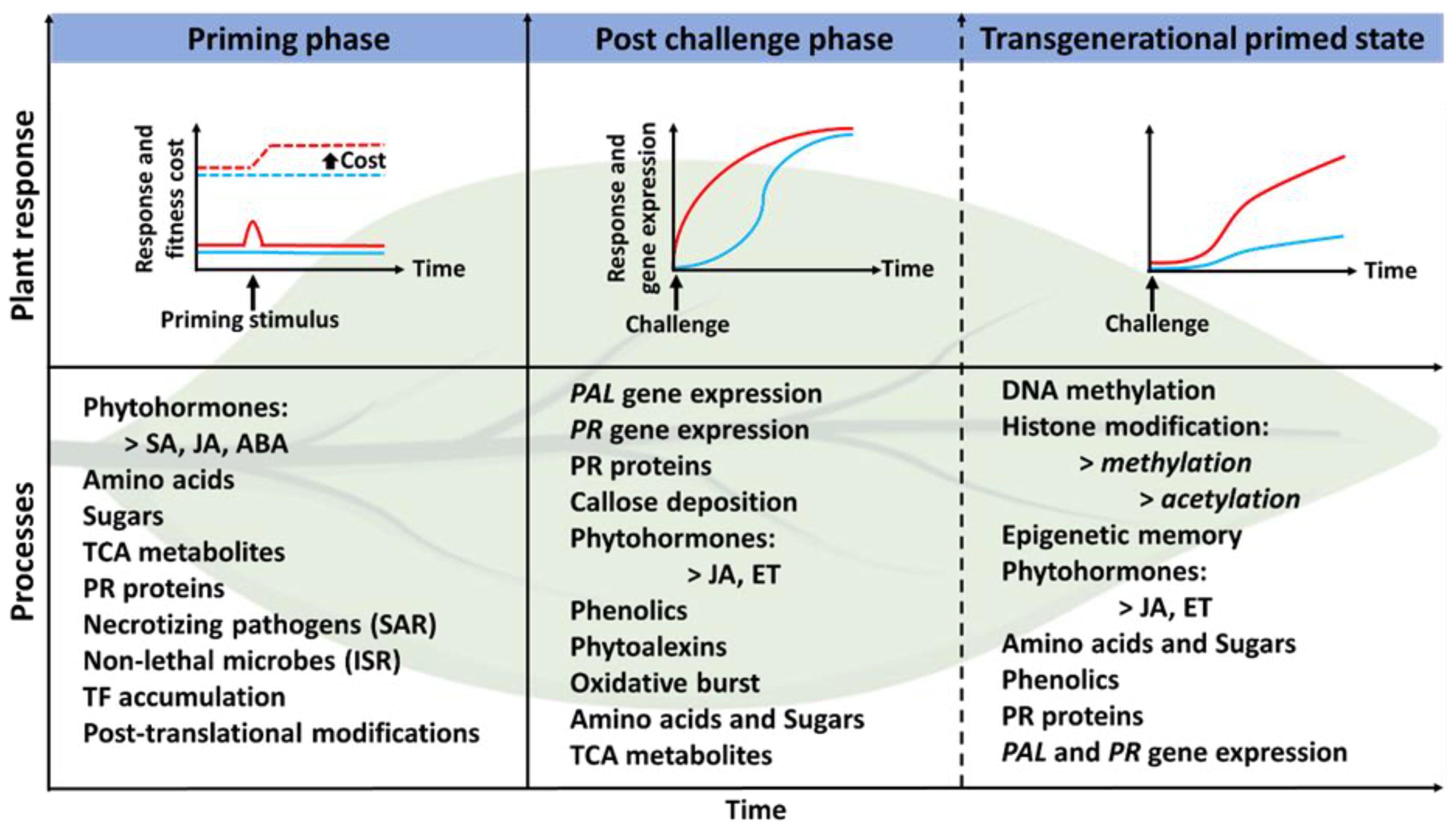
5. Induced Systemic Resistance and Plant Priming: PGPR-Mediated Plant Defence and Stress Tolerance from a Metabolomics Perspective
PGPR-Priming/Induced Systemic Resistance
6. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth-promoting rhizobacteria (PGPR): A review. Cogent. Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Etesami, H. Can interaction between silicon and plant growth-promoting rhizobacteria benefit in alleviating abiotic and biotic stresses in crop plants? Agric. Ecosyst. Environ. 2018, 253, 98–112. [Google Scholar] [CrossRef]
- Tsaniklidis, G.; Pappi, P.; Tsafouros, A.; Charova, S.N.; Nikoloudakis, N.; Roussos, P.A.; Paschalidis, K.A.; Delis, C. Polyamine homeostasis in tomato biotic/abiotic stress cross-tolerance. Gene 2020, 727, 144230. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Shao, H.; Qi, W.; Daly, P.; Sharma, A.; Shaghaleh, H.; Hamoud, Y.A.; El-Esawi, M.A.; Pan, R.; Wan, Q.; et al. Seed priming and foliar application with jasmonic acid enhance salinity stress tolerance of soybean (Glycine max L.) seedlings. J. Sci. Food Agric. 2020, 101, 2027–2041. [Google Scholar] [CrossRef]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef]
- Fróna, D.; Szenderák, J.; Harangi-Rákos, M. The challenge of feeding the world. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef] [Green Version]
- United Nations (UN). Population, Food Security, Nutrition and Sustainable Development; Economic and Social Council: New York, NY, USA, 2021. Available online: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/undesa_pd_2021_e_cn.9_2021_2_advanceunedited.pdf (accessed on 5 May 2021).
- Pessarakli, M.; Haghighi, M.; Sheibanirad, A. Plant responses under environmental stress conditions. Adv. Plants Agric. Res. 2015, 2, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Ahanger, M.A.; Akram, N.A.; Ashraf, M. Plant responses to environmental stresses–from gene to biotechnology. AoB Plants 2017, 9, plx025. [Google Scholar] [CrossRef] [Green Version]
- Gull, A.; Lone, A.A.; Wani, U.I. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stresses in Plants; de Oliveira, A.B., Ed.; IntechOpen: London, UK, 2019; pp. 1–6. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Burgess, M. World Human Population Problems. In Encyclopedia of the Anthropocene; Elsevier Inc.: Grand Rapids, MI, USA, 2018; pp. 313–317. [Google Scholar] [CrossRef]
- Tamburino, L.; Bravo, G.; Clough, Y.; Nicholas, K.A. From population to production: 50 years of scientific literature on how to feed the world. Glob. Food Sec. 2020, 24, 100346. [Google Scholar] [CrossRef]
- Tripathi, A.D.; Mishra, R.; Maurya, K.K.; Singh, R.B.; Wilson, D.W. Estimates for World Population and Global Food Availability for Global Health. In The Role of Functional Food Security in Global Health; Singh, B.R., Watson, R.R., Takahashi, T., Eds.; Academic Press: London, UK, 2019; Chapter 1; pp. 3–24. [Google Scholar] [CrossRef]
- Mishra, J.; Arora, N.K. Secondary metabolites of fluorescent pseudomonads in biocontrol of phytopathogens for sustainable agriculture. Appl. Soil Ecol. 2017, 125, 35–45. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 01741. [Google Scholar] [CrossRef] [PubMed]
