
Neuraminidases—Key Players in the Inflammatory Response after Pathophysiological Cardiac Stress and Potential New Therapeutic Targets in Cardiac Disease


Abstract
:Simple Summary
Abstract
1. Introduction
2. Mammalian Neuraminidases
3. Functions and Implications of Neuraminidases
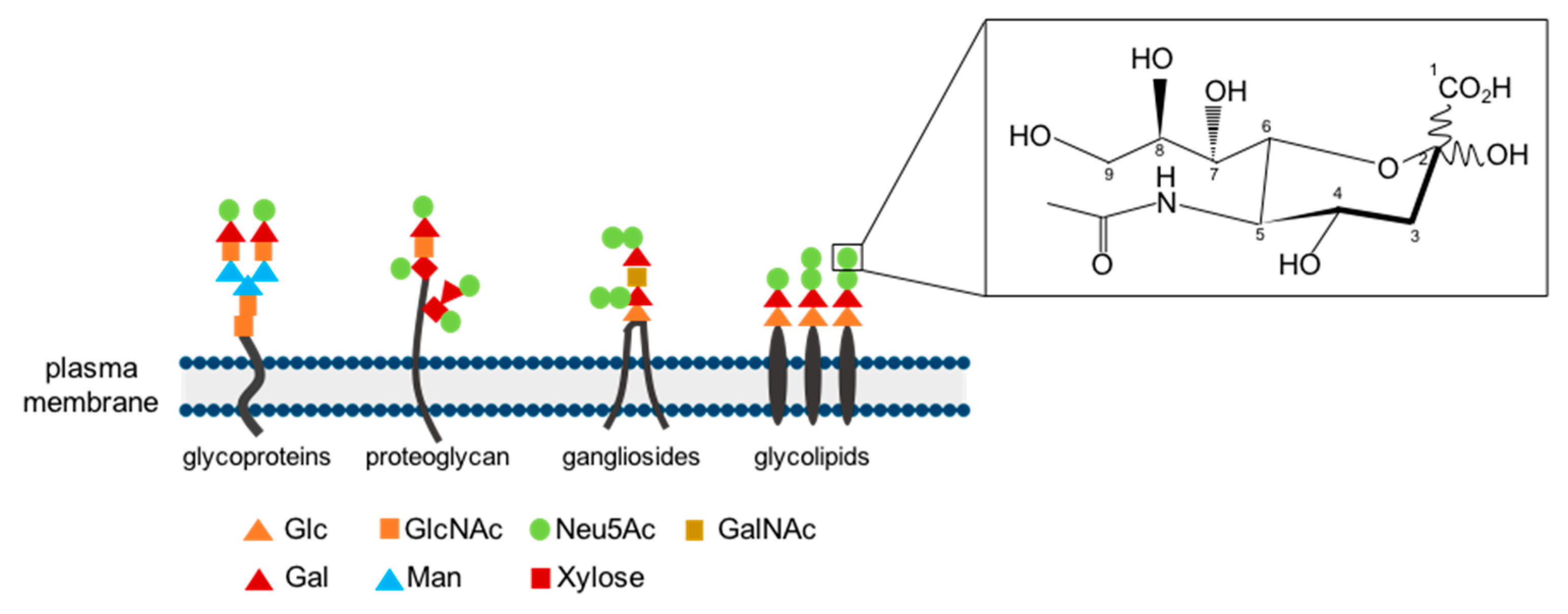
4. Occurrence and Substrate Specificity of Neuraminidases
5. Neuraminidases and Their Role in the Immune System
6. Neuraminidases in the Context of Cardiac Pathologies
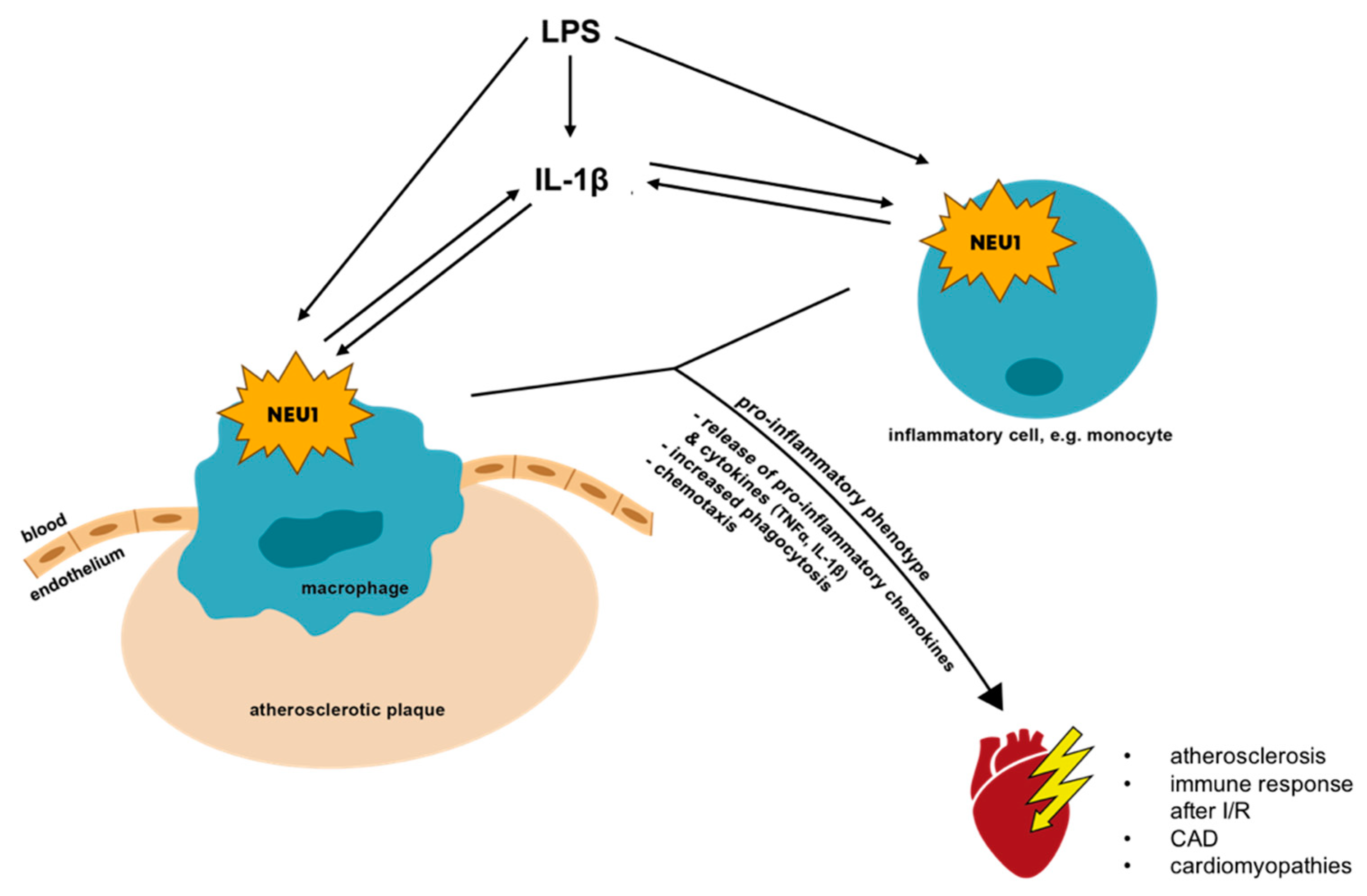
6.1. Inflammatory Vascular Diseases: Atherosclerosis, Coronary Artery Disease and Ischemia/Reperfusion Injury
6.1.1. Atherosclerosis and Coronary Artery Disease
6.1.2. Ischemia/Reperfusion Injuries
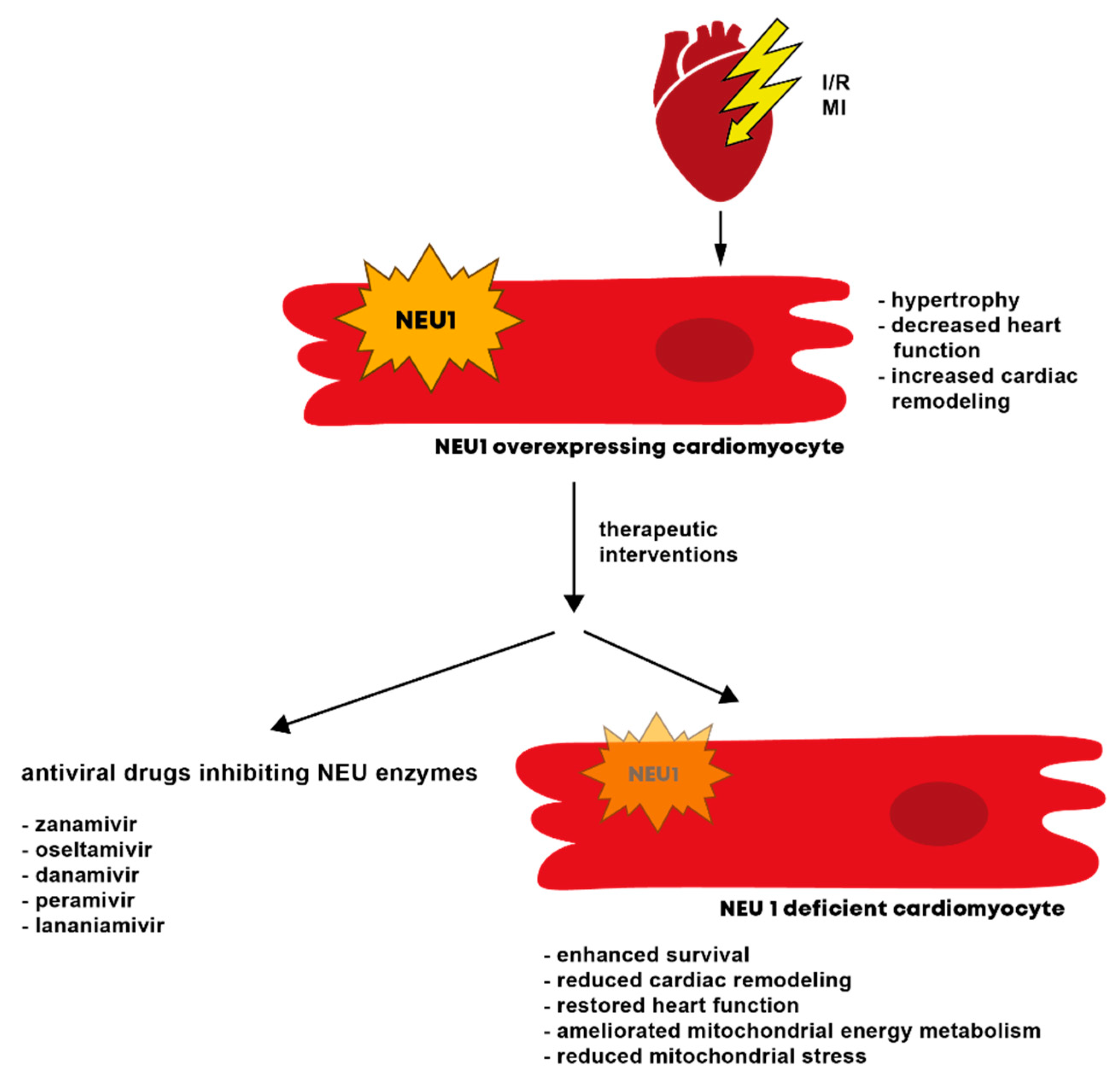
6.2. Cardiomyopathy and Heart Failure
7. Neuraminidase Activity and the Resulting Sialic Acid Level as a Diagnostic Biomarker for Cardiovascular Diseases and concomitant Inflammation
8. The Potential of Neuraminidases as Therapeutic Targets
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varki, A.; Gagneux, P. Multifarious roles of sialic acids in immunity. Ann. N. Y. Acad. Sci. 2012, 1253, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, G.; Guan, F. Biological Functions and Analytical Strategies of Sialic Acids in Tumor. Cells 2020, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Poznyak, A.V.; Zhang, D.; Grechko, A.V.; Wu, W.K.; Orekhov, A.N. The role of sialic acids in the initiation of atherosclerosis. Minerva Cardioangiol. 2020, 68, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, P.; Kaufmann, T. Sialic Acids and Their Influence on Human NK Cell Function. Cells 2021, 10, 263. [Google Scholar] [CrossRef]
- Lubbers, J.; Rodriguez, E.; van Kooyk, Y. Modulation of Immune Tolerance via Siglec-Sialic Acid Interactions. Front. Immunol. 2018, 9, 2807. [Google Scholar] [CrossRef]
- Pshezhetsky, A.V.; Ashmarina, L.I. Desialylation of surface receptors as a new dimension in cell signaling. Biochemistry 2013, 78, 736–745. [Google Scholar] [CrossRef]
- Masola, V.; Zaza, G.; Arduini, A.; Onisto, M.; Gambaro, G. Endothelial Glycocalyx as a Regulator of Fibrotic Processes. Int. J. Mol. Sci. 2021, 22, 2996. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Nieddu, G.; Piperigkou, Z.; Kyriakopoulou, K.; Karamanos, N.; Formato, M. Circulating Heparan Sulfate Proteoglycans as Biomarkers in Health and Disease. Semin Thromb. Hemost 2021, 47, 295–307. [Google Scholar] [CrossRef]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef]
- Pries, A.R.; Secomb, T.W.; Gaehtgens, P. The endothelial surface layer. Pflugers Arch. 2000, 440, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Sieve, I.; Munster-Kuhnel, A.K.; Hilfiker-Kleiner, D. Regulation and function of endothelial glycocalyx layer in vascular diseases. Vasc. Pharmacol. 2018, 100, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Potje, S.R.; Paula, T.D.; Paulo, M.; Bendhack, L.M. The Role of Glycocalyx and Caveolae in Vascular Homeostasis and Diseases. Front. Physiol. 2020, 11, 620840. [Google Scholar] [CrossRef] [PubMed]
- Urschel, K.; Tauchi, M.; Achenbach, S.; Dietel, B. Investigation of Wall Shear Stress in Cardiovascular Research and in Clinical Practice-From Bench to Bedside. Int. J. Mol. Sci. 2021, 22, 5635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Chen, Q.Q.; Li, J.; Zhang, L.; Qi, L.W. Neuraminidase 1 and its Inhibitors from Chinese Herbal Medicines: An Emerging Role for Cardiovascular Diseases. Am. J. Chin. Med. 2021, 49, 843–862. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.; La Rocca, P.; Allevi, P.; Pappone, C.; Anastasia, L. Intramolecular Lactones of Sialic Acids. Int. J. Mol. Sci. 2020, 21, 8098. [Google Scholar] [CrossRef]
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Curr. Opin. Struct. Biol. 2009, 19, 507–514. [Google Scholar] [CrossRef]
- Bourguet, E.; Figurska, S.; Fra Czek, M.M. Human Neuraminidases: Structures and Stereoselective Inhibitors. J. Med. Chem. 2022, 65, 3002–3025. [Google Scholar] [CrossRef]
- Kim, Y.J.; Varki, A. Perspectives on the significance of altered glycosylation of glycoproteins in cancer. Glycoconj. J. 1997, 14, 569–576. [Google Scholar] [CrossRef]
- Dennis, J.W.; Granovsky, M.; Warren, C.E. Protein glycosylation in development and disease. Bioessays 1999, 21, 412–421. [Google Scholar] [CrossRef]
- Krzeslak, A.; Gaj, Z.; Pomorski, L.; Lipinska, A. Sialylation of intracellular proteins of thyroid lesions. Oncol. Rep. 2007, 17, 1237–1242. [Google Scholar] [CrossRef]
- Glanz, V.Y.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Sialidase activity in human pathologies. Eur. J. Pharmacol. 2019, 842, 345–350. [Google Scholar] [CrossRef]
- Seyrantepe, V.; Poupetova, H.; Froissart, R.; Zabot, M.T.; Maire, I.; Pshezhetsky, A.V. Molecular pathology of NEU1 gene in sialidosis. Hum. Mutat. 2003, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef]
