
Effects of Ventromedial Hypothalamic Nucleus (VMN) Aromatase Gene Knockdown on VMN Glycogen Metabolism and Glucoregulatory Neurotransmission
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Neural Tissue Sample Procurement
2.4. Western Blot Analysis of VMN Target Proteins
2.5. LC-ESI-MS Analysis of VMN Glycogen Concentrations
2.6. LC-ESI-MS Analysis of VMN Estradiol Concentrations
2.7. Plasma Glucose and Counter-Regulatory Hormone Analyses
2.8. Statistical Analyses
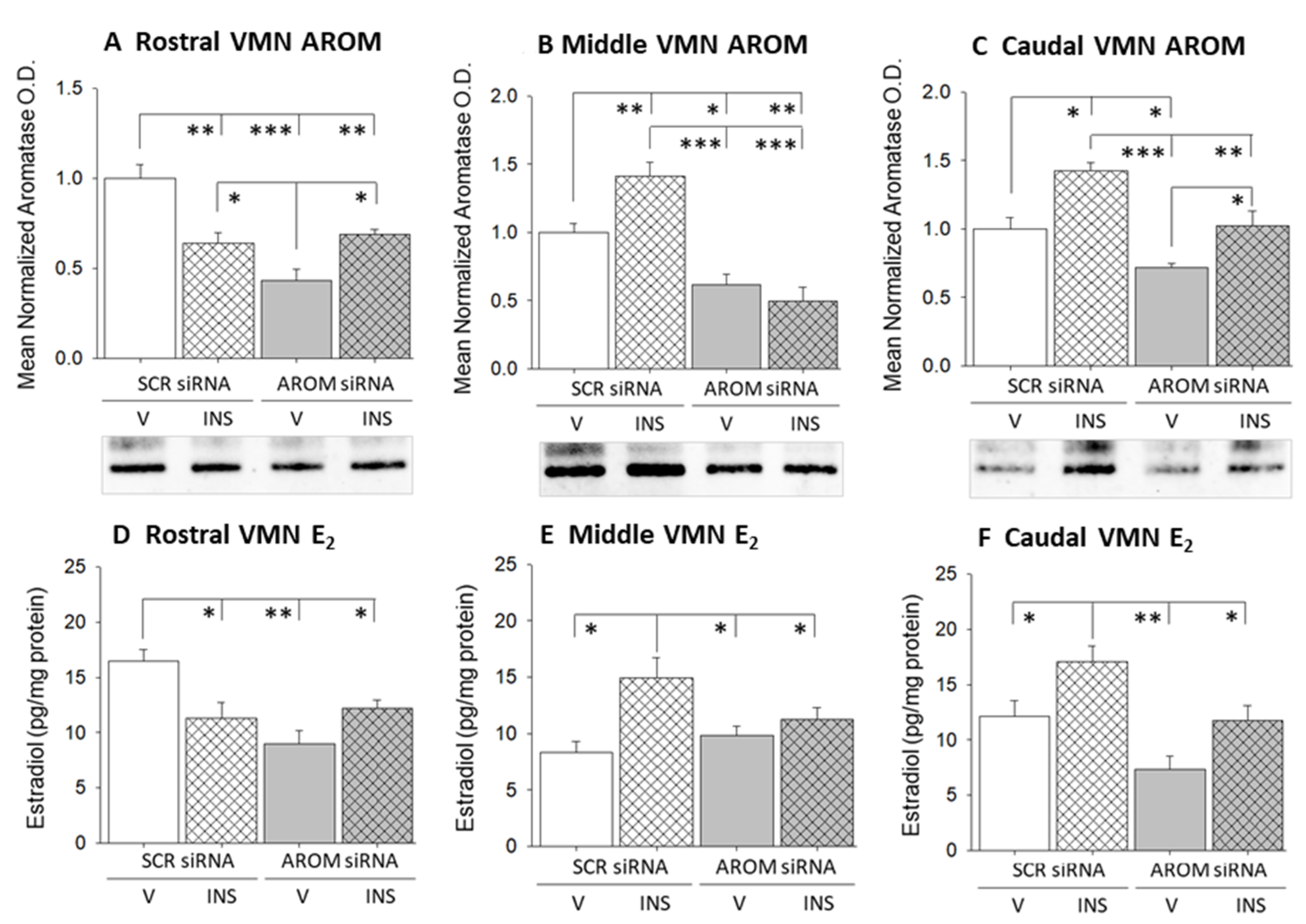
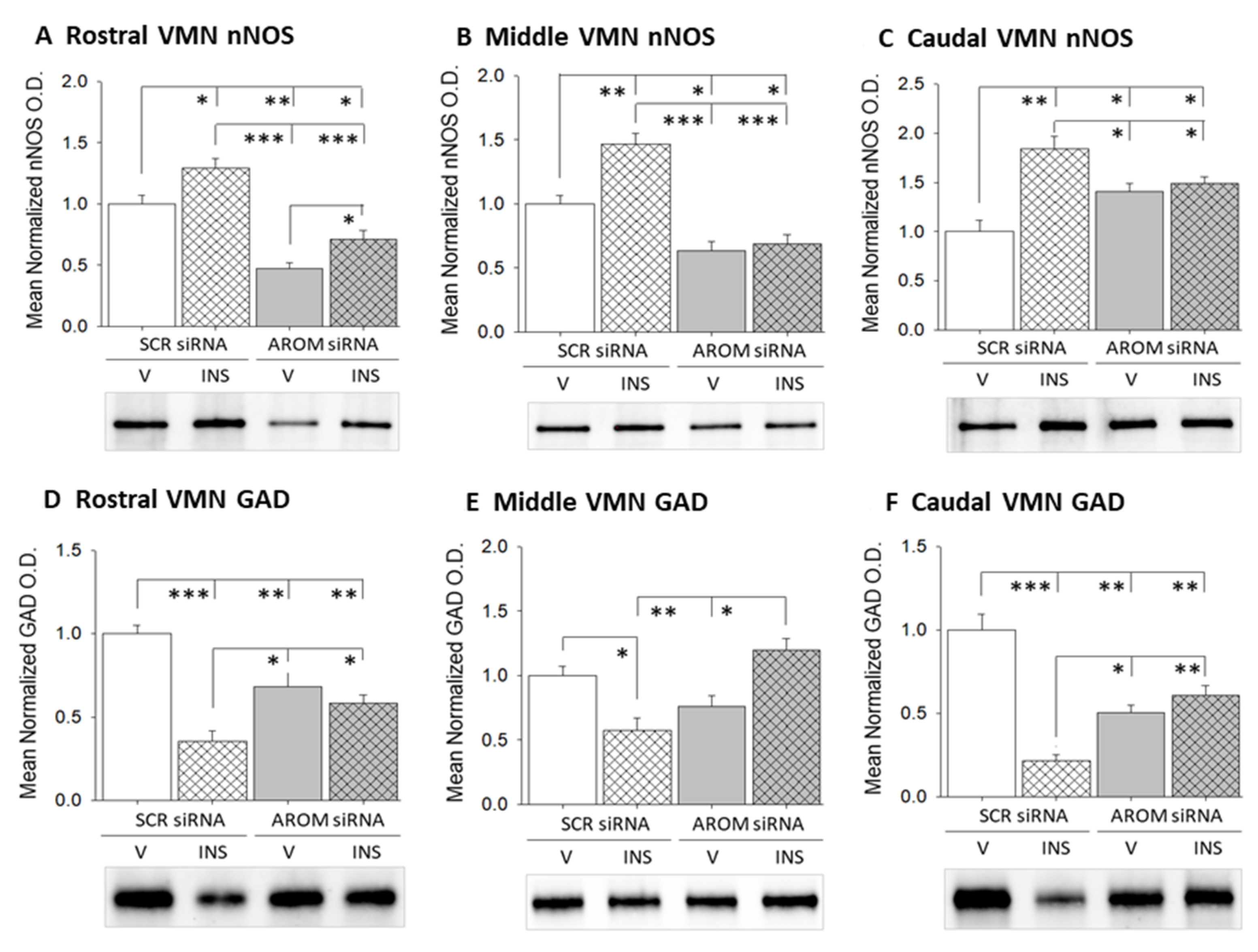
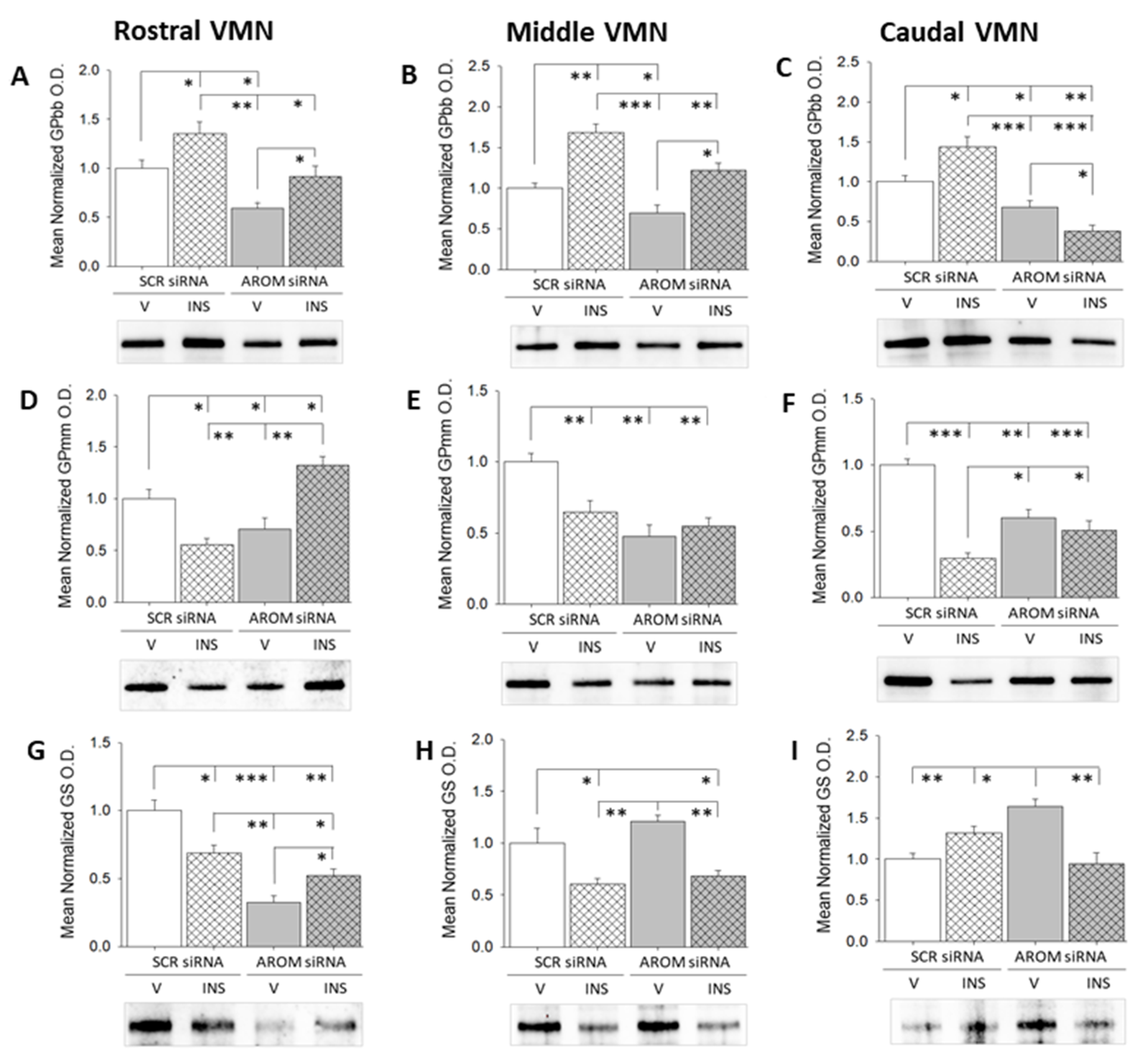
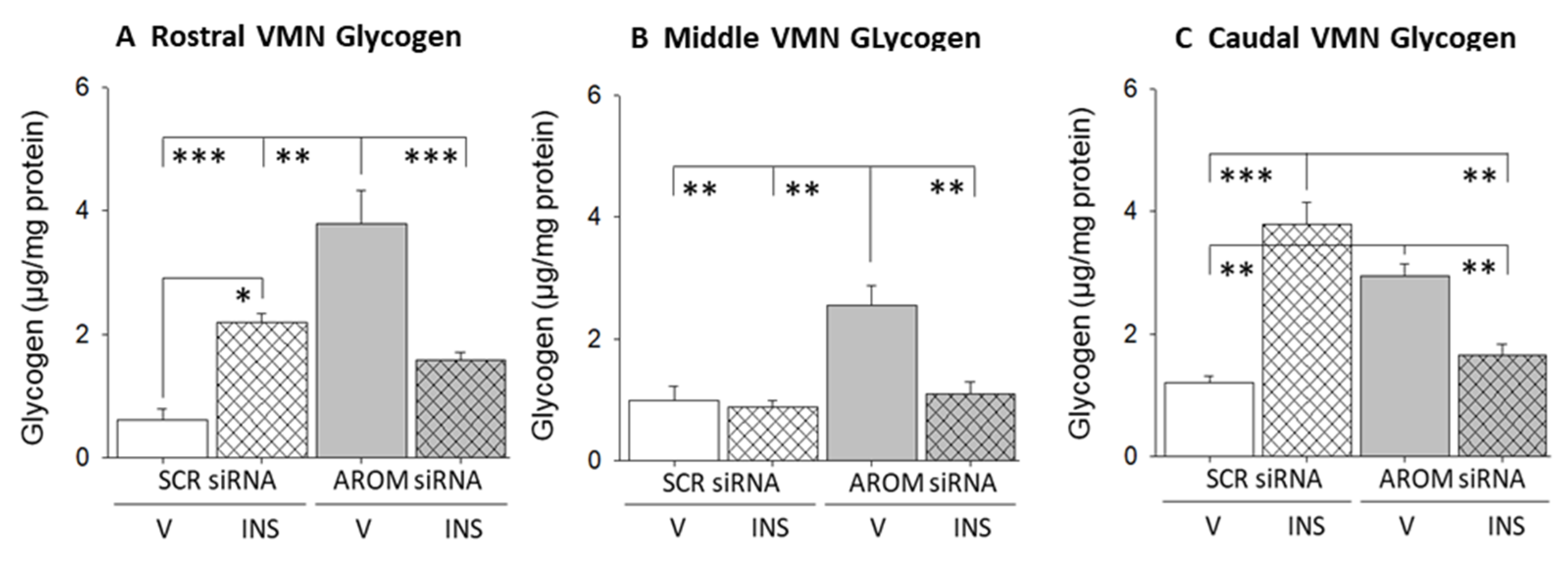
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPK | 5′-AMP-activated protein kinase |
| ER | estrogen receptor |
| ERα | ER-alpha |
| ERβ | ER-beta |
| GAD | glutamate decarboxylase65/67 |
| GP | glycogen phosphorylase |
| GPbb | GP-brain type |
| GPmm | GP-muscle type |
| INS | insulin |
| IIH | insulin-induced hypoglycemia |
| nNOS | neuronal nitric oxide synthase |
| NO | nitric oxide |
| VMN | ventromedial hypothalamic nucleus |
References
- Rudolph, L.M.; Cornil, C.A.; Mittelman-Smith, M.A.; Rainville, J.R.; Remage-Healey, L.; Sinchak, K.; Micevych, P.E. Actions of Steroids: New Neurotransmitters. J. Neurosci. 2016, 36, 11449–11458. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, S.C.; Korach, K.S. Estrogen Receptors: New Directions in the New Millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Lauber, M.E.; Lichtensteiger, W. Pre- and postnatal ontogeny of aromatase cytochrome P450 messenger ribonucleic acid expression in the male rat brain studied by in situ hybridization. Endocrinology 1994, 135, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Foidart, A.; Harada, N.; Balthazart, J. Aromatase-immunoreactive cells are present in mouse brain areas that are known to express high levels of aromatase activity. Cell Tissue Res. 1995, 280, 561–574. [Google Scholar] [CrossRef]
