

Effect of Explant Source on Phenotypic Changes of In Vitro Grown Cannabis Plantlets over Multiple Subcultures



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Explant’s Source Description
2.2. In Vitro Culture Condition and Origin of Subcultures
2.3. Experimental Design and Statistical Analysis
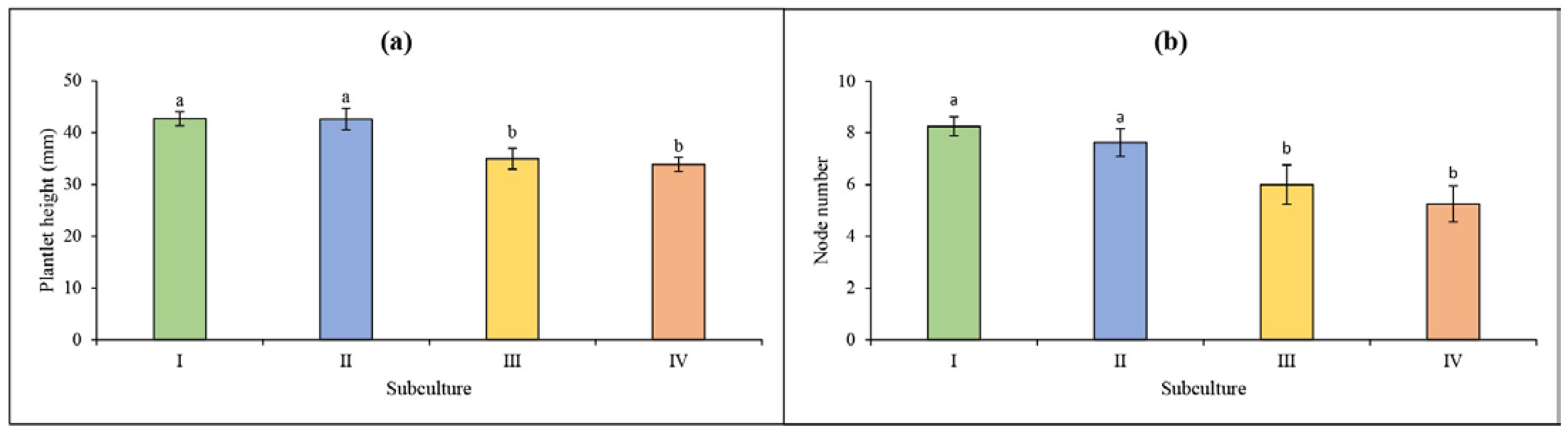
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Phineas Jones, A.M. Recent advances in cannabis biotechnology. Ind. Crops Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- Braithwaite, I.; Bhagavan, C.; Doppen, M.; Kung, S.; Oldfield, K.; Newton-Howes, G. Medicinal applications of cannabis/cannabinoids. Curr. Opin. Psychol. 2021, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, A.; Zafar, U.; Ahmed, W.; Shabbir, M.A.; Sameen, A.; Sahar, A.; Bhat, Z.F.; Kowalczewski, P.Ł.; Jarzębski, M.; Aadil, R.M. Applications of Cannabis sativa L. in food and its therapeutic potential: From a prohibited drug to a nutritional supplement. Molecules 2021, 26, 7699. [Google Scholar] [CrossRef] [PubMed]
- Parvez, A.M.; Lewis, J.D.; Afzal, M.T. Potential of industrial hemp (Cannabis sativa L.) for bioenergy production in Canada: Status, challenges and outlook. Renew. Sustain. Energy Rev. 2021, 141, 110784. [Google Scholar] [CrossRef]
- Hesami, M.; Pepe, M.; Baiton, A.; Salami, S.A.; Jones, A.M. New insight into ornamental applications of cannabis: Perspectives and challenges. Plants 2022, 11, 2383. [Google Scholar] [CrossRef]
- Adhikary, D.; Kulkarni, M.; El-Mezawy, A.; Mobini, S.; Elhiti, M.; Gjuric, R.; Ray, A.; Polowick, P.; Slaski, J.J.; Jones, M.P.; et al. Medical cannabis and industrial hemp tissue culture: Present status and future potential. Front. Plant Sci. 2021, 12, 627240. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.; Hesami, M.; Jones, A.M.P. The past, present and future of Cannabis sativa tissue culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef]
- Adamek, K.; Jones, A.M.P.; Torkamaneh, D. Accumulation of somatic mutations leads to genetic mosaicism in cannabis. Plant Genom. 2022, 15, e20169. [Google Scholar] [CrossRef]
- Pepe, M.; Hesami, M.; Small, F.; Jones, A.M.P. Comparative Analysis of Machine Learning and Evolutionary Optimization Algorithms for Precision Micropropagation of Cannabis sativa: Prediction and Validation of in vitro Shoot Growth and Development Based on the Optimization of Light and Carbohydrate Sources. Front. Plant Sci. 2021, 12, 757869. [Google Scholar] [CrossRef]
- Pepe, M.; Leonardos, E.D.; Marie, T.R.J.G.; Kyne, S.T.; Hesami, M.; Jones, A.M.; Grodzinski, B. A noninvasive gas exchange method to test and model photosynthetic proficiency and growth rates of in vitro plant cultures: Preliminary implication for Cannabis sativa L. Biology 2022, 11, 729. [Google Scholar] [CrossRef]