- Womack, J.E.; Yan, D.L.; Potier, M. Gene for neuraminidase activity on mouse chromosome 17 near h-2: Pleiotropic effects on multiple hydrolases. Science 1981, 212, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Kijimoto-Ochiai, S.; Koda, T.; Suwama, T.; Matsukawa, H.; Fujii, M.; Tomobe, K.; Nishimura, M. Low expression of Neu2 sialidase in the thymus of SM/J mice-existence of neuraminidase positive cells “Neu-medullocyte” in the murine thymus. Glycoconj. J. 2008, 25, 787–796. [Google Scholar] [CrossRef]
- Heimerl, M.; Sieve, I.; Ricke-Hoch, M.; Erschow, S.; Battmer, K.; Scherr, M.; Hilfiker-Kleiner, D. Neuraminidase-1 promotes heart failure after ischemia/reperfusion injury by affecting cardiomyocytes and invading monocytes/macrophages. Basic Res. Cardiol. 2020, 115, 62. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Seyrantepe, V.; Landry, K.; Ahmad, R.; Ahmad, A.; Stamatos, N.M.; Pshezhetsky, A.V. Monocyte differentiation up-regulates the expression of the lysosomal sialidase, Neu1, and triggers its targeting to the plasma membrane via major histocompatibility complex class II-positive compartments. J. Biol. Chem. 2006, 281, 27526–27538. [Google Scholar] [CrossRef]
- Howlader, M.A.; Li, C.; Zou, C.; Chakraberty, R.; Ebesoh, N.; Cairo, C.W. Neuraminidase-3 Is a Negative Regulator of LFA-1 Adhesion. Front. Chem. 2019, 7, 791. [Google Scholar] [CrossRef]
- Timur, Z.K.; Inci, O.K.; Demir, S.A.; Seyrantepe, V. Sialidase neu4 deficiency is associated with neuroinflammation in mice. Glycoconj. J. 2021, 38, 649–667. [Google Scholar] [CrossRef]
- Monti, E.; Bassi, M.T.; Bresciani, R.; Civini, S.; Croci, G.L.; Papini, N.; Riboni, M.; Zanchetti, G.; Ballabio, A.; Preti, A.; et al. Molecular cloning and characterization of NEU4, the fourth member of the human sialidase gene family. Genomics 2004, 83, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Sieve, I.; Ricke-Hoch, M.; Kasten, M.; Battmer, K.; Stapel, B.; Falk, C.S.; Leisegang, M.S.; Haverich, A.; Scherr, M.; Hilfiker-Kleiner, D. A positive feedback loop between IL-1beta, LPS and NEU1 may promote atherosclerosis by enhancing a pro-inflammatory state in monocytes and macrophages. Vasc. Pharmacol. 2018, 103–105, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.Q.; Ma, G.; Liu, J.F.; Cai, Y.Y.; Zhang, J.Y.; Wei, T.T.; Pan, A.; Jiang, S.; Xiao, Y.; Xiao, P.; et al. Neuraminidase 1 is a driver of experimental cardiac hypertrophy. Eur. Heart J. 2021, 42, 3770–3782. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Ha, D.I.; Son, C.; Kang, J.G.; Hwang, H.; Moon, S.B.; Kim, M.; Nam, J.; Kim, J.S.; Song, S.Y.; et al. Defect in cytosolic Neu2 sialidase abrogates lipid metabolism and impairs muscle function in vivo. Sci. Rep. 2022, 12, 3216. [Google Scholar] [CrossRef] [PubMed]
- Tringali, C.; Papini, N.; Fusi, P.; Croci, G.; Borsani, G.; Preti, A.; Tortora, P.; Tettamanti, G.; Venerando, B.; Monti, E. Properties of recombinant human cytosolic sialidase HsNEU2. The enzyme hydrolyzes monomerically dispersed GM1 ganglioside molecules. J. Biol. Chem. 2004, 279, 3169–3179. [Google Scholar] [CrossRef]
- Chen, S.; Li, M.; Jiang, W.; Zheng, H.; Qi, L.W.; Jiang, S. The role of Neu1 in the protective effect of dipsacoside B on acetaminophen-induced liver injury. Ann. Transl. Med. 2020, 8, 823. [Google Scholar] [CrossRef]
- Annunziata, I.; Patterson, A.; Helton, D.; Hu, H.; Moshiach, S.; Gomero, E.; Nixon, R.; d’Azzo, A. Lysosomal NEU1 deficiency affects amyloid precursor protein levels and amyloid-beta secretion via deregulated lysosomal exocytosis. Nat. Commun. 2013, 4, 2734. [Google Scholar] [CrossRef]
- Fanzani, A.; Giuliani, R.; Colombo, F.; Zizioli, D.; Presta, M.; Preti, A.; Marchesini, S. Overexpression of cytosolic sialidase Neu2 induces myoblast differentiation in C2C12 cells. FEBS Lett. 2003, 547, 183–188. [Google Scholar] [CrossRef]
- Fanzani, A.; Colombo, F.; Giuliani, R.; Preti, A.; Marchesini, S. Insulin-like growth factor 1 signaling regulates cytosolic sialidase Neu2 expression during myoblast differentiation and hypertrophy. FEBS J. 2006, 273, 3709–3721. [Google Scholar] [CrossRef]
- Fanzani, A.; Colombo, F.; Giuliani, R.; Preti, A.; Marchesini, S. Cytosolic sialidase Neu2 upregulation during PC12 cells differentiation. FEBS Lett. 2004, 566, 178–182. [Google Scholar] [CrossRef]
- Valaperta, R.; Valsecchi, M.; Rocchetta, F.; Aureli, M.; Prioni, S.; Prinetti, A.; Chigorno, V.; Sonnino, S. Induction of axonal differentiation by silencing plasma membrane-associated sialidase Neu3 in neuroblastoma cells. J. Neurochem. 2007, 100, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Proshin, S.; Yamaguchi, K.; Yamashita, Y.; Katakura, R.; Yamamoto, K.; Shima, H.; Hosono, M.; Miyagi, T. Sialidase NEU3 defines invasive potential of human glioblastoma cells by regulating calpain-mediated proteolysis of focal adhesion proteins. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2778–2788. [Google Scholar] [CrossRef]