- Roselli, C.E.; Klosterman, S.A.; Fasasi, T.A. Sex differences in androgen responsiveness in the rat brain: Regional differences in the induction of aromatase activity. Neuroendocrinology 1996, 64, 139–145. [Google Scholar] [CrossRef]
- Roselli, C.E.; Klosterman, S.A. Sexual differentiation of aromatase activity in the rat brain: Effects of perinatal steroid exposure. Endocrinology 1998, 139, 3193–3201. [Google Scholar] [CrossRef]
- Roselli, C.E.; Abdelgadir, S.E.; Rønnekleiv, O.K.; Klosterman, S.A. Anatomic distribution and regulation of aromatase gene expression in the rat brain. Biol. Reprod. 1998, 58, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Veney, S.L.; Rissman, E.F. Immunolocalization of androgen receptors and aromatase enzyme in the adult musk shrew brain. Neuroendocrinology 2000, 72, 29–36. [Google Scholar] [CrossRef]
- Beck, L.A.; Wade, J. Sexually dimorphic estrogen receptor alpha mRNA expression in the preoptic area and ventromedial hypothalamus of green anole lizards. Horm. Behav. 2009, 55, 398–403. [Google Scholar] [CrossRef]
- Dickens, M.J.; Cornil, C.A.; Balthazart, J. Acute stress differentially affects aromatase activity in specific brain nuclei of adult male and female quail. Endocrinology 2011, 152, 4242–4251. [Google Scholar] [CrossRef]
- Ubuka, T.; Tsutsui, K. Review: Neuroestrogen regulation of socio-sexual behavior of males. Front. Neurosci. 2015, 8, 323. [Google Scholar] [CrossRef] [PubMed]
- Cornil, C.A. On the role of brain aromatase in females—Why are estrogens produced locally when they are available systemically? J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2018, 204, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Balthazart, J.; Cornil, C.A.; Taziaux, M.; Charlier, T.D.; Baillien, M.; Ball, G.F. Rapid