- Pepe, M.; Marie, T.R.J.G.; Leonardos, E.D.; Hesami, M.; Rana, N.; Jones, A.M.P.; Grodzinski, B. Tissue culture coupled with a gas exchange system offers new perspectives on phenotyping the developmental biology of Solanum lycopersicum L. cv. ‘MicroTom’. Front. Plant Sci. 2022, 13, 1025477. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Pepe, M.; Baiton, A.; Jones, A.M.P. Current status and future prospects in cannabinoid production through in vitro culture and synthetic biology. Biotechnol. Adv. 2023, 62, 108074. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, O.; Hesami, M.; Pepe, M.; Dutta, A.; Jones, A.M.P. In vitro plant tissue culture as the fifth generation of bioenergy. Sci. Rep. 2022, 12, 5038. [Google Scholar] [CrossRef]
- Downey, C.D.; Golenia, G.; Boudko, E.A.; Jones, A.M.P. Cryopreservation of 13 commercial Cannabis sativa genotypes using in vitro nodal explants. Plants 2021, 10, 1794. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Baiton, A.; Alizadeh, M.; Pepe, M.; Torkamaneh, D.; Jones, A.M. Advances and perspectives in tissue culture and genetic engineering of cannabis. Int. J. Mol. Sci. 2021, 22, 5671. [Google Scholar] [CrossRef]
- Kulus, D. Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. In Vitro Cell. Dev. Biol. Plant 2020, 56, 447–457. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; Klerk, G.-J.D. (Eds.) Plant Tissue Culture Procedure—Background. In Plant Propagation by Tissue Culture: Volume 1. The Background; Springer: Dordrecht, The Netherlands, 2008; pp. 1–28. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. In vitro manipulation and propagation of medicinal plants. Biotechnol. Adv. 2000, 18, 91–120. [Google Scholar] [CrossRef]
- Page, S.R.G.; Monthony, A.S.; Jones, A.M.P. DKW basal salts improve micropropagation and callogenesis compared with MS basal salts in multiple commercial cultivars of Cannabis sativa. Botany 2021, 99, 269–279. [Google Scholar] [CrossRef]
- Isah, T. Nodal segment explant type and preconditioning influence in vitro shoot morphogenesis in Ginkgo biloba L. Plant Physiol. Rep. 2020, 25, 74–86. [Google Scholar] [CrossRef]
- Martini, A.N.; Vlachou, G.; Papafotiou, M. Effect of explant origin and medium plant growth regulators on in vitro shoot proliferation and rooting of Salvia tomentosa, a native sage of the Northeastern Mediterranean Basin. Agronomy 2022, 12, 1889. [Google Scholar] [CrossRef]
- Marks, T.R.; Myers, P.E. Effect of explant location upon early culture development in vitro. J. Hortic. Sci. 1992, 67, 583–591. [Google Scholar] [CrossRef]
