Contributions of Chondroitin Sulfate, Keratan Sulfate and N-linked Oligosaccharides to Inhibition of Neurite Outgrowth by Aggrecan


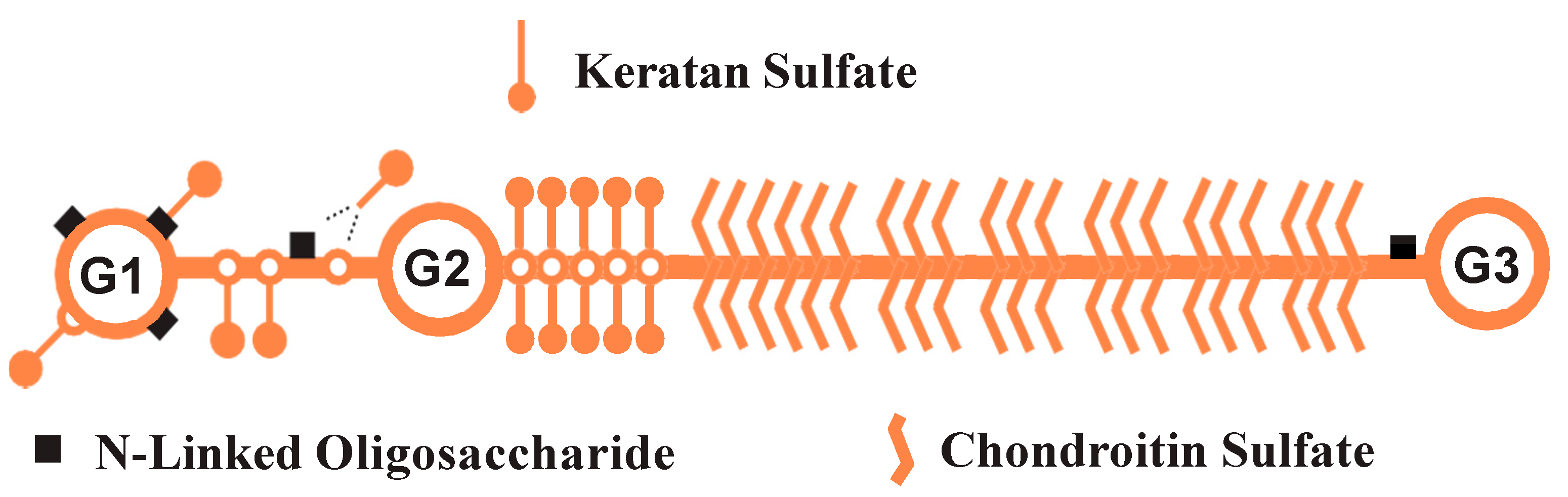{kind=link}