- Adeniji, A.A.; Babalola, O.O.; Du Toit, L. Metabolomic applications for understanding complex tripartite plant-microbe interactions: Strategies and perspectives. Biotechnol. Rep. 2020, 25, e00425. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, J.; Noon, B.; Mohigmi, M.J.; Mazurie, A.; Dickensheets, D.L.; Bothner, B. Real-time digitization of metabolomics patterns from a living system using mass spectrometry. J. Am. Soc. Mass Spectrom. 2014, 25, 1755–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tugizimana, F.; Piater, L.A.; Dubery, I. Plant metabolomics: A new frontier in phytochemical analysis. S. Afr. J. Sci. 2013, 109, 1–11. [Google Scholar] [CrossRef]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Cold Spring Harb. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [Green Version]
- Bedair, M.; Glenn, K.C. Evaluation of the use of untargeted metabolomics in the safety assessment of genetically modified crops. Metabolomics 2020, 16, 111. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The chemistry of plant-microbe interactions in the rhizosphere and the potential for metabolomics to reveal signalling related to defence priming and induced systemic resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Tugizimana, F.; Mhlongo, M.I.; Piater, L.A.; Dubery, I.A. Metabolomics in plant priming research: The way forward? Int. J. Mol. Sci. 2018, 19, 1759. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.; Kang, S. Metabolomics, an essential tool in exploring and harnessing microbial chemical ecology. Phytobiomes J. 2020, 4, 195–210. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth-promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth-promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, Z. PGPR: Present role, mechanism of action and future prospects along bottlenecks in commercialization. J. Environ. Qual. 2021, 41, 9–15. [Google Scholar] [CrossRef]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems biology and multi-omics integration: Viewpoints from the metabolomics research community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, B.K.; Smith, M.; Baghela, A.; Lee, A.H.Y.; Hancock, R.E.W. Systems biology approaches to understanding the human immune system. Front. Immunol. 2020, 11, 1683. [Google Scholar] [CrossRef]
- Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Labuschagne, N.; Dubery, I.A. Unravelling the metabolic reconfiguration of the post-challenge primed state in Sorghum bicolor responding to Colletotrichum sublineolum infection. Metabolites 2019, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Hamany Djande, C.Y.; Pretorius, C.; Tugizimana, F.; Piater, L.A.; Dubery, I.A. Metabolomics: A tool for cultivar phenotyping and investigation of grain crops. Agronomy 2020, 10, 831. [Google Scholar] [CrossRef]
- Baharum, S.N.; Azizan, K.A. Omics Applications for Systems Bology. In Metabolomics in Systems Biology; Aziat, W.M., Goh, H., Baharum, S.N., Eds.; Springer Nature: Cham, Switzerland, 2018; Chapter 4; pp. 51–67. [Google Scholar] [CrossRef]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef]
- Kokova, D.; Mayboroda, O.A. Twenty years on: Metabolomics in helminth research. Trends Parasitol. 2019, 35, 282–288. [Google Scholar] [CrossRef]
- Burgess, K.; Rankin, N.; Weidt, S. Metabolomics. In Handbook of Pharmacogenomics and Stratified Medicine; Academic Press: Oxford, UK, 2014; Chapter 10; pp. 181–205. [Google Scholar] [CrossRef]
- Gowda, G.A.; Alvarado, L.Z.; Raftery, D. Metabolomics. In Nutrition in the Prevention and Treatment of Disease, 4th ed.; Academic Press: London, UK, 2017; Chapter 5; pp. 103–122. [Google Scholar] [CrossRef]