changes in production and behavioral action of estrogens. Neuroscience 2006, 138, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Krause, W.C.; Ingraham, H.A. Origins and functions of the ventrolateral VMH: A complex neuronal cluster orchestrating sex differences in metabolism and behavior. Adv. Exp. Med. Biol. 2017, 1043, 199–213. [Google Scholar] [CrossRef]
- Cryer, P.E. Hypoglycemia-associated autonomic failure in diabetes. Handb. Clin. Neurol. 2013, 117, 295–307. [Google Scholar] [CrossRef]
- Cryer, P.E. Glycemic goals in diabetes: Trade-off between glycemic control and iatrogenic hypoglycemia. Diabetes 2014, 63, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.; LeSauter, J.; Dupre, C.; Pfaff, D.W. Relationship of arousal to circadian anticipatory behavior: Ventromedial hypothalamus: One node in a hunger-arousal network. Eur. J. Neurosci. 2009, 30, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Micevych, P.E.; Meisel, R.L. Integrating neural circuits controlling female sexual behavior. Front. Syst. Neurosci. 2017, 11, 42. [Google Scholar] [CrossRef]
- Shimazu, T.; Minokoshi, T. Systemic glucoregulation by glucose-sensing neurons in the ventromedial hypothalamic nucleus (VMN). J. Endocr. Soc. 2017, 1, 449–450. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Y.; Hu, F. AMPK in the ventromedial nucleus of the hypothalamus: A key regulatory for thermogenesis. Front. Endocrinol. 2020, 11, 578830. [Google Scholar] [CrossRef]
- Henderson, L.A.; Macefield, V.G. The role of the dorsomedial and ventromedial hypothalamus in regulating behaviorally coupled and resting autonomic drive. Handb. Clin. Neurol. 2021, 180, 167–200. [Google Scholar] [CrossRef]