- Scaringi, R.; Piccoli, M.; Papini, N.; Cirillo, F.; Conforti, E.; Bergante, S.; Tringali, C.; Garatti, A.; Gelfi, C.; Venerando, B.; et al. NEU3 sialidase is activated under hypoxia and protects skeletal muscle cells from apoptosis through the activation of the epidermal growth factor receptor signaling pathway and the hypoxia-inducible factor (HIF)-1alpha. J. Biol. Chem. 2013, 288, 3153–3162. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Hata, K.; Koseki, K.; Shiozaki, K.; Akita, H.; Wada, T.; Moriya, S.; Miyagi, T. Evidence for mitochondrial localization of a novel human sialidase (NEU4). Biochem. J. 2005, 390, 85–93. [Google Scholar] [CrossRef]
- Shiozaki, K.; Koseki, K.; Yamaguchi, K.; Shiozaki, M.; Narimatsu, H.; Miyagi, T. Developmental change of sialidase neu4 expression in murine brain and its involvement in the regulation of neuronal cell differentiation. J. Biol. Chem. 2009, 284, 21157–21164. [Google Scholar] [CrossRef] [PubMed]
- Bonten, E.J.; Campos, Y.; Zaitsev, V.; Nourse, A.; Waddell, B.; Lewis, W.; Taylor, G.; d’Azzo, A. Heterodimerization of the sialidase NEU1 with the chaperone protective protein/cathepsin A prevents its premature oligomerization. J. Biol. Chem. 2009, 284, 28430–28441. [Google Scholar] [CrossRef]
- Luu, A.R.; Wong, C.; Agrawal, V.; Wise, N.; Handyside, B.; Lo, M.J.; Pacheco, G.; Felix, J.B.; Giaramita, A.; d’Azzo, A.; et al. Intermittent enzyme replacement therapy with recombinant human beta-galactosidase prevents neuraminidase 1 deficiency. J. Biol. Chem. 2020, 295, 13556–13569. [Google Scholar] [CrossRef]
- Van Diggelen, O.P.; Schram, A.W.; Sinnott, M.L.; Smith, P.J.; Robinson, D.; Galjaard, H. Turnover of beta-galactosidase in fibroblasts from patients with genetically different types of beta-galactosidase deficiency. Biochem. J. 1981, 200, 143–151. [Google Scholar] [CrossRef]
- van der Spoel, A.; Bonten, E.; d’Azzo, A. Transport of human lysosomal neuraminidase to mature lysosomes requires protective protein/cathepsin A. EMBO J. 1998, 17, 1588–1597. [Google Scholar] [CrossRef]
- Carrillo, M.B.; Milner, C.M.; Ball, S.T.; Snoek, M.; Campbell, R.D. Cloning and characterization of a sialidase from the murine histocompatibility-2 complex: Low levels of mRNA and a single amino acid mutation are responsible for reduced sialidase activity in mice carrying the Neu1a allele. Glycobiology 1997, 7, 975–986. [Google Scholar] [CrossRef]
- Milner, C.M.; Smith, S.V.; Carrillo, M.B.; Taylor, G.L.; Hollinshead, M.; Campbell, R.D. Identification of a sialidase encoded in the human major histocompatibility complex. J. Biol. Chem. 1997, 272, 4549–4558. [Google Scholar] [CrossRef] [PubMed]
- Bonten, E.; van der Spoel, A.; Fornerod, M.; Grosveld, G.; d’Azzo, A. Characterization of human lysosomal neuraminidase defines the molecular basis of the metabolic storage disorder sialidosis. Genes Dev. 1996, 10, 3156–3169. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Kim, A.R.; Lee, C.; Kim, N.K.D.; Kim, N.S.; Park, W.Y.; Kim, M.; Youn, J.; Cho, J.W.; Kim, J.S. Type 1 Sialidosis Patient With a Novel Deletion Mutation in the NEU1 Gene: Case Report and Literature Review. Cerebellum 2019, 18, 659–664. [Google Scholar] [CrossRef]
- Khan, A.; Das, S.; Sergi, C. Therapeutic Potential of Neu1 in Alzheimer’s Disease Via the Immune System. Am. J. Alzheimers Dis. Other Demen. 2021, 36, 1533317521996147. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Schauer, R. Sialic Acids. In Essentials of Glycobiology, 2nd ed.; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor (NY): New York, NY, USA, 2009. [Google Scholar]
- Wang, L.; Seino, J.; Tomotake, H.; Funakoshi, Y.; Hirayama, H.; Suzuki, T. Co-Expression of NEU2 and GBA3 Causes a Drastic Reduction in Cytosolic Sialyl Free N-glycans in Human MKN45 Stomach Cancer Cells-Evidence for the Physical Interaction of NEU2 and GBA3. Biomolecules 2015, 5, 1499–1514. [Google Scholar] [CrossRef]
- Breiden, B.; Sandhoff, K. Ganglioside Metabolism and Its Inherited Diseases. Methods Mol. Biol. 2018, 1804, 97–141. [Google Scholar] [CrossRef]
- Smutova, V.; Albohy, A.; Pan, X.; Korchagina, E.; Miyagi, T.; Bovin, N.; Cairo, C.W.; Pshezhetsky, A.V. Structural basis for substrate specificity of mammalian neuraminidases. PLoS ONE 2014, 9, e106320. [Google Scholar] [CrossRef]
- Bigi, A.; Morosi, L.; Pozzi, C.; Forcella, M.; Tettamanti, G.; Venerando, B.; Monti, E.; Fusi, P. Human sialidase NEU4 long and short are extrinsic proteins bound to outer mitochondrial membrane and the endoplasmic reticulum, respectively. Glycobiology 2010, 20, 148–157. [Google Scholar] [CrossRef]
- Bigi, A.; Tringali, C.; Forcella, M.; Mozzi, A.; Venerando, B.; Monti, E.; Fusi, P. A proline-rich loop mediates specific functions of human sialidase NEU4 in SK-N-BE neuronal differentiation. Glycobiology 2013, 23, 1499–1509. [Google Scholar] [CrossRef]
- De Maria, R.; Lenti, L.; Malisan, F.; d’Agostino, F.; Tomassini, B.; Zeuner, A.; Rippo, M.R.; Testi, R. Requirement for GD3 ganglioside in CD95- and ceramide-induced apoptosis. Science 1997, 277, 1652–1655. [Google Scholar] [CrossRef]