- Diray-Arce, J.; Conti, M.G.; Petrova, B.; Kanarek, N.; Angelidou, A.; Levy, O. Integrative metabolomics to identify molecular signatures of responses to vaccines and infections. Metabolites 2020, 10, 492. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, X.; Wang, C.; Zhang, H.; Cai, Z. Non-targeted and targeted metabolomics approaches to diagnosing lung cancer and predicting patient prognosis. Oncotarget 2016, 7, 63437–63448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilker, M.; Schwachtje, J.; Baier, M.; Balazadeh, S.; Bäurle, I.; Geiselhardt, S.; Hincha, D.K.; Kunze, R.; Mueller-Roeber, B.; Rillig, M.C.; et al. Priming and memory of stress responses in organisms lacking a nervous system. Biol. Rev. Biol. Proc. Camb. Philos. Soc. 2016, 91, 1118–1133. [Google Scholar] [CrossRef] [PubMed]
- Ramjegathesh, R.; Samiyappan, R.; Raguchander, T.; Prabakar, K.; Saravanakumar, D. Plant-PGPR Interactions for Pest and Disease Resistance in Sustainable Agriculture. In Bacteria in Agrobiology: Disease Management; Maheshwari, D.K., Ed.; Springer-Verlag: Berlin, Germany, 2013; Chapter 11; pp. 293–320. [Google Scholar] [CrossRef]
- Uzoh, I.M.; Babalola, O.O. Rhizosphere biodiversity as a premise for application in bio-economy. Agric. Ecosyst. Environ. 2018, 265, 524–534. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defence signalling. Microbiol. Res. 2020, 238, 126486. [Google Scholar] [CrossRef]
- Strehmel, N.; Böttcher, C.; Schmidt, S.; Scheel, D. Profiling of secondary metabolites in root exudates of Arabidopsis thaliana. Phytochemistry 2014, 108, 35–46. [Google Scholar] [CrossRef]
- Halder, S.; Sengupta, S. Microbial Ecology at Rhizosphere: Bioengineering and Future Prospective. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Choudhary, D.K., Ed.; Springer Nature: Singapore, 2016; Chapter 4; pp. 63–96. [Google Scholar] [CrossRef]
- Park, J.K.; Kang, B.R.; Ryu, C.; Anderson, A.J.; Kim, Y.C. Polyamine is a critical determinant of Pseudomonas chlororaphis O6 for GacS-dependent bacterial cell growth and biocontrol capacity. Mol. Plant Pathol. 2018, 19, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.; Patra, K.J. Revitalization of plant growth-promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Gray, E.J.; Smith, D.L. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant-bacterium signalling processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Kang, D.; Kirienko, D.R.; Webster, P.; Kirienko, N.V.; Fisher, A.L. Pyoverdine, a siderophore from Pseudomonas aeruginosa, translocate into C. elegans, removes iron and activates a distinct host response. Virulence 2018, 9, 804–817. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Turlings, T.C.J. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Cavaglieri, L.; Orlando, J.; Etcheverry, M. Rhizosphere microbial community structure at different maize plant growth stages and root locations. Microbiol. Res. 2009, 164, 391–399. [Google Scholar] [CrossRef]
- Dazzo, F.B.; Ganter, S. Rhizosphere. In Encyclopaedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: London, UK, 2009; pp. 335–349. [Google Scholar] [CrossRef]
- Grobelak, A.; Kokot, P.; Hutchison, D.; Grosser, A.; Kacprzak, M. Plant growth-promoting rhizobacteria as an alternative to mineral fertilizers in assisted bioremediation—Sustainable land and waste management. J. Environ. Manag. 2018, 227, 1–9. [Google Scholar] [CrossRef]