- Watts, A.G.; Donovan, C.M. Sweet talk in the brain: Glucosensing, neural networks, and hypoglycemic counterregulation. Front. Neuroendocrinology 2010, 31, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Donovan, C.M.; Watts, A.G. Peripheral and central glucose sensing in hypoglycemic detection. Physiology 2014, 29, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Oomura, Y.; Ono, H.; Ooyama, H.; Wayner, M.J. Glucose and osmosensitive neurons of the rat hypothalamus. Nature 1969, 222, 282–284. [Google Scholar] [CrossRef]
- Ashford, M.L.J.; Boden, P.R.; Treherne, J.M. Glucose-induced excitation of hypothalamic neurons is mediated by ATP-sensitive K+ channels. Pfugers Arch. 1990, 415, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Silver, I.A.; Erecińska, M. Glucose-induced intracellular ion changes in sugar-sensitive hypothalamic neurons. J. Neurophysiol. 1998, 79, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Borg, M.A.; Sherwin, R.S.; Borg, W.P.; Tamborlane, W.V.; Shulman, G.I. Local ventromedial hypothalamus glucose perfusion blocks counterregulation during systemic hypoglycemia in awake rats. J. Clin. Investig. 1997, 99, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Borg, M.A.; Tamborlane, W.V.; Shulman, G.I.; Sherwin, R.S. Local lactate perfusion of the ventromedial hypothalamus suppresses hypoglycemic counterregulation. Diabetes 2003, 52, 663–666. [Google Scholar] [CrossRef]
- Chan, O.; Zhu, W.; Ding, Y.; McCrimmon, R.J.; Sherwin, R.S. Blockade of GABA(A) receptors in the ventromedial hypothalamus further stimulates glucagon and sympathoadrenal but not the hypothalamo-pituitary-adrenal response to hypoglycemia. Diabetes 2006, 55, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Fioramonti, X.; Marsollier, N.; Song, Z.; Fakira, K.A.; Patel, R.M.; Brown, S.; Duparc, T.; Pica-Mendez, A.; Sanders, N.M.; Knauf, C.; et al. Ventromedial hypothalamic nitric oxide production is necessary for hypoglycemia detection and counterregulation. Diabetes 2010, 59, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Routh, V.H.; Hao, L.; Santiago, A.M.; Sheng, Z.; Zhou, C. Hypothalamic glucose sensing: Making ends meet. Front. Syst. Neurosci. 