- Comelli, E.M.; Amado, M.; Lustig, S.R.; Paulson, J.C. Identification and expression of Neu4, a novel murine sialidase. Gene 2003, 321, 155–161. [Google Scholar] [CrossRef]
- Schauer, R.; Kamerling, J.P. Exploration of the Sialic Acid World. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Varki, A. The sialome-far more than the sum of its parts. OMICS 2010, 14, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, F.; Tiralongo, E.; Tiralongo, J. Sialic acid-specific lectins: Occurrence, specificity and function. Cell Mol. Life Sci. 2006, 63, 1331–1354. [Google Scholar] [CrossRef]
- Klaas, M.; Oetke, C.; Lewis, L.E.; Erwig, L.P.; Heikema, A.P.; Easton, A.; Willison, H.J.; Crocker, P.R. Sialoadhesin promotes rapid proinflammatory and type I IFN responses to a sialylated pathogen, Campylobacter jejuni. J. Immunol. 2012, 189, 2414–2422. [Google Scholar] [CrossRef]
- Chang, Y.C.; Olson, J.; Louie, A.; Crocker, P.R.; Varki, A.; Nizet, V. Role of macrophage sialoadhesin in host defense against the sialylated pathogen group B Streptococcus. J. Mol. Med. 2014, 92, 951–959. [Google Scholar] [CrossRef]
- Natori, Y.; Nasui, M.; Edo, K.; Sato, S.; Sakurai, T.; Kizaki, T.; Kihara-Negishi, F. NEU1 sialidase controls gene expression and secretion of IL-6 and MCP-1 through NF-kappaB pathway in 3T3-L1 adipocytes. J. Biochem. 2017, 162, 137–143. [Google Scholar] [CrossRef]
- Karmakar, J.; Roy, S.; Mandal, C. Modulation of TLR4 Sialylation Mediated by a Sialidase Neu1 and Impairment of Its Signaling in Leishmania donovani Infected Macrophages. Front. Immunol. 2019, 10, 2360. [Google Scholar] [CrossRef]
- Kawecki, C.; Bocquet, O.; Schmelzer, C.E.H.; Heinz, A.; Ihling, C.; Wahart, A.; Romier, B.; Bennasroune, A.; Blaise, S.; Terryn, C.; et al. Identification of CD36 as a new interaction partner of membrane NEU1: Potential implication in the pro-atherogenic effects of the elastin receptor complex. Cell Mol. Life Sci. 2019, 76, 791–807. [Google Scholar] [CrossRef]
- Pshezhetsky, A.V.; Hinek, A. Where catabolism meets signalling: Neuraminidase 1 as a modulator of cell receptors. Glycoconj. J. 2011, 28, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhalek, S.; Szewczuk, M.R. Neu1 sialidase and matrix metalloproteinase-9 cross-talk regulates nucleic acid-induced endosomal TOLL-like receptor-7 and -9 activation, cellular signaling and pro-inflammatory responses. Cell Signal 2013, 25, 2093–2105. [Google Scholar] [CrossRef] [PubMed]
- Seyrantepe, V.; Iannello, A.; Liang, F.; Kanshin, E.; Jayanth, P.; Samarani, S.; Szewczuk, M.R.; Ahmad, A.; Pshezhetsky, A.V. Regulation of phagocytosis in macrophages by neuraminidase 1. J. Biol. Chem. 2010, 285, 206–215. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.; Peng, X.; Zheng, Z.; Le, A.; Guo, J.; Ma, L.; Shi, H.; Yao, K.; Zhang, S.; et al. Neuraminidase 1 Exacerbating Aortic Dissection by Governing a Pro-Inflammatory Program in Macrophages. Front. Cardiovasc. Med. 2021, 8, 788645. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhalek, S.; Amith, S.R.; Franchuk, S.L.; Jayanth, P.; Guo, M.; Finlay, T.; Gilmour, A.; Guzzo, C.; Gee, K.; Beyaert, R.; et al. Neu1 sialidase and matrix metalloproteinase-9 cross-talk is essential for Toll-like receptor activation and cellular signaling. J. Biol. Chem. 2011, 286, 36532–36549. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, J.; Mandal, C. Interplay Between Sialic Acids, Siglec-E, and Neu1 Regulates MyD88- and TRIF-Dependent Pathways for TLR4-Activation During Leishmania donovani Infection. Front. Immunol. 2021, 12, 626110. [Google Scholar] [CrossRef]
- Allendorf, D.H.; Franssen, E.H.; Brown, G.C. Lipopolysaccharide activates microglia via neuraminidase 1 desialylation of Toll-like Receptor 4. J. Neurochem. 2020, 155, 403–416. [Google Scholar] [CrossRef]
- Abdulkhalek, S.; Guo, M.; Amith, S.R.; Jayanth, P.; Szewczuk, M.R. G-protein coupled receptor agonists mediate Neu1 sialidase and matrix metalloproteinase-9 cross-talk to induce transactivation of TOLL-like receptors and cellular signaling. Cell Signal 2012, 24, 2035–2042. [Google Scholar] [CrossRef]
- Katoh, S.; Kaminuma, O.; Hiroi, T.; Mori, A.; Ohtomo, T.; Maeda, S.; Shimizu, H.; Obase, Y.; Oka, M. CD44 is critical for airway accumulation of antigen-specific Th2, but not Th1, cells induced by antigen challenge in mice. Eur. J. Immunol. 2011, 41, 3198–3207. [Google Scholar] [CrossRef]
- Katoh, S.; Maeda, S.; Fukuoka, H.; Wada, T.; Moriya, S.; Mori, A.; Yamaguchi, K.; Senda, S.; Miyagi, T. A crucial role of sialidase Neu1 in hyaluronan receptor function of CD44 in T helper type 2-mediated airway inflammation of murine acute asthmatic model. Clin. Exp. Immunol. 2010, 161, 233–241. [Google Scholar] [CrossRef]
- Howlader, M.A.; Demina, E.P.; Samarani, S.; Guo, T.; Caillon, A.; Ahmad, A.; Pshezhetsky, A.V.; Cairo, C.W. The Janus-like role of neuraminidase isoenzymes in inflammation. FASEB J. 2022, 36, e22285. [Google Scholar] [CrossRef]