- Nabais, C.M.; Labuto, G.; Goncalves, S.; Buscardo, E.; Somensatto, D.; Nogueira, A.R.; Freitas, H. Effects of root age on the mineral allocation of metals, amino acids and sugars in different cell fractions of the perennial grass Paspalum notatum (bahiagrass). Plant Physiol. Biochem. 2011, 49, 1442–1447. [Google Scholar] [CrossRef]
- Mavrodi, O.V.; Mavrodi, D.V.; Parejko, J.A.; Thomashow, L.S.; Weller, D.M. Irrigation differentially impacts populations if indigenous antibiotic-producing Pseudomonas spp. in the rhizosphere of wheat. Appl. Environ. Microbiol. 2012, 78, 3214–3220. [Google Scholar] [CrossRef] [Green Version]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere engineering with plant growth-promoting microorganisms for agriculture and ecological sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.; Zhang, S. Induced Systemic Resistance and promotion of plant growth by Bacillus spp. Phytophathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, S.; Kabir, S.; Shabbir, U.; Batool, R. Plant growth-promoting rhizobacteria in sustainable agriculture: From theoretical to pragmatic approach. Symbiosis 2019, 78, 115–123. [Google Scholar] [CrossRef]
- Mhatre, P.H.; Karthik, C.; Kadirvelu, K.; Divya, K.L.; Venkatasalam, E.P.; Srinivasan, S.; Ramkumar, G.; Saranya, C.; Shanmuganathan, R. Plant growth-promoting rhizobacteria (PGPR): A potential alternative tool for nematodes bio-control. Biocatal. Agric. Biotechnol. 2019, 17, 119–128. [Google Scholar] [CrossRef]
- Agrawal, R.; Verma, A.; Satlewal, A. Bioprospecting PGPR Microflora by Novel Immune-Based Techniques. In New and Future Developments in Microbial Biotechnology and Bioengineering: Crop Improvement through Microbial Biotechnology; Prasad, R., Gill, S.S., Tuteja, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 24; pp. 465–478. [Google Scholar] [CrossRef]
- Lin, Y.; Watts, D.B.; Kloepper, W.J.; Adesemoye, A.O.; Feng, Y. Effect of Plant growth-promoting rhizobacteria at various nitrogen rates on corn growth. Agric. Sci. 2019, 10, 1542–1565. [Google Scholar] [CrossRef] [Green Version]
- Noumavo, P.A.; Kochoni, E.; Didagbé, Y.O.; Adjanohoun, A. Effect of different plant growth-promoting rhizobacteria on maize seed germination and seedling development. Am. J. Plant Sci. 2013, 4, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Asghar, H.N.; Arshad, M. Rhizobacteria capable of producing ACC-deaminase may mitigate salt stress in wheat. Soil Biol. Biochem. 2010, 74, 533–542. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Kumar, S.; Kumar, H.D.; Saini, J.K.; Agrawal, R.; Satlewal, A.; Ansari, M.W. Rhizosphere Metabolite Profiling: An Opportunity to Understand Plant-Microbe Interactions for Crop Improvement. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 17; pp. 343–361. [Google Scholar] [CrossRef]
- Pétriacq, P.; Williams, A.; Cotton, A.; McFarlane, A.E.; Rolfe, S.A.; Ton, J. Metabolite profiling of non-sterile rhizosphere soil. Plant J. 2017, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Smercina, D.N.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Impacts of nitrogen addition on switchgrass root-associated diazotrophic community structure and function. FEMS Microbiol. Ecol. 2020, 96, fiaa208. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.L.; Monsoori, N.; Da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loque, D.; Bowen, P.B.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, R.P.; Martyn, A.; Kopriva, S. Exometabolomic profiling of bacterial strains as cultivated using Arabidopsis root extract as the sole carbon source. Mol. Plant Microbe Interact. 