2014, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.H.; Bheemanapally, K.; Alhamami, H.N.; Briski, K.P. Effects of intracerebroventricular glycogen phosphorylase inhibitor CP-316,819 infusion on hypothalamic glycogen content and metabolic neuron AMPK activity and neurotransmitter expression in the male rat. J. Mol. Neurosci. 2020, 70, 647–658. [Google Scholar] [CrossRef]
- Ali, M.H.; Alshamrani, A.A.; Napit, P.R.; Briski, K.P. Single-cell multiplex qPCR evidence for sex-dimorphic glutamate decarboxylase, estrogen receptor, and 5′-AMP-activated protein kinase alpha subunit mRNA expression by ventromedial hypothalamic nucleus GABAergic neurons. J. Chem. Neuroanat. 2022, 27, 102132. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.S.M.H.; Uddin, M.M.; Mandal, S.K.; Ibrahim, M.M.H.; Alhamami, H.N.; Briski, K.P. Sex differences in forebrain estrogen receptor regulation of hypoglycemic patterns of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus gluco-regulatory neurotransmitter and astrocyte glycogen metabolic enzyme expression. Neuropeptides 2018, 72, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Bheemanapally, K.; Ibrahim, M.M.H.; Briski, K.P. Sex-dimorphic neuroestradiol regulation of ventromedial hypothalamic nucleus glucoregulatory transmitter and glycogen metabolism enzyme protein expression in the rat. BMC Neurosci. 2020, 21, 51. [Google Scholar] [CrossRef]
- Stobart, J.L.; Anderson, C.M. Role of astrocytes as gatekeepers of neuronal energy supply. Front. Cell. Neurosci. 2013, 7, 1–21. [Google Scholar] [CrossRef]
