
Effects of Hypoxia Stress on Survival, Antioxidant and Anaerobic Metabolic Enzymes, and Related Gene Expression of Red Swamp Crayfish Procambarus clarkii



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Ethics
2.2. Oxygen Consumption Rate and Asphyxiation Point Experiment
2.3. Hypoxia Stress Experiment
2.4. Sample Collection
2.5. Enzyme Activity Assay
2.6. RNA Isolation and Quantitative RT–PCR
2.7. Data Analysis
3. Results
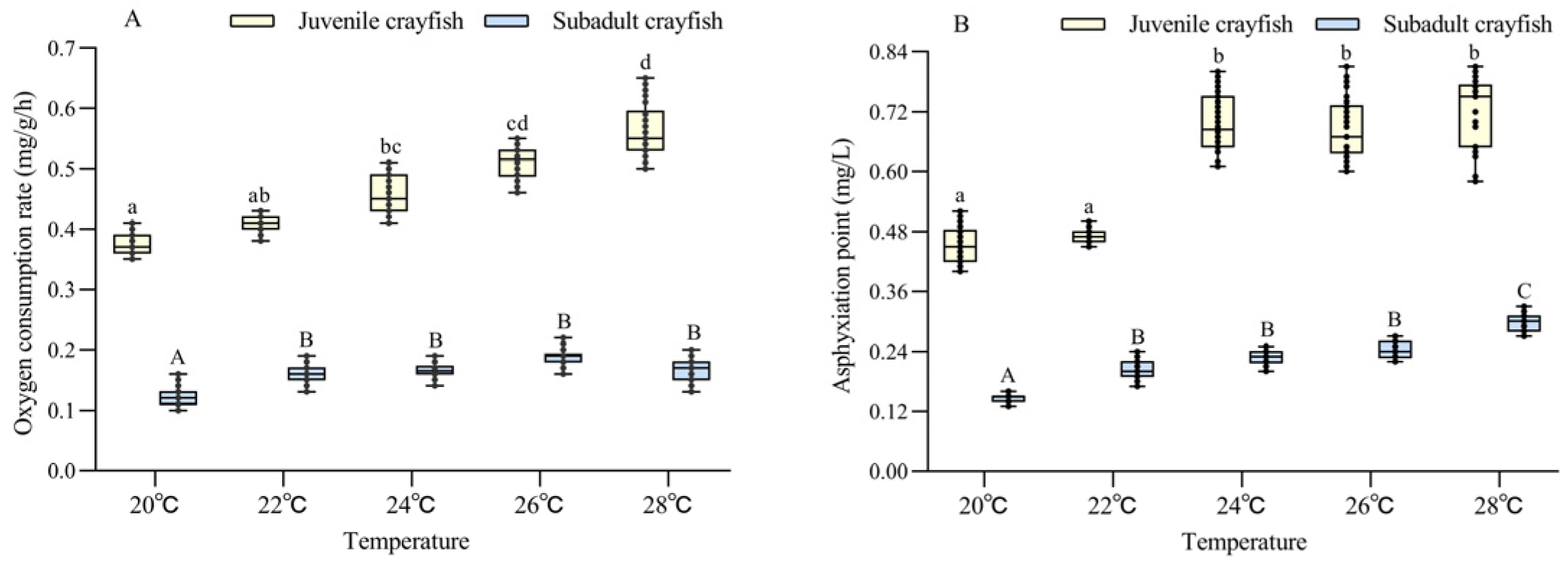
3.1. Oxygen Consumption Rate, Asphyxiation Point, and Mortality
3.2. Activities of Antioxidant Enzymes and Anaerobic Metabolic Enzymes in Juvenile P. clarkii
3.3. Activities of Antioxidant Enzymes and Anaerobic Metabolic Enzymes in Subadult P. clarkii
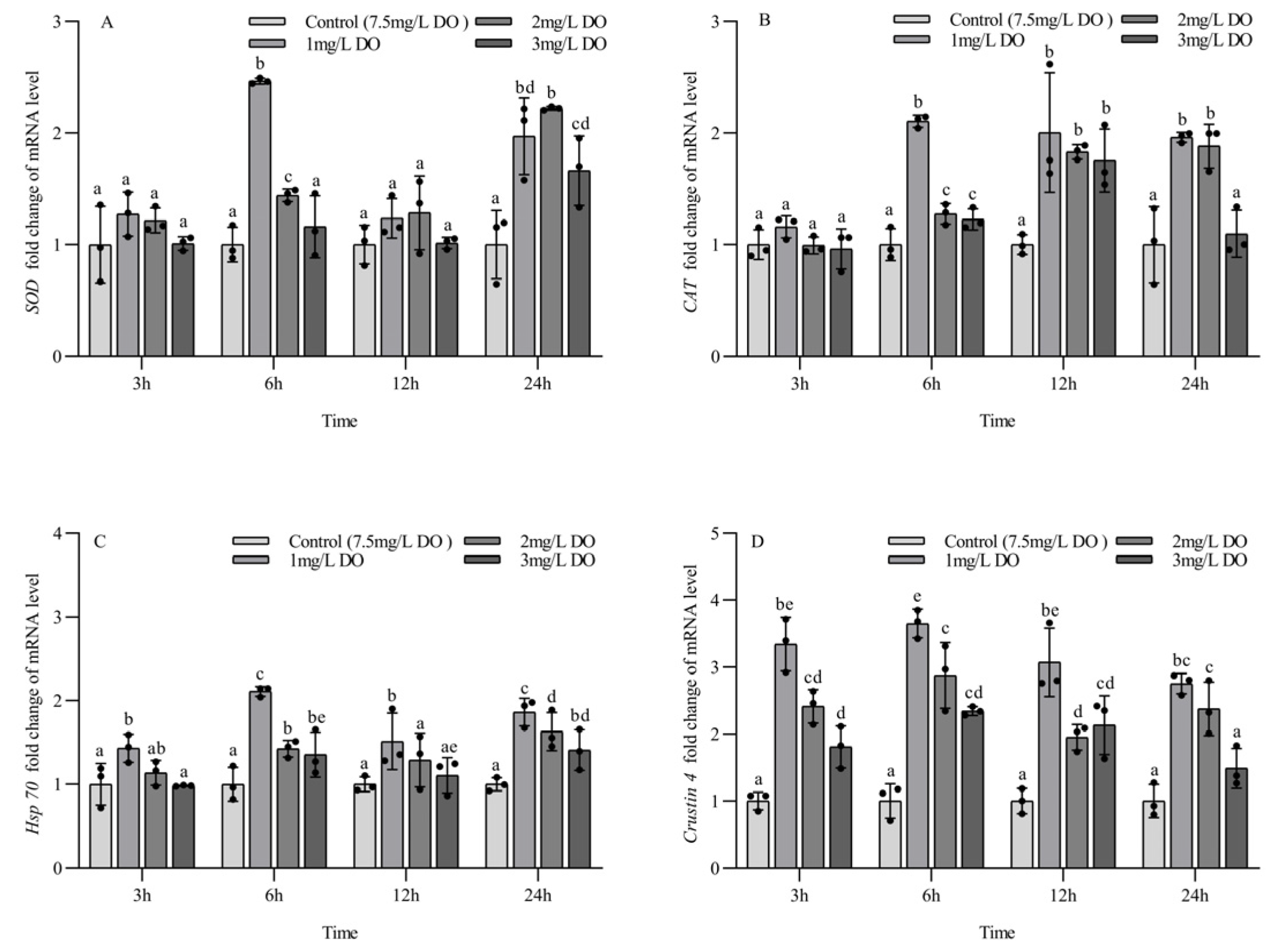
3.4. Antioxidant and Stress Gene Expression of Juvenile P. clarkii
3.5. Antioxidant and Stress Gene Expression of Subadult P. clarkii
4. Discussion
4.1. Effect of Temperature on the Oxygen Consumption Rate and Asphyxiation Point of P. clarkii
4.2. Effect of Hypoxia on the Activity of Antioxidant and Anaerobic Metabolic Enzymes in P. clarkii
4.3. Effect of Hypoxia on the Expression of Antioxidant and Stress Genes in P. clarkii
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shu, X.Y. Problems and industrial development of Procambarus clarkii. China Fish. 2010, 08, 22–25. (In Chinese) [Google Scholar] [CrossRef]
- Guo, L.; Lin, W.; Cao, C.; Li, C. Integrated rice-crayfish farming system does not mitigate the global warming potential during rice season. Sci. Total Environ. 2023, 867, 161520. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Li, X.; Onchari, M.M.; Li, W.; Bu, Y.; Lek, S.; Jin, S. High feeding level alters physiological status but does not improve feed conversion efficiency and growth performance of juvenile red swamp crayfish Procambarus clarkii (Girard, 1852). Aquaculture 2021, 537, 736507. [Google Scholar] [CrossRef]
- Yu, X.J.; Hao, X.J.; Dang, Z.Q.; Yang, L.K.; Zhang, Y.H.; Cai, J.; Zhang, Y. China crayfish industry development report. China Fish. 2022, 6, 47–54. (In Chinese) [Google Scholar] [CrossRef]
- Liu, D.L.; Huang, J.H.; Yang, L.S.; Tan, L.Q. Effects of multiple environmental factors on molting death of Procambarus clarkii and countermeasures. Nanfang Shuichan Kexue 2020, 2, 29–35. (In Chinese) [Google Scholar] [CrossRef]