- Duta-Cornescu, G.; Constantin, N.; Pojoga, D.-M.; Nicuta, D.; Simon-Gruita, A. Somaclonal variation—Advantage or disadvantage in micropropagation of the medicinal plants. Int. J. Mol. Sci. 2023, 24, 838. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.F.; Rodríguez, R.; Cañal, M.J. Genomic DNA methylation–demethylation during aging and reinvigoration of Pinus radiata. Tree Physiol. 2002, 22, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Shen, C.H.; Lin, W.D.; Chu, H.A.; Huang, B.L.; Kuo, C.I.; Yeh, K.W.; Huang, L.C.; Chang, I.F. Small RNAs of Sequoia sempervirens during rejuvenation and phase change. Plant Biol. 2013, 15, 27–36. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, Y.; Li, Y. Plant rejuvenation: From phenotypes to mechanisms. Plant Cell Rep. 2020, 39, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, J.; Wu, T.; Cao, J. Shoot regeneration and the relationship between organogenic capacity and endogenous hormonal contents in pumpkin. Plant Cell Tissue Organ Cult. 2008, 93, 323–331. [Google Scholar] [CrossRef]
- Antonopoulou, C.; Dimassi, K.; Chatzissavvidis, C.; Papadakis, I.; Therios, I. The effect of explant type and nutrient medium on the in vitro proliferation of olive (Olea europaea L. ‘Chondrolia chalkidikis’). Acta Hortic. 2012, 949, 185–189. [Google Scholar] [CrossRef]
- Graner, E.M.; Calderan-Meneghetti, E.; Leone, G.F.; de Almeida, C.V.; de Almeida, M. Long-term in vitro culture affects phenotypic plasticity of Neoregelia johannis plants. Plant Cell Tissue Organ Cult. 2019, 137, 511–524. [Google Scholar] [CrossRef]
- de Souza, R.; Adams, C.R.; de Melo, R.C.; Guidolin, A.F.; Michel, A.; Coimbra, J.L.M. Phenotypical changes in hop induced by micropropagation. Plant Cell Tissue Organ Cult. 2021, 147, 379–387. [Google Scholar] [CrossRef]
- Żebrowska, J.I. Effect of quantitative plant traits on the efficiency of in vitro cloning of strawberry (Fragaria × ananassa Duch.). J. Hortic. Sci. Biotechnol. 2015, 90, 407–412. [Google Scholar] [CrossRef]
- Cabral, G.B.; Carneiro, V.T.C.; Lacerda, A.L.; do Valle, C.B.; Martinelli, A.P.; Dusi, D.M.A. Somatic embryogenesis and organogenesis in apomictic and sexual Brachiaria brizantha. Plant Cell Tissue Organ Cult. 2011, 107, 271–282. [Google Scholar] [CrossRef]
- Driver, J.A.; Kuniyuki, A.H. In vitro propagation of paradox walnut rootstock. HortScience 1984, 19, 507–509. [Google Scholar] [CrossRef]
- Mestinšek-Mubi, Š.; Svetik, S.; Flajšman, M.; Murovec, J. In vitro tissue culture and genetic analysis of two high-CBD medical cannabis (Cannabis sativa L.) breeding lines. Genetika 2020, 52, 925–941. [Google Scholar] [CrossRef]
- Isah, T.; Zafar, N. In vitro rejuvenation of nodal segment explants for clonal propagation of Rauvolfia serpentina (L) Benth ex Kurz. Res. Sq. 2022, 202, 1957127. [Google Scholar] [CrossRef]
- Puddephat, I.J.; Alderson, P.G.; Wright, N.A. Influence of explant source, plant growth regulators and culture environment on culture initiation and establishment of Quercus robur L. in vitro. J. Exp. Bot. 1997, 48, 951–962. [Google Scholar] [CrossRef]