- Laming, P.R.; Kimelberg, H.; Robinson, S.; Salm, A.; Hawrylak, N.; Müller, C.; Roots, B.; Ng, K. Neuronal-glial interactions and behaviour. Neurosci. Biobehav. Rev. 2000, 24, 295–340. [Google Scholar]
- Gruetter, R. Glycogen: The forgotten cerebral energy store. J. Neurosci. Res. 2003, 74, 179–183. [Google Scholar] [CrossRef]
- Brown, A.M. Brain glycogen re-awakened. J. Neurochem. 2004, 89, 537–552. [Google Scholar] [CrossRef]
- Bélanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef]
- Nadeau, O.W.; Fontes, j.D.; Carlson, G.M. The regulation of glycogenolysis in the brain. J. Biol. Chem. 2018, 293, 7099–7109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.S.; Pedersen, S.E.; Walls, A.B.; Waagepetersen, H.S.; Bak, L.K. Isoform-selective regulation of glycogen phosphorylase by energy deprivation and phosphorylation in astrocytes. Glia 2015, 63, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Alhamami, H.N.; Alshamrani, A.; Briski, K.P. Inhibition of glycogen phosphorylase stimulates ventromedial hypothalamic nucleus AMP-activated protein kinase: Activity and neuronal nitric oxide synthase protein expression in male rats. Physiol. Rep. 2017, 5, e13484. [Google Scholar] [CrossRef]
- Bheemanapally, K.; Ibrahim, M.M.H.; Briski, K.P. Combinatory high-resolution microdissection/ultra-performance liquid chromatographic-mass spectrometry approach for small tissue volume analysis of rat brain glycogen. J. Pharmaceut. Biomed. Anal. 2020, 178, 112884. [Google Scholar] [CrossRef]
- Gilda, J.E.; Gomes, A.V. Western blotting using in-gel protein labeling as a normalization control: Stain-free technology. Methods Mol. Biol. 2015, 1295, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.P. Tubulin or not tubulin: Heading toward total protein staining as loading control in Western blots. Proteomics 2017, 17, 1600189. [Google Scholar] [CrossRef] [PubMed]