- Shu, B.; Li, J.T.; Zhang, X.X.; Xian, J.A.; Zhou, X.Y.; Wang, D.M. A Review: Effects of Environmental Factors on Immune Indices of Red Swamp Crayfish Procambarus clarkii. Shuichanxue Zazhi 2020, 33, 75. (In Chinese) [Google Scholar] [CrossRef]
- Estrada-Cárdenas, P.; Cruz-Moreno, D.G.; González-Ruiz, R.; Peregrino-Uriarte, A.B.; Leyva-Carrillo, L.; Camacho-Jiménez, L.; Yepiz-Plascencia, G. Combined hypoxia and high temperature affect differentially the response of antioxidant enzymes, glutathione and hydrogen peroxide in the white shrimp Litopenaeus vannamei. Comp. Biochem. Phys. A 2021, 254, 110909. [Google Scholar] [CrossRef] [PubMed]
- Hadley, K.R.; Paterson, A.M.; Reid, R.A.; Rusak, J.A.; Somers, K.M.; Ingram, R.; Smol, J.P. Altered pH and reduced calcium levels drive near extirpation of native crayfish, Cambarus bartonii, in Algonquin Park, Ontario, Canada. Freshw. Sci. 2015, 34, 918–932. [Google Scholar] [CrossRef]
- Jiang, L.X.; Pan, L.Q.; Xiao, G.Q. Effects of ammonia-N on immune parameters of white shrimp Litopenaeus vannamei. Zhongguo Shuichan Kexue 2004, 11, 537–541. (In Chinese) [Google Scholar] [CrossRef]
- Wang, P.; Liu, H.; Zhao, S.; Yu, S.; Xie, S.; Hua, S.; Gao, H. Hypoxia stress affects the physiological responses, apoptosis and innate immunity of Kuruma shrimp, Marsupenaeus japonicus. Fish. Shellfish. Immunol. 2022, 122, 206–214. [Google Scholar] [CrossRef]
- Liu, Q.J. Effects of Straw Return and Food Feeding on Water Quality in Rice Crawfish System. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- France, R.L. Comparative tolerance to low pH of three life stages of the crayfish Orconectes virilis. Can. J. Zool. 1984, 62, 2360–2363. [Google Scholar] [CrossRef]
- Zhong, J.W.; Zhu, Y.A.; Fu, P.S.; An, L.; Lv, B.Y. Toxicity of nitrite nitrogen on Procambarus clarkii and effects of nitrite nitrogen on immune-related factor. J. Yangtze Univ. (Mat. Sci. Ed.) 2014, 11, 36–40. (In Chinese) [Google Scholar] [CrossRef]
- Camacho-Jiménez, L.; Peregrino-Uriarte, A.B.; Martínez-Quintana, J.A.; Yepiz-Plascencia, G. The glyceraldehyde-3-phosphate dehydrogenase of the shrimp Litopenaeus vannamei: Molecular cloning, characterization and expression during hypoxia. Mar. Environ. Res. 2018, 138, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Zou, E.M.; Du, N.S.; Lai, W. The effects of mass temperature and thermal acclimation on the respiration rate of the Chinese freshwater crab Eriocheir sinensis (Crustacea: Decapod). Zool. Res. 1995, 16, 49–58. [Google Scholar] [CrossRef]