- Zulfiqar, B.; Abbasi, N.A.; Ahmad, T.; Hafiz, I.A. Effect of explant sources and different concentrations of plant growth regulators on in vitro shoot proliferation and rooting of avocado (Persea americana Mill.) cv.“Fuerte”. Pak. J. Bot. 2009, 41, 2333–2346. [Google Scholar]
- Mora, L.Y.C.; Tarazona, D.Y.G.; Bohórquez Quintero, M.d.l.A.; Barrera, E.J.A.; Ruíz, J.S.U.; Moreno, D.M.A.; Pérez, Z.Z.O. Impact of initial explants on in vitro propagation of native potato (Solanum tuberosum, Andigena group). Plant Cell Tissue Organ Cult. 2022, 150, 627–636. [Google Scholar] [CrossRef]
- de Aguiar, T.C.F.; Negri, A.P.; Boff, P.; Dalla Costa, M.; Boff, M.I.C.; RombolÃ, A.D. Effects of explant position and orientation, medium pH and nitrogen sources on micropropagation of blackberry. Ibero-Am. J. Environ. Sci. 2021, 12, 84–92. [Google Scholar] [CrossRef]
- Aruna, V.; Kiranmai, C.; Karuppusamy, S.; Pullaiah, T. Effect of medium, explants, cytokinins and node position on in vitro shoot multiplication of Caralluma lasiantha (Wight) NE Br., an endemic and medicinally important plant. Afr. J. Biotechnol. 2012, 11, 15523–15528. [Google Scholar] [CrossRef]
- Shekafandeh, A.; Khosh-Khui, M. Effects of bud position and culture medium on shoot proliferation from nodal culture of two mature guava cultivars. Asian J. Plant Sci. 2008, 7, 177–182. [Google Scholar] [CrossRef]
- Sreekumar, S.; Seeni, S.; Pushpangadan, P. Micropropagation of Hemidesmus indicus for cultivation and production of 2-hydroxy 4-methoxy benzaldehyde. Plant Cell Tissue Organ Cult. 2000, 62, 211–218. [Google Scholar] [CrossRef]
- Hesami, M.; Daneshvar, M.H.; Lotfi, A. In vitro shoot proliferation through cotyledonary node and shoot tip explants of Ficus religiosa L. Plant Tissue Cult. Biotechnol. 2017, 27, 85–88. [Google Scholar] [CrossRef]
- Basto, S.; Serrano, C.; Hodson de Jaramillo, E. Effects of donor plant age and explants on in vitro culture of Cedrela montana Moritz ex Turcz. Univ. Sci. 2012, 17, 263–271. [Google Scholar] [CrossRef]
- Shukla, M.R.; Jones, A.M.P.; Sullivan, J.A.; Liu, C.; Gosling, S.; Saxena, P.K. In vitro conservation of American elm (Ulmus americana): Potential role of auxin metabolism in sustained plant proliferation. Can. J. For. Res. 2012, 42, 686–697. [Google Scholar] [CrossRef]
- Miguel, C.; Marum, L. An epigenetic view of plant cells cultured in vitro: Somaclonal variation and beyond. J. Exp. Bot. 2011, 62, 3713–3725. [Google Scholar] [CrossRef] [PubMed]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Adly, W.M.R.M.; Niedbała, G.; El-Denary, M.E.; Mohamed, M.A.; Piekutowska, M.; Wojciechowski, T.; Abd El-Salam, E.-S.T.; Fouad, A.S. Somaclonal Variation for Genetic Improvement of Starch Accumulation in Potato (Solanum tuberosum) Tubers. Plants 2023, 12, 232. [Google Scholar] [CrossRef]