- Bheemanapally, K.; Ibrahim, M.M.; Briski, K.P. Ultra-high-performance liquid chromatography-electrospray ionization-mass spectrometry for high-neuroanatomical resolution quantification of brain estradiol concentrations. J. Pharmaceut. Biomed. Anal. 2020, 191, 113606. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Mahmood, A.S.M.H.; Ibrahim, M.M.H.; Briski, K.P. Sex dimorphic estrogen receptor regulation of ventromedial hypothalamic nucleus glucoregulatory neuron adrenergic receptor expression in hypoglycemic male and female rats. Brain Res. 2019, 1720, 146311. [Google Scholar] [CrossRef]
- Ibrahim, M.M.H.; Alhamami, H.N.; Briski, K.P. Norepinephrine regulation of ventromedial hypothalamic nucleus metabolic transmitter biomarker and astrocyte enzyme and receptor expression: Impact of 5′ AMP-activated protein kinase. Brain Res. 2019, 1711, 48–57. [Google Scholar] [CrossRef]
- Zhang, Q.G.; Wang, R.; Tang, H.; Dong, Y.; Chan, A.; Sareddy, G.R.; Vadlamadui, R.K.; Brann, D.W. Brain-derived estrogen exerts anti-inflammatory and neuroprotective actions in the rat hippocampus. Mol. Cell. Endocrinol. 2014, 389, 84–91. [Google Scholar] [CrossRef]
- Oride, A.; Kanasaki, H.; Tumurbaatar, T.; Zolzaya, T.; Okada, H.; Hara, T.; Kyo, S. Effects of the Fertility Drugs Clomiphene Citrate and Letrozole on Kiss-1 Expression in Hypothalamic Kiss-1-Expressing Cell Models. Reprod. Sci. 2020, 27, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Cornil, C.A.; Ball, G.F.; Balthazart, J. Sexually differentiated and neuroanatomically specific co-expression of aromatase neurons and GAD67 in the male and female quail brain. Eur. J. Neurosci. 2020, 52, 2963–2981. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rostral VMN | Middle VMN | Caudal VMN | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SCR a | siRNA | AROM b | siRNA | SCR | siRNA | AROM | siRNA | SCR | siRNA | AROM | siRNA | |