2018, 31, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.B.; Heuberger, A.L.; Broeckling, C.D.; Jahn, C.E. Non-Targeted metabolomics reveals sorghum rhizosphere-associated exudates are influenced by the belowground interaction of substrate and sorghum genotype. Int. J. Mol. Sci. 2019, 20, 431. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Rey, T.; Schornack, S. Interactions of beneficial and detrimental root-colonizing filamentous microbes with plant hosts. Genome Biol. 2013, 14, 121. [Google Scholar] [CrossRef]
- Feussner, I.; Polle, A. What the transcriptome does not tell-proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef]
- Akram, W.; Aslam, H.; Ahmad, S.R.; Anjum, T.; Yasin, N.A.; Khan, W.U.; Ahmad, A.; Guo, J.; Wu, T.; Luo, W.; et al. Bacillus megaterium strain A12 ameliorates salinity stress in tomato plants through multiple mechanisms. J. Plant Interact. 2019, 14, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Smith, D.L. Plant Growth Promoting Rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Reuben, S.; Rai, A.; Pillai, B.V.S.; Rodrigues, A.; Swarup, S. A bacterial quercetin oxidoreductase QuoA-mediated perturbation in the phenylpropanoid metabolic network increases lignification with a concomitant decrease in phenolamides in Arabidopsis. J. Exp. Bot. 2013, 64, 5183–5194. [Google Scholar] [CrossRef]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an emerging tool for the study of plant-pathogen interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Labuschagne, N. Differential metabolic reprogramming in Paenibacillus alvei-primed Sorghum bicolor seedlings in response to Fusarium pseudograminearum infection. Metabolites 2019, 9, 150. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, V.K.; Tripathi, V.; Singh, P.P.; Singh, A.K. Plant Growth-Promoting Rhizobacteria (PGPR): Perspective in Agriculture under Biotic and Abiotic Stress. In New and Future Developments in Microbial Biotechnology and Bioengineering: Crop Improvement through Microbial Biotechnology; Prasad, R., Gill, S.S., Tuteja, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Chapter 16; pp. 333–342. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Rozier, C.; Hamzaoui, J.; Lemoine, G.D.; Czarnes, S.; Legendre, L. Field-based assessment mechanisms of maize yield enhancement by Azospirillum lipoferum CRT1. Sci. Rep. 2016, 7, 7416. [Google Scholar] [CrossRef] [Green Version]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic profiling of PGPR-treated tomato plants reveal priming-related adaptations of secondary metabolites and aromatic amino acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef]
- Schaker, P.; Peters, L.P.; Cataldi, T.R.; Labate, C.A.; Caldana, C.; Monteiro-Vitorello, C.B. Metabolome dynamics of smutted sugarcane reveals mechanisms involved in disease progression and whip emission. Front. Plant Sci. 2017, 8, 882. [Google Scholar] [CrossRef] [Green Version]
- Venturi, V.; Keel, C. Signalling in the rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef]
- Stokes, J.M.; Lopatkin, A.J.; Lobritz, M.A.; Collins, J.J. Bacterial metabolism and antibiotic efficacy. Cell Metab. 2019, 30, 251–259. [Google Scholar] [CrossRef]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma-pathogen-plant interactions, using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [Green Version]