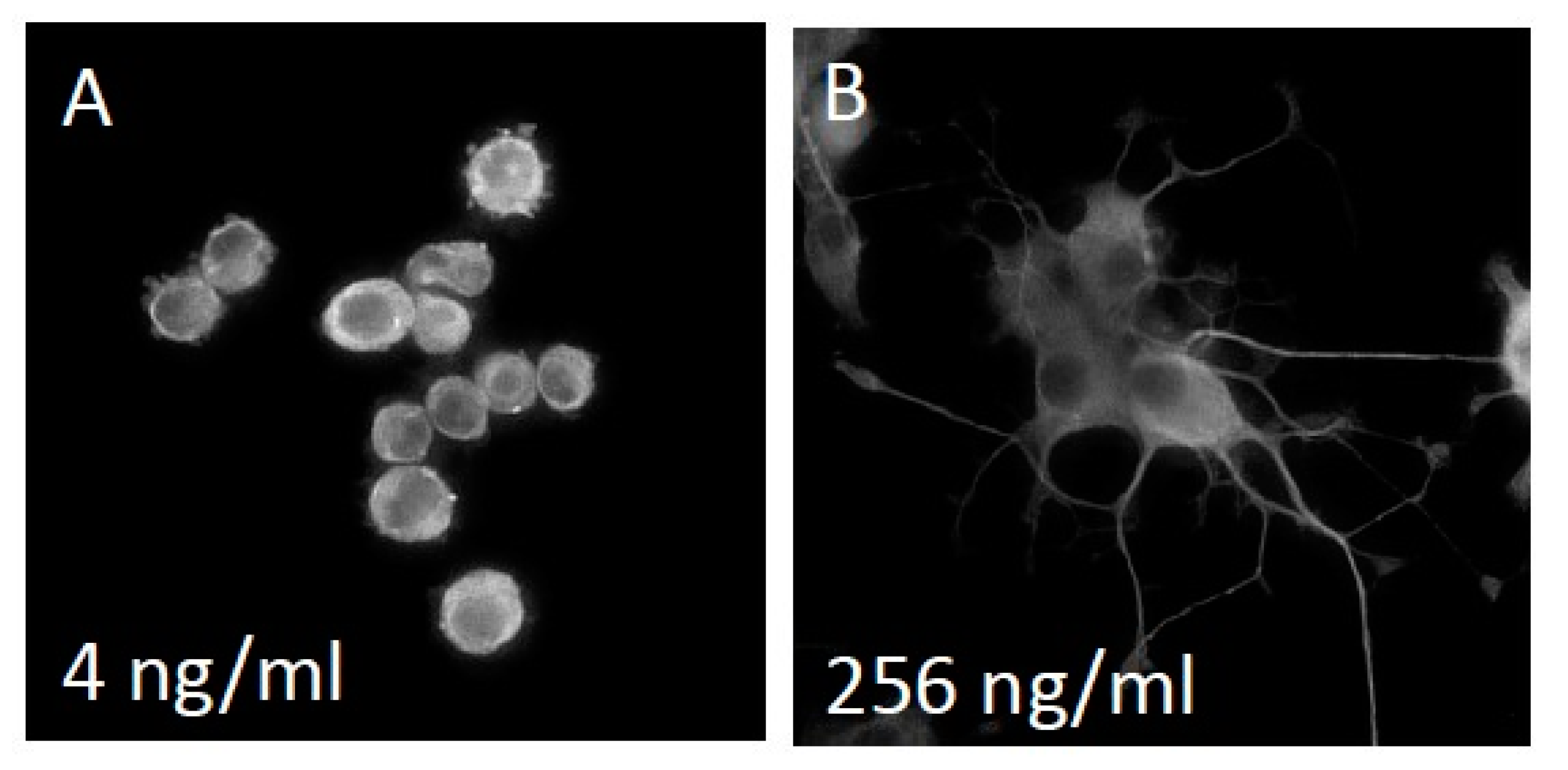{kind=link}
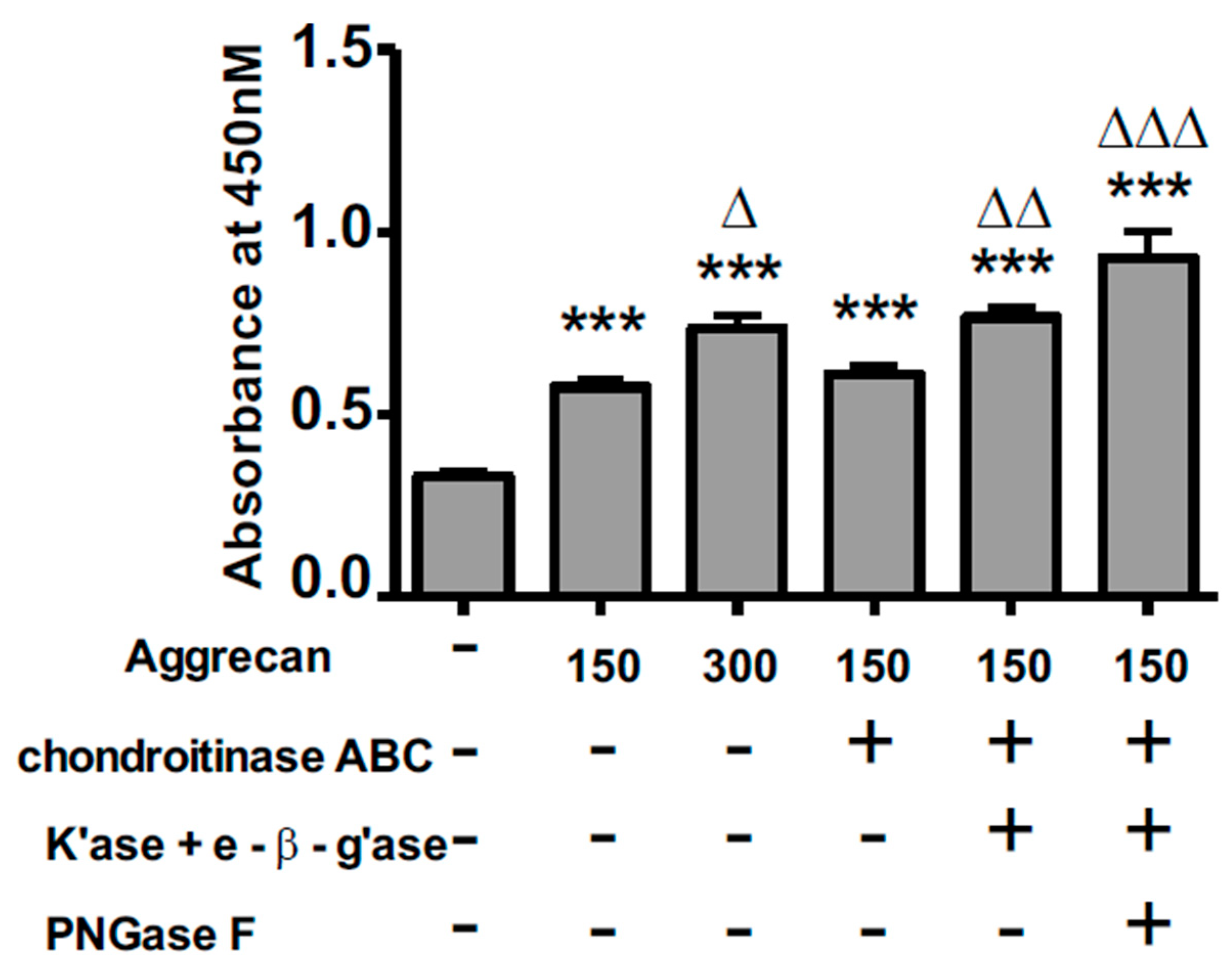{kind=link}
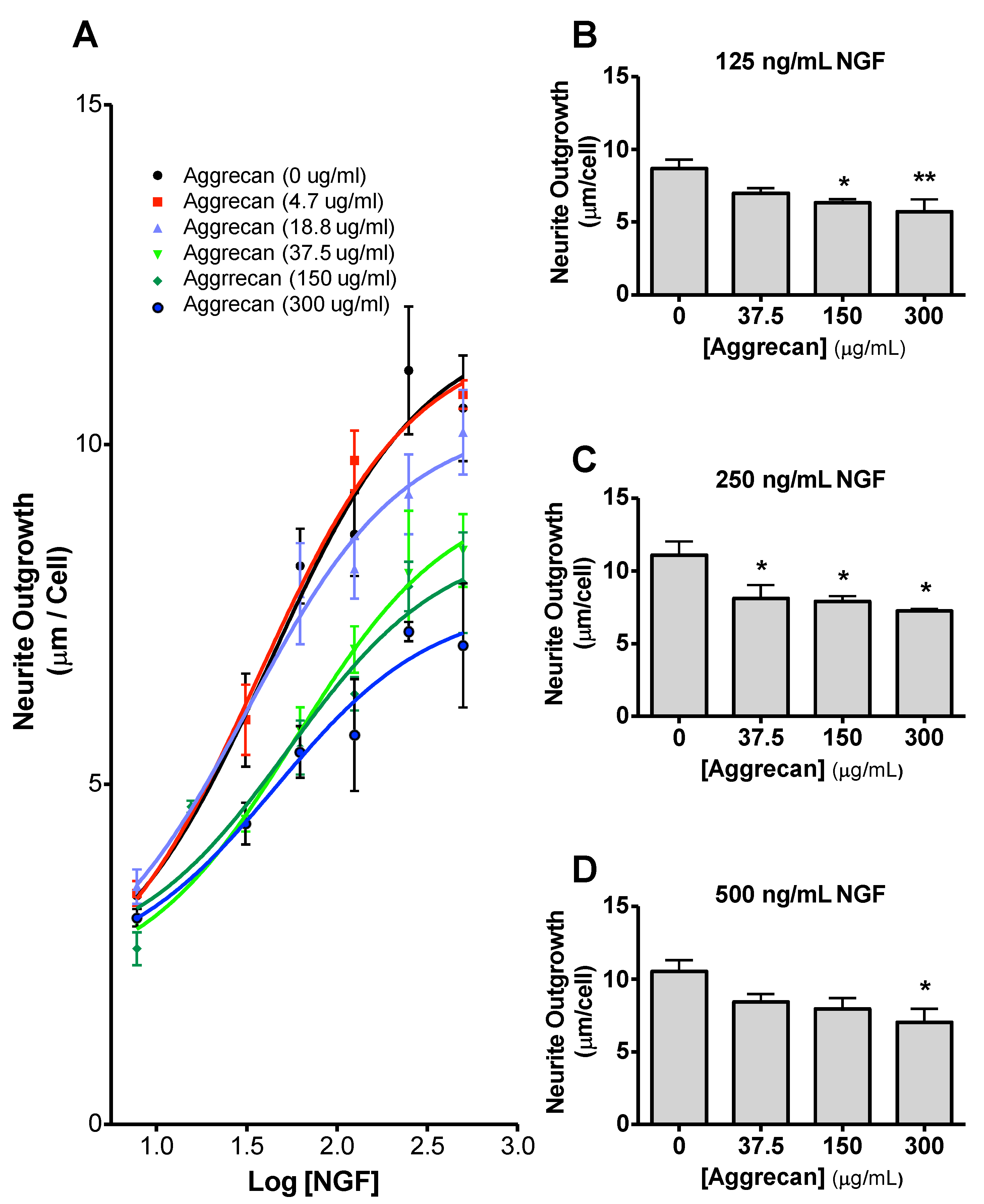{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Aggrecan