- Pastelín Solano, M.C.; Salinas Ruíz, J.; González Arnao, M.T.; Castañeda Castro, O.; Galindo Tovar, M.E.; Bello Bello, J.J. Evaluation of in vitro shoot multiplication and ISSR marker based assessment of somaclonal variants at different subcultures of vanilla (Vanilla planifolia Jacks). Physiol. Mol. Biol. Plants 2019, 25, 561–567. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, T.-T.; Zhang, J. Effect of subculture times on genetic fidelity, endogenous hormone level and pharmaceutical potential of Tetrastigma hemsleyanum callus. Plant Cell Tissue Organ Cult. 2015, 122, 67–77. [Google Scholar] [CrossRef]
- Khan, S.; Saeed, B.; Kauser, N. Establishment of genetic fidelity of in vitro raised banana plantlets. Pak. J. Bot. 2011, 43, 233–242. [Google Scholar]
- Farahani, F.; Yari, R.; Masoud, S. Somaclonal variation in Dezful cultivar of olive (Olea europaea subsp. europaea). Gene Conserve 2011, 40, 216–233. [Google Scholar]
- Clarindo, W.R.; Carvalho, C.R.; Mendonça, M.A.C. Ploidy instability in long-term in vitro cultures of Coffea arabica L. monitored by flow cytometry. Plant Growth Regul. 2012, 68, 533–538. [Google Scholar] [CrossRef]
- Rival, A.; Ilbert, P.; Labeyrie, A.; Torres, E.; Doulbeau, S.; Personne, A.; Dussert, S.; Beulé, T.; Durand-Gasselin, T.; Tregear, J.W.; et al. Variations in genomic DNA methylation during the long-term in vitro proliferation of oil palm embryogenic suspension cultures. Plant Cell Rep. 2013, 32, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Igamberdiev, A.U.; Debnath, S.C. Tissue culture-induced DNA methylation in crop plants: A review. Mol. Biol. Rep. 2021, 48, 823–841. [Google Scholar] [CrossRef] [PubMed]
- Gulyás, A.; Dobránszki, J.; Kiss, E.; da Silva, J.A.T.; Posta, K.; Hidvégi, N. Changes in DNA methylation pattern of apple long-term in vitro shoot culture and acclimatized plants. J. Plant Physiol. 2019, 239, 18–27. [Google Scholar] [CrossRef]
- Borges, F.; Donoghue, M.T.A.; LeBlanc, C.; Wear, E.E.; Tanurdžić, M.; Berube, B.; Brooks, A.; Thompson, W.F.; Hanley-Bowdoin, L.; Martienssen, R.A. Loss of Small-RNA-Directed DNA Methylation in the Plant Cell Cycle Promotes Germline Reprogramming and Somaclonal Variation. Curr. Biol. 2021, 31, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.; Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Morpho-Anatomy of Marijuana (Cannabis sativa L.). In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 123–136. [Google Scholar] [CrossRef]
- Spitzer-Rimon, B.; Duchin, S.; Bernstein, N.; Kamenetsky, R. Architecture and Florogenesis in Female Cannabis sativa Plants. Front. Plant Sci. 2019, 10, 350. [Google Scholar] [CrossRef]
- Spitzer-Rimon, B.; Shafran-Tomer, H.; Gottlieb, G.H.; Doron-Faigenboim, A.; Zemach, H.; Kamenetsky-Goldstein, R.; Flaishman, M. Non-photoperiodic transition of female cannabis seedlings from juvenile to adult reproductive stage. Plant Reprod. 2022, 35, 265–277. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Potential roles of epigenetic memory on the quality of clonal cannabis plants: Content and profile of secondary metabolites. Med. Usage Cannabis Cannabinoids 2023, 1, 1–14. [Google Scholar]