- Howlader, M.A.; Guo, T.; Cairo, C.W. Inhibitors of Human Neuraminidase Enzymes Block Transmigration in vitro. Front. Mol. Biosci. 2022, 9, 835757. [Google Scholar] [CrossRef] [PubMed]
- Stamatos, N.M.; Carubelli, I.; van de Vlekkert, D.; Bonten, E.J.; Papini, N.; Feng, C.; Venerando, B.; d’Azzo, A.; Cross, A.S.; Wang, L.X.; et al. LPS-induced cytokine production in human dendritic cells is regulated by sialidase activity. J. Leukoc Biol. 2010, 88, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Nan, X.; Carubelli, I.; Stamatos, N.M. Sialidase expression in activated human T lymphocytes influences production of IFN-gamma. J. Leukoc Biol. 2007, 81, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, J.; Bian, H.; Wu, P.; Kuvelkar, R.; Kung, T.T.; Crawley, Y.; Egan, R.W.; Billah, M.M. Induction of lysosomal and plasma membrane-bound sialidases in human T-cells via T-cell receptor. Biochem. J. 2004, 380, 425–433. [Google Scholar] [CrossRef]
- Shkandina, T.; Herrmann, M.; Bilyy, R. Sweet kiss of dying cell: Sialidase activity on apoptotic cell is able to act toward its neighbors. Autoimmunity 2012, 45, 574–578. [Google Scholar] [CrossRef]
- Bilyy, R.O.; Shkandina, T.; Tomin, A.; Munoz, L.E.; Franz, S.; Antonyuk, V.; Kit, Y.Y.; Zirngibl, M.; Furnrohr, B.G.; Janko, C.; et al. Macrophages discriminate glycosylation patterns of apoptotic cell-derived microparticles. J. Biol. Chem. 2012, 287, 496–503. [Google Scholar] [CrossRef]
- Wu, C.Y.; Chuang, H.Y.; Wong, C.H. Influenza virus neuraminidase regulates host CD8(+) T-cell response in mice. Commun. Biol. 2020, 3, 748. [Google Scholar] [CrossRef]
- Lee, W.C.; Lee, W.L.; Shyong, W.Y.; Yang, L.W.; Ko, M.C.; Sheu, B.C.; Hsieh, S.L.; Wang, P.H. Increased concentration of sialidases by HeLa cells might influence the cytotoxic ability of NK cells. Taiwan J. Obstet. Gynecol. 2012, 51, 192–198. [Google Scholar] [CrossRef]
- Jebari-Benslaiman, S.; Galicia-Garcia, U.; Larrea-Sebal, A.; Olaetxea, J.R.; Alloza, I.; Vandenbroeck, K.; Benito-Vicente, A.; Martin, C. Pathophysiology of Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 3346. [Google Scholar] [CrossRef]
- Severino, P.; D’Amato, A.; Pucci, M.; Infusino, F.; Adamo, F.; Birtolo, L.I.; Netti, L.; Montefusco, G.; Chimenti, C.; Lavalle, C.; et al. Ischemic Heart Disease Pathophysiology Paradigms Overview: From Plaque Activation to Microvascular Dysfunction. Int. J. Mol. Sci. 2020, 21, 8118. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, J.; Liu, Y.; Xu, D. Sialic acid metabolism as a potential therapeutic target of atherosclerosis. Lipids Health Dis. 2019, 18, 173. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, R.; Lu, X.; Kassab, G.S. Role of glycocalyx in flow-induced production of nitric oxide and reactive oxygen species. Free Radic. Biol. Med. 2009, 47, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Psefteli, P.M.; Kitscha, P.; Vizcay, G.; Fleck, R.; Chapple, S.J.; Mann, G.E.; Fowler, M.; Siow, R.C. Glycocalyx sialic acids regulate Nrf2-mediated signaling by fluid shear stress in human endothelial cells. Redox Biol. 2021, 38, 101816. [Google Scholar] [CrossRef]
- Betteridge, K.B.; Arkill, K.P.; Neal, C.R.; Harper, S.J.; Foster, R.R.; Satchell, S.C.; Bates, D.O.; Salmon, A.H.J. Sialic acids regulate microvessel permeability, revealed by novel in vivo studies of endothelial glycocalyx structure and function. J. Physiol. 2017, 595, 5015–5035. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Gyulay, G.; Mitchell, M.; White, E.; Trigatti, B.L.; Igdoura, S.A. Hypomorphic sialidase expression decreases serum cholesterol by downregulation of VLDL production in mice. J. Lipid Res. 2012, 53, 2573–2585. [Google Scholar] [CrossRef]
- White, E.J.; Gyulay, G.; Lhotak, S.; Szewczyk, M.M.; Chong, T.; Fuller, M.T.; Dadoo, O.; Fox-Robichaud, A.E.; Austin, R.C.; Trigatti, B.L.; et al. Sialidase down-regulation reduces non-HDL cholesterol, inhibits leukocyte transmigration, and attenuates atherosclerosis in ApoE knockout mice. J. Biol. Chem. 2018, 293, 14689–14706. [Google Scholar] [CrossRef]
- Gayral, S.; Garnotel, R.; Castaing-Berthou, A.; Blaise, S.; Fougerat, A.; Berge, E.; Montheil, A.; Malet, N.; Wymann, M.P.; Maurice, P.; et al. Elastin-derived peptides potentiate atherosclerosis through the immune Neu1-PI3Kgamma pathway. Cardiovasc. Res. 2014, 102, 118–127. [Google Scholar] [CrossRef]
- Demina, E.P.; Smutova, V.; Pan, X.; Fougerat, A.; Guo, T.; Zou, C.; Chakraberty, R.; Snarr, B.D.; Shiao, T.C.; Roy, R.; et al. Neuraminidases 1 and 3 Trigger Atherosclerosis by Desialylating Low-Density Lipoproteins and Increasing Their Uptake by Macrophages. J. Am. Heart Assoc. 2021, 10, e018756. [Google Scholar] [CrossRef]
- Kunjathoor, V.V.; Febbraio, M.; Podrez, E.A.; Moore, K.J.; Andersson, L.; Koehn, S.; Rhee, J.S.; Silverstein, R.; Hoff, H.F.; Freeman, M.W. Scavenger receptors class A-I/II and CD36 are the principal receptors responsible for the uptake of modified low density lipoprotein leading to lipid loading in macrophages. J. Biol. Chem. 2002, 277, 49982–49988. [Google Scholar] [CrossRef]