- Marra, R.; Ambrosino, P.; Carbone, V.; Vinale, F.; Woo, S.L.; Ruocco, M.; Ciliento, R.; Lanzuise, S.; Ferraioli, S.; Soriente, I.; et al. Study of the three-way interaction between Trichoderma atroviride, plant and fungal pathogens by using a proteomic approach. Curr. Genet. 2006, 50, 307–321. [Google Scholar] [CrossRef]
- Allwood, J.W.; Clarke, A.; Goodacre, R.; Mur, L.A.J. Dual metabolomics: A novel approach to understanding plant-pathogen interactions. Phytochemistry 2010, 71, 590–597. [Google Scholar] [CrossRef]
- Olanrewaju, O.S. Isolation of Bacterial Strains for Improved Maize Production. Master’s Thesis, North-West University, Mahikeng, South Africa, 2016. Available online: https://dspace.nwu.ac.za/handle/10394/26090 (accessed on 23 July 2021).
- Saia, S.; Rappa, V.; Ruisi, P.; Abenavoli, M.R.; Sunseri, F.; Giambalvo, D.; Frenda, A.S.; Martinelli, F. Soil inoculation with symbiotic microorganisms promotes plant growth and nutrient transporter genes expression in durum wheat. Front. Plant Sci. 2015, 6, 815. [Google Scholar] [CrossRef] [Green Version]
- Creek, D.J.; Chokkanthukalam, A.; Jankevics, A.; Burgess, K.E.V.; Breitling, R.; Barrett, M.P. Stable isotope-assisted metabolomics for network-wide metabolic pathway elucidation. Anal. Chem. 2012, 84, 8442–8447. [Google Scholar] [CrossRef]
- Kleiner, M.; Dong, X.; Hinzke, T.; Wippler, J.; Thorson, E.; Mayer, B.; Strous, M. Metaproteomics method to determine carbon sources and assimilation pathways of species in microbial communities. Proc. Natl. Acad. Sci. USA 2018, 115, E5576–E5584. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Schwier, M.; Krumbach, J.; Kopriva, S.; Jacoby, P.R. Metabolomics in Plant-Microbe Interactions in the Roots. In Advances in Botanical Research; Pétriacq, P., Bouchereau, A., Eds.; Academic Press: London, UK, 2021; Chapter 5; Volume 98, pp. 133–161. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Mahe, S.; Ineson, P.; Staddon, P.; Ostle, N.; Cliquet, J.B.; Francez, A.; Fitter, A.H.; Young, J.P.W. Active root-inhabiting microbes identified by rapid incorporation of plant-derived carbon into RNA. Proc. Natl. Acad. Sci. USA 2007, 104, 16970–16975. [Google Scholar] [CrossRef] [Green Version]
- Pett-Ridge, J.; Firestone, M.K. Using stable isotopes to explore root-microbe-mineral interactions in soil. Rhizosphere 2017, 3, 244–253. [Google Scholar] [CrossRef]
- Nuccio, E.E.; Hodge, A.; Pett-Ridge, J.; Herman, D.J.; Weber, P.K.; Firestone, M.K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ. Microbiol. 2013, 15, 1870–1881. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Ryu, C.M. Chatting with a tiny belowground member of the holobiome: Communication between plants and growth-promoting rhizobacteria. Adv. Bot. Res. 2017, 82, 135–160. [Google Scholar]
- Willsey, T.; Chatterton, S.; Cárcamo, H. Interactions of root-feeding insects with fungal and oomycete plant pathogens. Front. Plant Sci. 2017, 8, 1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Wang, J.; Wang, W.; Tsui, C.K.; Cai, L. Changes in bacterial and fungal microbiomes associated with tomatoes of healthy and infected by Fusarium oxysporum f. sp. lycopersici. Microb. Ecol. 2020, 81, 1004–1017. [Google Scholar] [CrossRef]
- Maldonado-Gonzalez, M.M.; Bakker, P.A.H.M.; Prieto, P.; Marcado-Blanco, J. Arabidopsisthaliana as a tool to identify traits involved in Verticilliumdahliae biocontrol by the olive root endophyte Pseudomonas fluorescens PICF7. Front. Microbiol. 2015, 6, 266. [Google Scholar] [CrossRef]