- Bastías, A.; Almada, R.; Rojas, P.; Donoso, J.M.; Hinrichsen, P.; Sagredo, B. Aging gene pathway of microRNAs 156/157 and 172 is altered in juvenile and adult plants from in vitro propagated Prunus sp. Int. J. Agric. Nat. Resour. 2016, 43, 429–441. [Google Scholar] [CrossRef]
- Ashapkin, V.V.; Kutueva, L.I.; Vanyushin, B.F. Aging epigenetics: Accumulation of errors or realization of a specific program? Biochem. Mosc. 2015, 80, 1406–1417. [Google Scholar] [CrossRef] [PubMed]
- Cierjacks, A.; Kowarik, I.; Joshi, J.; Hempel, S.; Ristow, M.; von der Lippe, M.; Weber, E. Biological Flora of the British Isles: Robinia pseudoacacia. J. Ecol. 2013, 101, 1623–1640. [Google Scholar] [CrossRef]
- Masaka, K.; Torita, H.; Kon, H.; Fukuchi, M. Seasonality of sprouting in the exotic tree Robinia pseudoacacia L. in Hokkaido, northern Japan. J. For. Res. 2015, 20, 386–395. [Google Scholar] [CrossRef]
- Ye, B.-B.; Zhang, K.; Wang, J.-W. The role of miR156 in rejuvenation in Arabidopsis thaliana. J. Integr. Plant Biol. 2020, 62, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.H.; Lineberger, R.D. In vitro rejuvenation of Betula (Betulaceae): Morphological evaluation. Am. J. Bot. 1992, 79, 618–625. [Google Scholar] [CrossRef]
- Monteuuis, O. Rejuvenation of a 100-year-old Sequoiadendron giganteum through in vitro meristem culture. I. Organogenic and morphological arguments. Physiol. Plant. 1991, 81, 111–115. [Google Scholar] [CrossRef]
- Lu, Z.; Zhu, L.; Lu, J.; Shen, N.; Wang, L.; Liu, S.; Wang, Q.; Yu, W.; Kato-Noguchi, H.; Li, W.; et al. Rejuvenation increases leaf biomass and flavonoid accumulation in Ginkgo biloba. Hortic. Res. 2022, 9, uhab018. [Google Scholar] [CrossRef]
- Popov, V.N.; Syromyatnikov, M.Y.; Franceschi, C.; Moskalev, A.A.; Krutovsky, K.V. Genetic mechanisms of aging in plants: What can we learn from them? Ageing Res. Rev. 2022, 77, 101601. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subculture | Explant’s Source | PH ± SE (mm) | NN ± SE | NS ± SE | CSA ± SE (mm2) |
|---|---|---|---|---|---|
| I | A | 43.45 ± 1.113 | 8.5 ± 0.501 | 1 ± 0.000 | 994 ± 31.093 |
| I | B | 45.335 ± 2.192 | 8.5 ± 0.501 | 1 ± 0.000 | 1004 ± 5.015 |
| I | C | 43.88 ± 2.648 | 8.5 ± 1.504 | 1 ± 0.000 | 1296 ± 60.179 |
| I | D | 38.25 ± 2.848 | 7.5 ± 0.501 | 1 ± 0.000 | 1119.5 ± 45.636 |
| II | A | 49.71 ± 1.525 | 8.5 ± 0.501 | 1 ± 0.000 | 1927.3 ± 74.221 |
| II | B | 43.995 ± 2.533 | 8.5 ± 1.504 | 1.5 ± 0.501 | 2293.5 ± 57.672 |
| II | C | 41.21 ± 2.046 | 7.5 ± 0.501 | 1 ± 0.000 | 1392 ± 67.200 |
| II | D | 35.475 ± 2.071 | 6 ± 1.003 | 1 ± 0.000 | 1025 ± 129.385 |
| III | A | 39.33 ± 0.923 | 7.5 ± 0.501 | 1 ± 0.000 | 1124.5 ± 58.675 |
| III | B | 41.005 ± 1.439 | 8 ± 1.003 | 1.5 ± 0.501 | 1388 ± 19.057 |