- Cuniberti, L.A.; Martinez, V.; Schachter, J.; Magarinos, G.; Meckert, P.C.; Laguens, R.P.; Levenson, J.; Werba, J.P. Sialic acid as a protective barrier against neointima development. Atherosclerosis 2005, 181, 225–231. [Google Scholar] [CrossRef]
- Moon, S.K.; Cho, S.H.; Kim, K.W.; Jeon, J.H.; Ko, J.H.; Kim, B.Y.; Kim, C.H. Overexpression of membrane sialic acid-specific sialidase Neu3 inhibits matrix metalloproteinase-9 expression in vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2007, 356, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Hinek, A.; Bodnaruk, T.D.; Bunda, S.; Wang, Y.; Liu, K. Neuraminidase-1, a subunit of the cell surface elastin receptor, desialylates and functionally inactivates adjacent receptors interacting with the mitogenic growth factors PDGF-BB and IGF-2. Am. J. Pathol. 2008, 173, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.G.; Byrne, H.M.; Macaskill, C.; Myerscough, M.R. A two-phase model of early fibrous cap formation in atherosclerosis. J. Theor. Biol. 2018, 456, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Swirski, F.K.; Nahrendorf, M. Cardioimmunology: The immune system in cardiac homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Nahrendorf, M.; Pittet, M.J.; Swirski, F.K. Monocytes: Protagonists of infarct inflammation and repair after myocardial infarction. Circulation 2010, 121, 2437–2445. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Regulation of the inflammatory response in cardiac repair. Circ. Res. 2012, 110, 159–173. [Google Scholar] [CrossRef]
- Hilgendorf, I.; Gerhardt, L.M.; Tan, T.C.; Winter, C.; Holderried, T.A.; Chousterman, B.G.; Iwamoto, Y.; Liao, R.; Zirlik, A.; Scherer-Crosbie, M.; et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ. Res. 2014, 114, 1611–1622. [Google Scholar] [CrossRef]
- Guo, Z.; Fan, D.; Liu, F.Y.; Ma, S.Q.; An, P.; Yang, D.; Wang, M.Y.; Yang, Z.; Tang, Q.Z. NEU1 Regulates Mitochondrial Energy Metabolism and Oxidative Stress Post-myocardial Infarction in Mice via the SIRT1/PGC-1 Alpha Axis. Front. Cardiovasc. Med. 2022, 9, 821317. [Google Scholar] [CrossRef]
- Hanson, V.A.; Shettigar, U.R.; Loungani, R.R.; Nadijcka, M.D. Plasma sialidase activity in acute myocardial infarction. Am. Heart J. 1987, 114, 59–63. [Google Scholar] [CrossRef]
- Suer Gokmen, S.; Kazezoglu, C.; Sunar, B.; Ozcelik, F.; Gungor, O.; Yorulmaz, F.; Gulen, S. Relationship between serum sialic acids, sialic acid-rich inflammation-sensitive proteins and cell damage in patients with acute myocardial infarction. Clin. Chem. Lab Med. 2006, 44, 199–206. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, T.T.; Li, Y.; Li, J.; Fan, Y.; Huang, F.Q.; Cai, Y.Y.; Ma, G.; Liu, J.F.; Chen, Q.Q.; et al. Functional Metabolomics Characterizes a Key Role for N-Acetylneuraminic Acid in Coronary Artery Diseases. Circulation 2018, 137, 1374–1390. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, M.; Xiao, L.; Wei, H.; Wen, Z.; Hu, D.; Yu, B.; Sun, Y.; Gao, J.; Shen, X.; et al. Prognostic Value of Elevated Levels of Plasma N-Acetylneuraminic Acid in Patients With Heart Failure. Circ Heart Fail 2021, 14, e008459. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Sadoshima, J. Mechanisms of physiological and pathological cardiac hypertrophy. Nat. Rev. Cardiol. 2018, 15, 387–407. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Schelbert, E.B.; Diez, J.; Butler, J. Myocardial Interstitial Fibrosis in Heart Failure: Biological and Translational Perspectives. J. Am. Coll. Cardiol. 2018, 71, 1696–1706. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Kim, S.J.; Chung, T.W.; Choi, H.J.; Kwak, C.H.; Song, K.H.; Suh, S.J.; Kwon, K.M.; Chang, Y.C.; Park, Y.G.; Chang, H.W.; et al. Ganglioside GM3 participates in the TGF-beta1-induced epithelial-mesenchymal transition of human lens epithelial cells. Biochem. J. 2013, 449, 241–251. [Google Scholar] [CrossRef]
- Ghiroldi, A.; Piccoli, M.; Creo, P.; Cirillo, F.; Rota, P.; D’Imperio, S.; Ciconte, G.; Monasky, M.M.; Micaglio, E.; Garatti, A.; et al. Role of sialidase Neu3 and ganglioside GM3 in cardiac fibroblasts activation. Biochem. J. 2020, 477, 3401–3415. [Google Scholar] [CrossRef]
- Riehle, C.; Bauersachs, J. Of mice and men: Models and mechanisms of diabetic cardiomyopathy. Basic Res. Cardiol. 2018, 114, 2. [Google Scholar] [CrossRef]
- Mitry, M.A.; Edwards, J.G. Doxorubicin induced heart failure: Phenotype and molecular mechanisms. Int. J. Cardiol. Heart Vasc. 2016, 10, 17–24. [Google Scholar] [CrossRef]
- Guo, Z.; Tuo, H.; Tang, N.; Liu, F.Y.; Ma, S.Q.; An, P.; Yang, D.; Wang, M.Y.; Fan, D.; Yang, Z.; et al. Neuraminidase 1 deficiency attenuates cardiac dysfunction, oxidative stress, fibrosis, inflammatory via AMPK-SIRT3 pathway in diabetic cardiomyopathy mice. Int. J. Biol. Sci. 2022, 18, 826–840. [Google Scholar] [CrossRef]