- Miotto-Vilanova, L.; Jacquard, C.; Courteaux, B.; Wortham, L.; Michel, J.; Clement, C.; Barka, E.A.; Sanchez, L. Burkholderia phytofirmans PsJN confers grapevine resistance against Botrytis cinerea via a direct antimicrobial effect combined with a better resource mobilization. Front. Plant Sci. 2016, 7, 1236. [Google Scholar] [CrossRef] [Green Version]
- Westman, S.M.; Kloth, K.J.; Hanson, J.; Ohlsson, A.B.; Albrectsen, B.R. Defence priming in arabidopsis-a meta-analysis. Sci. Rep. 2019, 9, 13309. [Google Scholar] [CrossRef]
- Sharifi, R.; Ryu, C. Revisiting bacterial volatile-mediated plant growth promotion: Lessons from the past and objectives for the future. Ann. Bot. 2018, 122, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Mulla, S.I.; Lee, K.; Chae, J.; Shukla, P. VOCs-mediated hormonal signalling and crosstalk with plant growth-promoting microbes. Crit. Rev. Biotechnol. 2018, 38, 1277–1296. [Google Scholar] [CrossRef]
- Villena, J.; Kitazawa, H.; Van Wees, S.C.M.; Pieterse, C.M.J.; Takahashi, H. Receptors and signalling pathways for recognition of bacteria in livestock and crops: Prospects for beneficial microbes in healthy growth strategies. Front. Immunol. 2018, 9, 2223. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcantara, E.; Angulo, M.; Pérez-vicente, R. Induced Systemic Resistance (ISR) and Fe deficiency responses in dicot plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, R.F.; Geraats, B.P.J.; Kuramae, E.E.; Van Loon, L.C.; Bakker, P.A.M.H. Effects of jasmonic acid, ethylene and salicylic acid signalling on the rhizosphere bacterial community of Arabidopsis thaliana. Mol. Plant Microbe Interact. 2011, 4, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, M.J.; Van Der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smigielski, L.; Laubach, E.; Pesch, L.; Glock, J.M.L.; Albrecht, F.; Slusarenko, A.; Panstruga, R.; Kuhn, H. Nodulation induces systemic resistance of Medicago truncatula and Pisum sativum against Erysiphe pisi and primes for powdery mildew-triggered salicylic acid accumulation. Mol. Plant-Microbe Interact. 2019, 32, 1243–1255. [Google Scholar] [CrossRef]
- Zhu, F.; Xi, D.; Yuan, S.; Xu, F.; Zhang, D.; Lin, H. Salicylic acid and jasmonic acid are essential for systemic resistance against Tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant-Microbe Interact. 2014, 27, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Beckers, G.J.M.; Jaskiewicz, M.; Liu, Y.; Underwood, R.W.; He, S.Y.; Zhang, S.; Conrath, U. Mitogen-activated protein kinase 3 and 6 are required for full priming of stress response in Arab. thaliana. Plant Cell 2009, 21, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for enhanced defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Yip, E.C.; Tooker, J.F.; Mescher, M.C.; De Moraes, M.C. Costs of plant defense priming: Exposure to volatile cues from a specialist herbivore increases short-term growth but reduces rhizome production in tall goldenrod (Solidago altissima). BMC Plant Biol. 2019, 19, 209. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Rasmann, S.; De Vos, M.; Casteel, C.L.; Tian, D.; Halitschke, R.; Sun, J.Y.; Agrawal, A.A.; Felton, G.W.; Jander, G. Herbivory in the previous generation primes plants for enhanced insect resistance. Plant Physiol. 2012, 158, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; Pedrosa, F.O.; Muller-Santos, M.; de Souza, E.M. What did we learn from plant growth-promoting rhizobacteria (PGPR)-grass associations studies through proteomic and metabolomic approaches? Front. Sustain. Food Syst. 2020, 4, 607343. [Google Scholar] [CrossRef]