- Giordano, F.J. Oxygen, oxidative stress, hypoxia, and heart failure. J. Clin. Investig. 2005, 115, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Krishnan, S.; Anand, D.; Kokkattunivarthil Uthaman, S.; Otta, S.K.; Karunasagar, I.; Kooloth Valappil, R. Immune responses and immune protection in crustaceans with special reference to shrimp. Rev. Aquacult. 2021, 13, 431–459. [Google Scholar] [CrossRef]
- Norouzitallab, P.; Baruah, K.; Vanrompay, D.; Bossier, P. Teaching shrimps self-defense to fight infections. Trends Biotechnol. 2019, 37, 16–19. [Google Scholar] [CrossRef]
- Wilhelm Filho, D.; Torres, M.A.; Zaniboni-Filho, E.; Pedrosa, R.C. Effect of different oxygen tensions on weight gain, feed conversion, and antioxidant status in piapara, Leporinus elongatus (Valenciennes, 1847). Aquaculture 2005, 244, 349–357. [Google Scholar] [CrossRef]
- Han, S.Y.; Wang, M.Q.; Liu, M.; Wang, B.J.; Jiang, K.Y.; Wang, L. Comparative sensitivity of the hepatopancreas and midgut in the white shrimp Litopenaeus vannamei to oxidative stress under cyclic serious/medium hypoxia. Aquaculture 2018, 490, 44–52. [Google Scholar] [CrossRef]
- Sun, S.; Yang, M.; Fu, H.; Ge, X.; Zou, J. Altered intestinal microbiota induced by chronic hypoxia drives the effects on lipid metabolism and the immune response of oriental river prawn Macrobrachium nipponense. Aquaculture 2020, 526, 735431. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, X.; Li, J.; Huang, G. The effects of body weight, temperature, salinity, pH, light intensity and feeding condition on lethal DO levels of whiteleg shrimp, Litopenaeus vannamei (Boone, 1931). Aquaculture 2006, 256, 579–587. [Google Scholar] [CrossRef]
- Wen, X.B.; Chen, L.Q.; Ai, C.X.; Jiang, H.B. Standard Metabolism of the juvenile Crab Eriocheir sicensis. Zool. Res. 2001, 22, 425–428. (In Chinese) [Google Scholar] [CrossRef]
- Chen, Q.; Chen, X.H.; Luo, Y.J.; Huang, J.; Li, G.F.; Ning, L.K. Preliminary determination of oxygen consumption rate and asphyxia point of Litopenaeus vannamei. J. Hydrol. 2001, 21, 14–15. (In Chinese) [Google Scholar] [CrossRef]