- Qin, Y.; Lv, C.; Zhang, X.; Ruan, W.; Xu, X.; Chen, C.; Ji, X.; Lu, L.; Guo, X. Neuraminidase1 Inhibitor Protects Against Doxorubicin-Induced Cardiotoxicity via Suppressing Drp1-Dependent Mitophagy. Front. Cell Dev. Biol. 2021, 9, 802502. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.M.; da Silva, R.L.; Vieira, J.L.; Fattah, T.; Lueneberg, M.E.; Gottschall, C.A. Role of vascular inflammation in coronary artery disease: Potential of anti-inflammatory drugs in the prevention of atherothrombosis. Inflammation and anti-inflammatory drugs in coronary artery disease. Am. J. Cardiovasc. Drugs 2015, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, G.; Eklund, G.A.; Gullberg, B.; Rastam, L. Serum sialic acid concentration and cardiovascular mortality. BMJ 1991, 302, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Haq, M.; Haq, S.; Tutt, P.; Crook, M. Serum total sialic acid and lipid-associated sialic acid in normal individuals and patients with myocardial infarction, and their relationship to acute phase proteins. Ann. Clin. Biochem. 1993, 30 Pt 4, 383–386. [Google Scholar] [CrossRef]
- Amiri Majd, A.; Rezaei, M.; Goodarzi, M.; Homayoonfar, S. Some biomarkers in acute myocardial infarction. J. Res. Health Sci. 2007, 7, 11–19. [Google Scholar]
- Rajendiran, K.S.; Ananthanarayanan, R.H.; Satheesh, S.; Rajappa, M. Elevated levels of serum sialic acid and high-sensitivity C-reactive protein: Markers of systemic inflammation in patients with chronic heart failure. Br. J. Biomed. Sci. 2014, 71, 29–32. [Google Scholar] [CrossRef]
- Moussa, M.A.; Alsaeid, M.; Refai, T.M.; Abdella, N.; Al-Sheikh, N.; Gomez, J.E. Association of serum sialic acid with cardiovascular metabolic risk factors in Kuwaiti children and adolescents with type 1 diabetes. Metabolism 2004, 53, 638–643. [Google Scholar] [CrossRef]
- Sathiyapriya, V.; Nandeesha, H.; Selvaraj, N.; Bobby, Z.; Agrawal, A.; Pavithran, P. Association between protein-bound sialic acid and high-sensitivity C-reactive protein in essential hypertension: A possible indication of underlying cardiovascular risk. Angiology 2008, 59, 721–726. [Google Scholar] [CrossRef]
- Crook, M.; Haq, M.; Haq, S.; Tutt, P. Plasma sialic acid and acute-phase proteins in patients with myocardial infarction. Angiology 1994, 45, 709–715. [Google Scholar] [CrossRef]
- Cheeseman, J.; Kuhnle, G.; Spencer, D.I.R.; Osborn, H.M.I. Assays for the identification and quantification of sialic acids: Challenges, opportunities and future perspectives. Bioorg. Med. Chem. 2021, 30, 115882. [Google Scholar] [CrossRef]
- Gopaul, K.P.; Crook, M.A. Sialic acid: A novel marker of cardiovascular disease? Clin. Biochem. 2006, 39, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, B.V.; Rawls, K.D.; Kolling, G.L.; Vinnakota, K.C.; Wallqvist, A.; Papin, J.A. Identifying functional metabolic shifts in heart failure with the integration of omics data and a heart-specific, genome-scale model. Cell Rep. 2021, 34, 108836. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.S.; van den Berg, T.K.; Mullen, G.E. Sialoadhesin—A macrophage-restricted marker of immunoregulation and inflammation. Immunology 2013, 138, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Tisoncik, J.R.; Guo, Y.; Cordero, K.S.; Yu, J.; Wang, J.; Cao, Y.; Rong, L. Identification of critical residues of influenza neuraminidase in viral particle release. Virol. J. 2011, 8, 14. [Google Scholar] [CrossRef]
- von Itzstein, M. The war against influenza: Discovery and development of sialidase inhibitors. Nat. Rev. Drug. Discov. 2007, 6, 967–974. [Google Scholar] [CrossRef]
- Smeeth, L.; Thomas, S.L.; Hall, A.J.; Hubbard, R.; Farrington, P.; Vallance, P. Risk of myocardial infarction and stroke after acute infection or vaccination. N. Engl. J. Med. 2004, 351, 2611–2618. [Google Scholar] [CrossRef]
- Madjid, M.; Miller, C.C.; Zarubaev, V.V.; Marinich, I.G.; Kiselev, O.I.; Lobzin, Y.V.; Filippov, A.E.; Casscells, S.W., 3rd. Influenza epidemics and acute respiratory disease activity are associated with a surge in autopsy-confirmed coronary heart disease death: Results from 8 years of autopsies in 34,892 subjects. Eur. Heart J. 2007, 28, 1205–1210. [Google Scholar] [CrossRef]
- Lou, Y.; Gan, J.; Peppercorn, A.; Gould, E.; Weller, S.; Piscitelli, S.C.; Patel, P. Effect of intravenous zanamivir on cardiac repolarization. Pharmacotherapy 2013, 33, 701–709. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Properties of the Four Mammalian NEU Enzymes | ||||
|---|---|---|---|---|
| NEU1 | NEU2 | NEU3 | NEU4 | |
| Human Gene Location | chromosome 6p21.3 [23] | chromosome 2q37 [24] | chromosome 11q13.5 [24] | chromosome 2q37.3 [24] |
| Murine Gene Location | chromosome 17 [25] | chromosome 1 [26] | chromosome 7 [24] | chromosome 10 [24] |
| Subcellular localisation | Lysosomal, translocation towards the plasma membrane upon different stimuli [7,27,28] | Cytosolic [24] | Associated with the plasma membrane [29] | Murine NEU4a and b, human NEU4S: ER membranes; human NEU4L: mitochondria, lysosomes [30,31] |