| V c | INS d | V | INS | V | INS | V | INS | V | INS | V | INS | |
| AROM | - | ↓ e | ↓ | ↑ f | - | ↑ | ↓ | N.C. g | - | ↑ | ↓ | ↑ |
| Estradiol | - | ↓ | ↓ | N.C | - | ↑ | N.C. | N.C. | - | ↑ | N.C. | N.C. |
| nNOS h | - | ↑ | ↓ | ↑ | - | ↑ | ↓ | N.C. | - | ↑ | ↑ | N.C. |
| GAD i | - | ↓ | ↓ | N.C. | - | ↓ | N.C. | ↑ | - | ↓ | ↓ | N.C. |
| GPbb j | - | ↑ | ↓ | ↑ | - | ↑ | ↓ | ↑ | - | ↑ | ↓ | ↓ |
| GPmm k | - | ↓ | N.C. | ↑ | - | ↓ | ↓ | N.C. | - | ↓ | ↓ | N.C. |
| GS l | - | ↓ | ↓ | ↑ | - | ↓ | N.C. | ↓ | - | N.C | ↑ | ↓ |
| Glycogen | - | ↑ | ↑ | ↓ | - | N.C. | ↑ | ↓ | - | ↑ | ↑ | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briski, K.P.; Mahmood, A.S.M.H.; Uddin, M.M.; Ibrahim, M.M.H.; Bheemanapally, K. Effects of Ventromedial Hypothalamic Nucleus (VMN) Aromatase Gene Knockdown on VMN Glycogen Metabolism and Glucoregulatory Neurotransmission. Biology 2023, 12, 242. https://doi.org/10.3390/biology12020242
Briski KP, Mahmood ASMH, Uddin MM, Ibrahim MMH, Bheemanapally K. Effects of Ventromedial Hypothalamic Nucleus (VMN) Aromatase Gene Knockdown on VMN Glycogen Metabolism and Glucoregulatory Neurotransmission. Biology. 2023; 12(2):242. https://doi.org/10.3390/biology12020242
Chicago/Turabian StyleBriski, Karen P., A. S. M. Hasan Mahmood, Md. Main Uddin, Mostafa M. H. Ibrahim, and Khaggeswar Bheemanapally. 2023. "Effects of Ventromedial Hypothalamic Nucleus (VMN) Aromatase Gene Knockdown on VMN Glycogen Metabolism and Glucoregulatory Neurotransmission" Biology 12, no. 2: 242. https://doi.org/10.3390/biology12020242
APA StyleBriski, K. P., Mahmood, A. S. M. H., Uddin, M. M., Ibrahim, M. M. H., & Bheemanapally, K. (2023). Effects of Ventromedial Hypothalamic Nucleus (VMN) Aromatase Gene Knockdown on VMN Glycogen Metabolism and Glucoregulatory Neurotransmission. Biology, 12(2), 242. https://doi.org/10.3390/biology12020242