- Nelson, S.G.; Armstrong, D.A.; Knight, A.W.; Li, H.W. The effects of temperature and salinity on the metabolic rate of juvenile Macrobrachium rosenbergii (Crustacea: Palaemonidae). Comp. Biochem. Phys. A 1977, 56, 533–537. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Liu, D.; Ru, S. Cu accumulation, detoxification and tolerance in the red swamp crayfish Procambarus clarkii. Ecotoxicol. Environ. Saf. 2019, 175, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Hammen, C.S. Aminotransferase activities and amino acid excretion of bivalve mollusks and brachiopods. Comp. Biochem. Phys. B 1968, 26, 697–705. [Google Scholar] [CrossRef]
- Beaufort, N.C. Biology of the blue crab, Callinectes sapidus rathbun, In the St. Johns River, Florida. Fish. B-Noaa. 1968, 67, 17–33. [Google Scholar]
- Christensen, E.A.; Svendsen, M.B.; Steffensen, J.F. The combined effect of body size and temperature on oxygen consumption rates and the size-dependency of preferred temperature in European perch Perca fluviatilis. J. Fish. Biol. 2020, 97, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Feng, W.R.; Zhou, L.H.; Ding, X.F.; Liang, M.; Tang, Y.K. Effects of temperature and pH on the oxygen consumption rate, ammonia discharge rate and asphyxiation point of juvenile Eriocheir sinensis. J. South. Agric. 2022, 05, 1415–1424. (In Chinese) [Google Scholar] [CrossRef]
- Dai, Q.N.; Wang, J.; Li, Q. On the oxygen consumption of Penaeus japouicus. Jimei Daxue Xuebao Ziran Kexueban 1994, 16, 25–29. (In Chinese) [Google Scholar] [CrossRef]
- Franco, R.; Sánchez-Olea, R.; Reyes-Reyes, E.M.; Panayiotidis, M.I. Environmental toxicity, oxidative stress and apoptosis: Menage a trois. Mutat.Res. Genet. Toxicol. Environ. Mutagen. 2009, 674, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, Y.M.; Xu, W.B.; Chen, D.Y.; Li, B.W.; Cheng, Y.X.; Shu, M.A. The effects of salinities stress on histopathological changes, serum biochemical index, non-specific immune and transcriptome analysis in red swamp crayfish Procambarus clarkii. Sci. Total Environ. 2022, 840, 156502. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Jiang, Q. The effect of a polystyrene nanoplastic on the intestinal microbes and oxidative stress defense of the freshwater crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Reiber, C.L.; McMahon, B.R. The effects of progressive hypoxia on the crustacean cardiovascular system: A comparison of the freshwater crayfish, (Procambarus clarkii), and the lobster (Homarus americanus). J. Comp. Physiol. B 1998, 168, 168–176. [Google Scholar] [CrossRef]
- Fujimori, T.; Abe, H. Physiological roles of free D-and L-alanine in the crayfish Procambarus clarkii with special reference to osmotic and anoxic stress responses. Comp. Biochem. Phys. A 2002, 131, 893–900. [Google Scholar] [CrossRef]