- Abd El-Daim, I.A.; Bejai, S.; Meijer, J. Bacillus velezensis 5113 induced metabolic and molecular reprogramming during abiotic stress tolerance in wheat. Sci. Rep. 2019, 9, 16282. [Google Scholar] [CrossRef] [Green Version]
- Fraire-Velázquez, S.; Balderas-Hernández, V.E. Abiotic Stress in Plants and Metabolic Responses. In Abiotic Stress—Plant Responses and Applications in Agriculture; Vahdati, K., Leslie, C., Eds.; InTech: Rijeka, Croatia, 2013; pp. 25–48. [Google Scholar] [CrossRef] [Green Version]
- Schwachtje, J.; Whitcomb, S.J.; Firmino, A.A.P.; Zuther, E.; Hincha, D.K.; Kopka, J. Induced, imprinted and primed responses to changing environments: Does metabolism store and process information? Front. Plant Sci. 2019, 10, 106. [Google Scholar] [CrossRef]
- Thellier, M.; Lüttge, U. Plant memory: A tentative model. Plant Biol. 2013, 15, 1–12. [Google Scholar] [CrossRef]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Hassen, A.I.; Labuschagne, N. Rhizobacteria-induced systemic resilience in Sorghum bicolor (L.) Moech against Fusarium pseudograminearum crown rot under drought stress conditions. Biol. Control. 2020, 151, 104395. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plant: Source Metabolome | PGPR (Treatment) Used | Significant Metabolite Perturbations | Reference |
|---|---|---|---|
| Soybean (Glycine max) | Bacillus simplex strain Sneb545 | Organic acids and amino acids | [48] |
| Tomato (Solanum lycopersicum) | Bacillus megaterium A12 (BMA12) | Sugars, amino acids, chlorophyll and carotenoids, antioxidants and phytohormones | [73] |
| Sorghum (Sorghum bicolor) | Paenibacillus alvei strain T22 | Amino acids, lipids, flavonoids, phytohormones | [77] |
| Tomato (Solanum lycopersicum) | Bacillus megaterium A12 (BMA12) | Phytohormones | [78] |
| Chickpea (Cicer arietinum) | Bacillus subtilis, Bacillus thuringiensis and Bacillus megaterium | Salicylate, tryptophan, saccharic acid, glyceric acid, aminophenol, 5-oxo-L-proline, L-carnitine, trans-cinnamate, succinate and syringic acid | [79] |
| Maise (Zea mays) | Azospirillum lipoferum CRT1 | Glucose, lactic acid, acidic intermediates of the pentose phosphate and ascorbate/aldarate pathways and defence-related hydroxycinnamic acids | [80] |
| Tomato (Solanum lycopersicum, cv. Moneymaker) | Pseudomonas fluorescens N04, P. koreensis N19, Paenibacillus alvei T22 and Lysinibacillus sphaericus T19 | Hydroxycinnamic acid derivatives, benzoates, flavonoids, glycoalkaloids, fatty acids, amino acids and phytohormones | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashabela, M.D.; Piater, L.A.; Dubery, I.A.; Tugizimana, F.; Mhlongo, M.I. Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review. Biology 2022, 11, 346. https://doi.org/10.3390/biology11030346
Mashabela MD, Piater LA, Dubery IA, Tugizimana F, Mhlongo MI. Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review. Biology. 2022; 11(3):346. https://doi.org/10.3390/biology11030346
Chicago/Turabian StyleMashabela, Manamele D., Lizelle A. Piater, Ian A. Dubery, Fidele Tugizimana, and Msizi I. Mhlongo. 2022. "Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review" Biology 11, no. 3: 346. https://doi.org/10.3390/biology11030346
APA StyleMashabela, M. D., Piater, L. A., Dubery, I. A., Tugizimana, F., & Mhlongo, M. I. (2022). Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review. Biology, 11(3), 346. https://doi.org/10.3390/biology11030346