| Expression pattern | kidneys, skeletal muscle, lung, placenta, brain, pancreas, inflammatory cells and cardiomyocytes [7,27,32,33] | Muscle-specific isoform [7,34] | Adrenal glands, heart, thymus, skeletal muscle and testis [7] | Brain, heart, placenta, liver and skeletal muscle [31] |
| Substrate preferences | Oligosaccharides with an α2,3 linkage [24] | Oligosaccharides, gangliosides, glycoproteins [24,26,35] | Gangliosides, most preferable with an α2,3 and α2,6 linkage [24] | Oligosaccharides, gangliosides, glycoproteins [24] |
| Physiological functions | Regulates exocytosis, modulator of inflammatory resposnse [27,36,37] | Myoblast and neuronal cell differentiation [38,39,40] | Neuronal cell differentitation, focal adhesion, cell invasion, cell survival, proliferation [41,42,43] | Neural differentiation, mitochondrial neuronal apoptosis [44,45] |
| NEU Inhibitors in Experimental Animal Studies | |||
|---|---|---|---|
| Drug | Model | Effects | References |
| Zanamivir (antiviral drug) | (1) Transverse aortic constriction (TAC)-induced cardiac hypertrophy (mice and rat model) (1) Isoproterenol (ISO)-induced hypertrophic rat models (2) Myocardial ischemia rat model (2) ISO-induced injury rat model, permanent left anterior descending coronary artery ligation | (1), (2) beneficial effects after post-treatment, reduced cardiac, hypertrophy, fibrosis, improved cardiac function | (1) [33] (2) [112] |
| Oseltamivir (antiviral drug) | (1) TAC-induced cardiac hypertrophy (mice and rat model) (1) ISO-induced hypertrophic rat models (2) Myocardial ischemia rat model (2) ISO-induced injury rat model, permanent left anterior descending coronary artery ligation (3) Doxorubicin (DOX) induced cardiomyopathy rat model (4) ISO- and Angiotensin (ANG)II-induced heart failure mouse models | (1), (2) beneficial effects after post-treatment, reduced cardiac, hypertrophy, fibrosis, improved cardiac function (3) beneficial effects after pre- and co-treatment (4) beneficial effects of post-treatment | (1) [33] (2) [112] (3) [122] (4) [113] |
| Compound-C09 (NEU1 inhibitor) | (5) TAC-induced cardiac hypertrophy | (5) beneficial effects after post-treatment, reduced cardiac, hypertrophy, fibrosis, improved cardiac function | (5) [33] |
| Danamivir | (6) Apolipoprotein E deficient (Apo E-/-) atherosclerosis mouse model | (6) reduced atherosclerotic lesion size | (6) [99] |
| C9-BA-DANA | (7) Apo E-/-atherosclerosis mouse model | (7) reduced atherosclerotic lesion size | (7) [99] |
| CG17701 | (8) Apo E-/-atherosclerosis mouse model | (8) reduced atherosclerotic lesion size | (8) [99] |
| CG14601 | (9) Apo E-/-atherosclerosis mouse model (1) (10) Low density lipoprotein receptor (Ldlr)-/-and high fat diet atherosclerosis mouse model | (9), (10) reduced atherosclerotic lesion size | (9), (10) [99] |
| CG22601 | (11) Apo E-/-atherosclerosis mouse model (12) Ldlr-/-and high fat diet atherosclerosis mouse model | (11), (12) reduced atherosclerotic lesion size | (11), (12) [99] |
| NEU Inhibitors Targeting Cardiovascular Disease in Clinical Trials | |||
|---|---|---|---|
| Drug | Study Description | Effects | Trial Registration |
| Zanamivir (antiviral drug) | Effect of intravenous zanamivir on cardiac conduction in healthy volunteers (n = 40); randomized controlled trial | Completed: Intravenous zanamivir does not affect cardiac repolarization | ClinicTrials.gov NCT01353729 |
| Oseltamivir (antiviral drug) | the effects of neuraminidase inhibitor oseltamivir in patients with chronic heart failure (n = 388); randomized, open-label, blank-controlled study | Ongoing | ClinicTrials.gov NCT05008679 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heimerl, M.; Gausepohl, T.; Mueller, J.H.; Ricke-Hoch, M. Neuraminidases—Key Players in the Inflammatory Response after Pathophysiological Cardiac Stress and Potential New Therapeutic Targets in Cardiac Disease. Biology 2022, 11, 1229. https://doi.org/10.3390/biology11081229
Heimerl M, Gausepohl T, Mueller JH, Ricke-Hoch M. Neuraminidases—Key Players in the Inflammatory Response after Pathophysiological Cardiac Stress and Potential New Therapeutic Targets in Cardiac Disease. Biology. 2022; 11(8):1229. https://doi.org/10.3390/biology11081229
Chicago/Turabian StyleHeimerl, Maren, Thomas Gausepohl, Julia H. Mueller, and Melanie Ricke-Hoch. 2022. "Neuraminidases—Key Players in the Inflammatory Response after Pathophysiological Cardiac Stress and Potential New Therapeutic Targets in Cardiac Disease" Biology 11, no. 8: 1229. https://doi.org/10.3390/biology11081229
APA StyleHeimerl, M., Gausepohl, T., Mueller, J. H., & Ricke-Hoch, M. (2022). Neuraminidases—Key Players in the Inflammatory Response after Pathophysiological Cardiac Stress and Potential New Therapeutic Targets in Cardiac Disease. Biology, 11(8), 1229. https://doi.org/10.3390/biology11081229