- da Silva-Castiglioni, D.; Oliveira, G.T.; Buckup, L. Metabolic responses of Parastacus defossus and Parastacus brasiliensis (Crustacea, Decapoda, Parastacidae) to hypoxia. Comp. Biochem. Phys. A 2010, 156, 436–444. [Google Scholar] [CrossRef] [PubMed]
- McMahon, B.R. Respiratory and circulatory compensation to hypoxia in crustaceans. Respir. Physiol. 2001, 128, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Truchot, J.P. Lactate increases the oxygen affinity of crab hemocyanin. J. Exp. Zool. 1980, 214, 205–208. [Google Scholar] [CrossRef]
- Qiu, R.; Cheng, Y.; Huang, X.; Wu, X.; Yang, X.; Tong, R. Effect of hypoxia on immunological, physiological response, and hepatopancreatic metabolism of juvenile Chinese mitten crab Eriocheir sinensis. Aquac. Int. 2011, 19, 283–299. [Google Scholar] [CrossRef]
- Qiu, D.G.; Huang, D.C.; Zheng, L.Y.; Wu, S.Q.; Liu, Y.H.; Qiu, F.Y.; Zheng, J.P.; Chen, X.M. Effects of temperature on respiration and excretion by different sizes of hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceo latus ♂). Zhongguo Shuichan kexue 2017, 24, 295–305. (In Chinese) [Google Scholar] [CrossRef]
- Eyckmans, M.; Celis, N.; Horemans, N.; Blust, R.; De Boeck, G. Exposure to waterborne copper reveals differences in oxidative stress response in three freshwater fish species. Aquat. Toxicol. 2011, 103, 112–120. [Google Scholar] [CrossRef]
- Benjamin, I.J. 20 Expression and function of stress proteins in the ischemic heart. CSH Monogr. 1994, 26, 533–552. [Google Scholar]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Basu, S.; Binder, R.J.; Suto, R.; Anderson, K.M.; Srivastava, P.K. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-κB pathway. Int. Immunol. 2000, 12, 1539–1546. [Google Scholar] [CrossRef]
- Zhu, Y.T. The Anti-Microbial Functions of Immune Effectors in the Hemocytes of Chinese Mitten Crab, Eriocheir sinensis. Ph.D. Thesis, East China Normal University, Shanghai, China, 2016. [Google Scholar]
- Du, Z.Q.; Li, B.; Shen, X.L.; Wang, K.; Du, J.; Yu, X.D.; Yuan, J.J. A new antimicrobial peptide isoform, Pc-crustin 4 involved in antibacterial innate immune response in fresh water crayfish, Procambarus clarkii. Fish. Shellfish. Immunol. 2019, 94, 861–870. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence(5′-3′) | GenBank Accession No. |