| III | C | 30.36 ± 1.033 | 5 ± 1.003 | 1 ± 0.000 | 889.5 ± 15.546 |
| III | D | 29.22 ± 0.802 | 3.5 ± 0.501 | 1 ± 0.000 | 803 ± 22.066 |
| IV | A | 36.865 ± 2.553 | 7 ± 1.003 | 1 ± 0.000 | 1182.5 ± 46.639 |
| IV | B | 36.54 ± 1.113 | 6.5 ± 1.504 | 1 ± 0.000 | 968.5 ± 37.612 |
| IV | C | 33.075 ± 1.449 | 4 ± 1.003 | 1 ± 0.000 | 957 ± 21.063 |
| IV | D | 28.995 ± 0.707 | 3.5 ± 0.501 | 1 ± 0.000 | 967.5 ± 46.639 |
| p value | |||||
| Explant | <0.0001 | 0.0010 | 0.5847 | <0.0001 | |
| Subculture | <0.0001 | 0.0014 | 0.1546 | <0.0001 | |
| Explant × subculture | 0.1400 | 0.6394 | 0.7271 | <0.0001 | |
| Subculture | Explant’s Source | NLl ± SE | LCLl ± SE (mm) | WCLl ± SE (mm) | NPS ± SE |
|---|---|---|---|---|---|
| I | A | 5 ± 0.000 | 44.20 ± 1.249 | 6.36 ± 0.375 | 18.4 ± 0.400 |
| I | B | 5 ± 0.000 | 47.30 ± 0.870 | 5.70 ± 0.329 | 21.6 ± 0.748 |
| I | C | 5 ± 0.000 | 49.53 ± 2.249 | 6.67 ± 0.260 | 22.8 ± 1.020 |
| I | D | 5 ± 0.000 | 58.81 ± 0.695 | 5.06 ± 0.151 | 19.2 ± 0.800 |
| II | A | 3 ± 0.000 | 40.66 ± 1.373 | 5.68 ± 0.212 | 20.8 ± 0.490 |
| II | B | 3 ± 0.000 | 40.76 ± 2.141 | 4.89 ± 0.178 | 21.2 ± 0.800 |
| II | C | 3 ± 0.000 | 42.19 ± 0.527 | 4.54 ± 0.149 | 17.2 ± 0.490 |
| II | D | 3 ± 0.000 | 45.94 ± 1.172 | 7.58 ± 0.296 | 19.2 ± 0.490 |
| III | A | 3 ± 0.000 | 33.37 ± 1.741 | 9.99 ± 0.297 | 20.8 ± 0.490 |
| III | B | 3 ± 0.000 | 34.60 ± 1.001 | 5.11 ± 0.330 | 16.4 ± 0.400 |
| III | C | 3 ± 0.000 | 40.85 ± 0.763 | 5.05 ± 0.097 | 20.8 ± 0.490 |
| III | D | 3 ± 0.000 | 42.27 ± 0.660 | 5.43 ± 0.328 | 18.8 ± 0.490 |
| IV | A | 3 ± 0.000 | 27.07 ± 1.339 | 8.57 ± 0.179 | 18.4 ± 0.400 |
| IV | B | 3 ± 0.000 | 35.30 ± 0.664 | 9.85 ± 0.303 | 18 ± 0.000 |
| IV | C | 3 ± 0.000 | 39.25 ± 1.242 | 11.60 ± 0.604 | 18 ± 0.000 |
| IV | D | 3 ± 0.000 | 40.44 ± 0.787 | 5.12 ± 0.283 | 19.2 ± 0.490 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hesami, M.; Adamek, K.; Pepe, M.; Jones, A.M.P. Effect of Explant Source on Phenotypic Changes of In Vitro Grown Cannabis Plantlets over Multiple Subcultures. Biology 2023, 12, 443. https://doi.org/10.3390/biology12030443
Hesami M, Adamek K, Pepe M, Jones AMP. Effect of Explant Source on Phenotypic Changes of In Vitro Grown Cannabis Plantlets over Multiple Subcultures. Biology. 2023; 12(3):443. https://doi.org/10.3390/biology12030443
Chicago/Turabian StyleHesami, Mohsen, Kristian Adamek, Marco Pepe, and Andrew Maxwell Phineas Jones. 2023. "Effect of Explant Source on Phenotypic Changes of In Vitro Grown Cannabis Plantlets over Multiple Subcultures" Biology 12, no. 3: 443. https://doi.org/10.3390/biology12030443
APA StyleHesami, M., Adamek, K., Pepe, M., & Jones, A. M. P. (2023). Effect of Explant Source on Phenotypic Changes of In Vitro Grown Cannabis Plantlets over Multiple Subcultures. Biology, 12(3), 443. https://doi.org/10.3390/biology12030443