|---|---|---|
| 18S-F | GTCAGGTCATCACCATCGGCA | HQ-414542.1 |
| 18S-R | CGGTCTCGTGAACACCAGCA | |
| SOD-F | CGCCGATGTAAGACTGGGACG | TRINITY-DN63424-c1-g1 |
| SOD-R | CTCCAGGTAAACACGGCTTCCAT | |
| CAT-F | TCCTGTGAACTGTCCCTATCGTG | TRINITY-DN27834-c0-g1 |
| CAT-R | AACCCAGTCTTCTTACAATCAACG | |
| Crustin-F | CTCTGACTGCCAGGTGTTT | NW-020872843.1 |
| Crustin-R | TGCGAGCTGTGATGGTTAG | |
| Hsp70-F | GTTGACCAAGATGAAGGAGAC | DQ-301506.1 |
| Hsp70-R | CTGACGCTGAGAGTCGTTG |
| Variable of Treatment | F Value | df | p Value |
|---|---|---|---|
| Oxygen consumption rate | |||
| Temperature | 13.26 | 4 | <0.001 |
| Size | 656.42 | 1 | <0.001 |
| Temperature × Size | 5.17 | 4 | 0.005 |
| Asphyxiation point | |||
| Temperature | 11.93 | 4 | <0.001 |
| Size | 260.70 | 1 | <0.001 |
| Temperature × Size | 2.71 | 4 | 0.059 |
| Variable of Treatment | F Value | df | p Value |
|---|---|---|---|
| SOD | |||
| Dissolved oxygen | 36.62 | 3 | <0.001 |
| Sampling time | 28.13 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 7.61 | 9 | <0.001 |
| CAT | |||
| Dissolved oxygen | 43.53 | 3 | <0.001 |
| Sampling time | 1.85 | 3 | 0.157 |
| Dissolved oxygen × Sampling time | 6.54 | 9 | <0.001 |
| LD | |||
| Dissolved oxygen | 27.41 | 3 | <0.001 |
| Sampling time | 139.05 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 12.52 | 9 | <0.001 |
| LDH | |||
| Dissolved oxygen | 88.67 | 3 | <0.001 |
| Sampling time | 36.69 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 12.78 | 9 | <0.001 |
| Variable of Treatment | F Value | df | p Value |
|---|---|---|---|
| SOD | |||
| Dissolved oxygen | 24.08 | 3 | <0.001 |
| Sampling time | 6.30 | 3 | 0.002 |
| Dissolved oxygen × Sampling time | 10.19 | 9 | <0.001 |
| CAT | |||
| Dissolved oxygen | 109.87 | 3 | <0.001 |
| Sampling time | 13.51 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 15.15 | 9 | <0.001 |
| LD | |||
| Dissolved oxygen | 55.17 | 3 | <0.001 |
| Sampling time | 7.46 | 3 | 0.001 |
| Dissolved oxygen × Sampling time | 8.49 | 9 | <0.001 |
| LDH | |||
| Dissolved oxygen | 109.85 | 3 | <0.001 |
| Sampling time | 12.91 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 10.41 | 9 | <0.001 |
| Variable of Treatment | F Value | df | p Value |
|---|---|---|---|
| SOD mRNA | |||
| Dissolved oxygen | 10.08 | 3 | <0.001 |
| Sampling time | 3.75 | 3 | 0.020 |
| Dissolved oxygen × Sampling time | 4.35 | 9 | 0.001 |
| CAT mRNA | |||
| Dissolved oxygen | 30.10 | 3 | <0.001 |
| Sampling time | 2.63 | 3 | 0.066 |
| Dissolved oxygen × Sampling time | 0.47 | 9 | 0.883 |
| Hsp70 mRNA | |||
| Dissolved oxygen | 46.64 | 3 | <0.001 |
| Sampling time | 7.70 | 3 | 0.001 |
| Dissolved oxygen × Sampling time | 7.13 | 9 | <0.001 |
| Crustin 4 mRNA | |||
| Dissolved oxygen | 130.75 | 3 | <0.001 |
| Sampling time | 8.33 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 8.17 | 9 | <0.001 |
| Variable of Treatment | F Value | df | p Value |
|---|---|---|---|
| SOD Mrna | |||
| Dissolved oxygen | 27.30 | 3 | <0.001 |
| Sampling time | 21.46 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 6.70 | 9 | <0.001 |
| CAT Mrna | |||
| Dissolved oxygen | 33.05 | 3 | <0.001 |
| Sampling time | 19.24 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 5.20 | 9 | <0.001 |
| Hsp70 Mrna | |||
| Dissolved oxygen | 27.74 | 3 | <0.001 |
| Sampling time | 8.73 | 3 | <0.001 |
| Dissolved oxygen × Sampling time | 1.61 | 9 | 0.154 |
| Crustin 4 Mrna | |||
| Dissolved oxygen | 78.54 | 3 | <0.001 |
| Sampling time | 5.32 | 3 | 0.004 |
| Dissolved oxygen × Sampling time | 1.53 | 9 | 0.178 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Q.; Luo, M.; Qin, L.; Guo, C.; Liu, J.; Zhang, T.; Feng, G.; Li, W. Effects of Hypoxia Stress on Survival, Antioxidant and Anaerobic Metabolic Enzymes, and Related Gene Expression of Red Swamp Crayfish Procambarus clarkii. Biology 2024, 13, 33. https://doi.org/10.3390/biology13010033
Zeng Q, Luo M, Qin L, Guo C, Liu J, Zhang T, Feng G, Li W. Effects of Hypoxia Stress on Survival, Antioxidant and Anaerobic Metabolic Enzymes, and Related Gene Expression of Red Swamp Crayfish Procambarus clarkii. Biology. 2024; 13(1):33. https://doi.org/10.3390/biology13010033
Chicago/Turabian StyleZeng, Qinghui, Mingzhong Luo, Lirong Qin, Chao Guo, Jiashou Liu, Tanglin Zhang, Guangpeng Feng, and Wei Li. 2024. "Effects of Hypoxia Stress on Survival, Antioxidant and Anaerobic Metabolic Enzymes, and Related Gene Expression of Red Swamp Crayfish Procambarus clarkii" Biology 13, no. 1: 33. https://doi.org/10.3390/biology13010033
APA StyleZeng, Q., Luo, M., Qin, L., Guo, C., Liu, J., Zhang, T., Feng, G., & Li, W. (2024). Effects of Hypoxia Stress on Survival, Antioxidant and Anaerobic Metabolic Enzymes, and Related Gene Expression of Red Swamp Crayfish Procambarus clarkii. Biology, 13(1), 33. https://doi.org/10.3390/biology13010033