2.2. Enzyme Digestions
2.3. ELISA Assay
2.4. Neurite Outgrowth Assay
2.5. Image Acquisition
2.6. Data Analysis
3. Results
3.1. NS-1 Cells Extend Neurites in a Dose-Dependent Fashion When Exposed to Varying Amounts of NGF
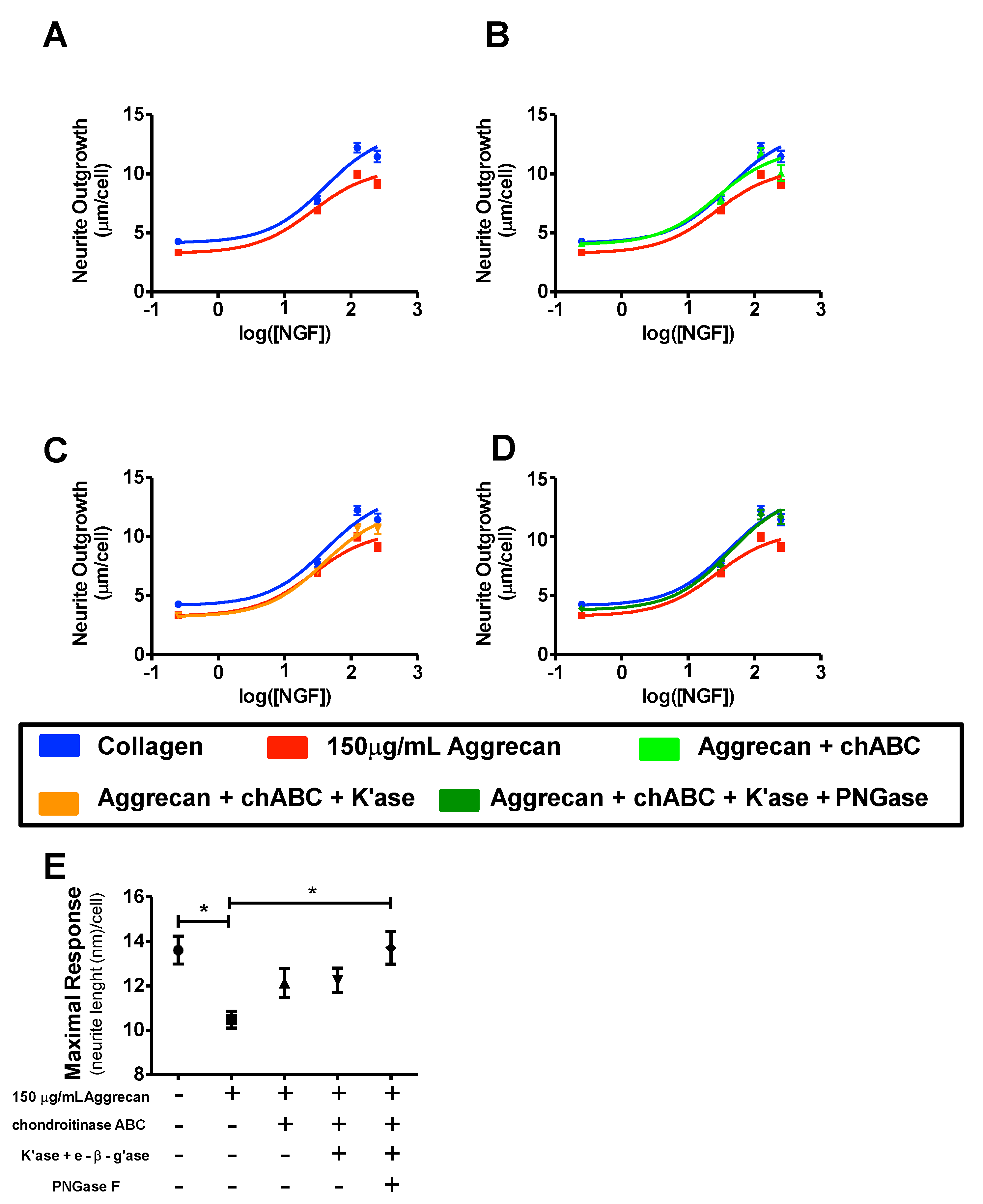
3.2. ELISA Assay Confirms Adsorption to Wells and Reveals Greater Access to G1-Domain Following Enzymatic Digestion
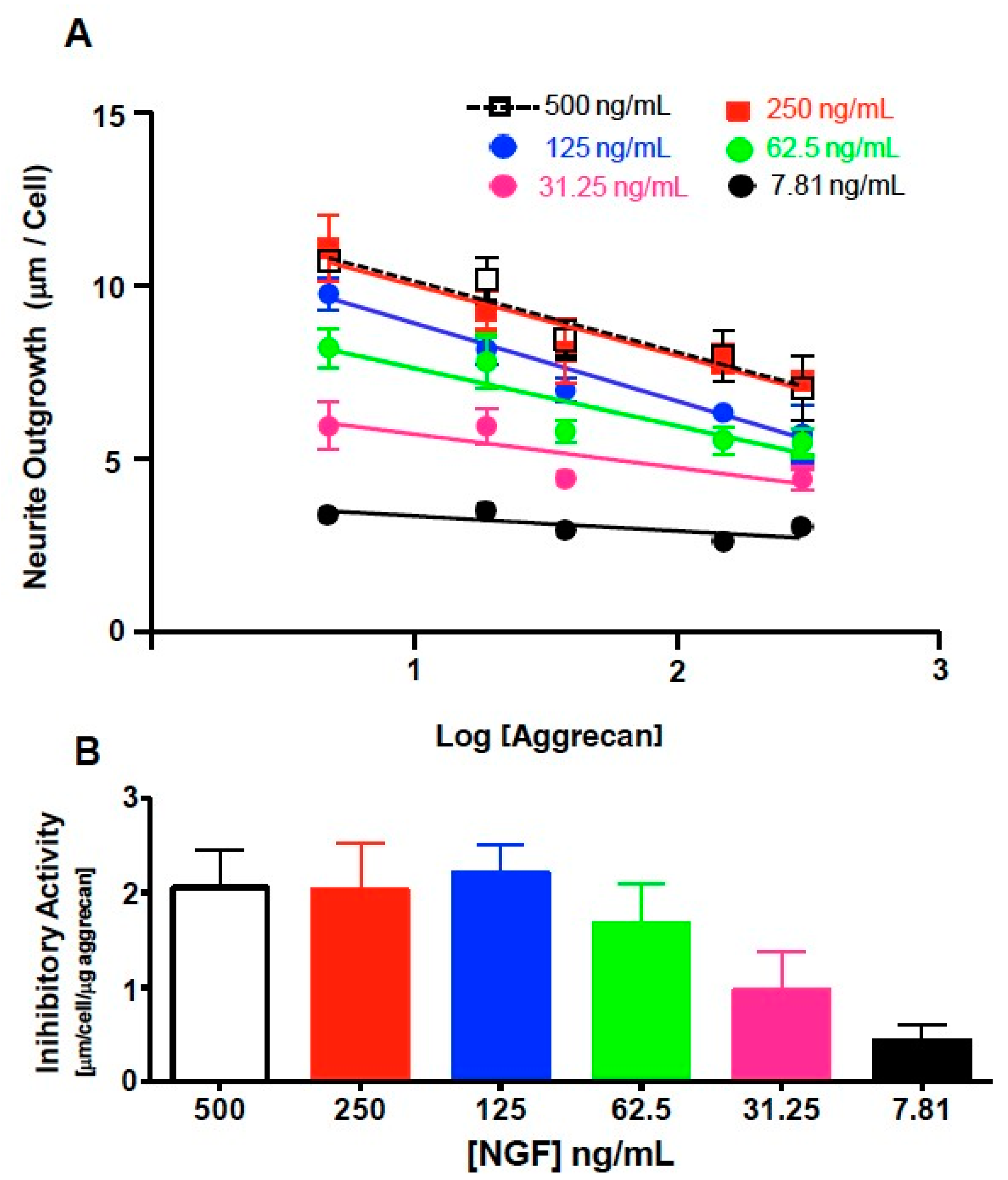
3.3. NS-1 Cell Neurite Outgrowth is Inhibited in a Concentration-Dependent Manner by Bound Aggrecan
3.4. Quantification of Inhibition of NGF-Stimulated Outgrowth Due to Aggrecan
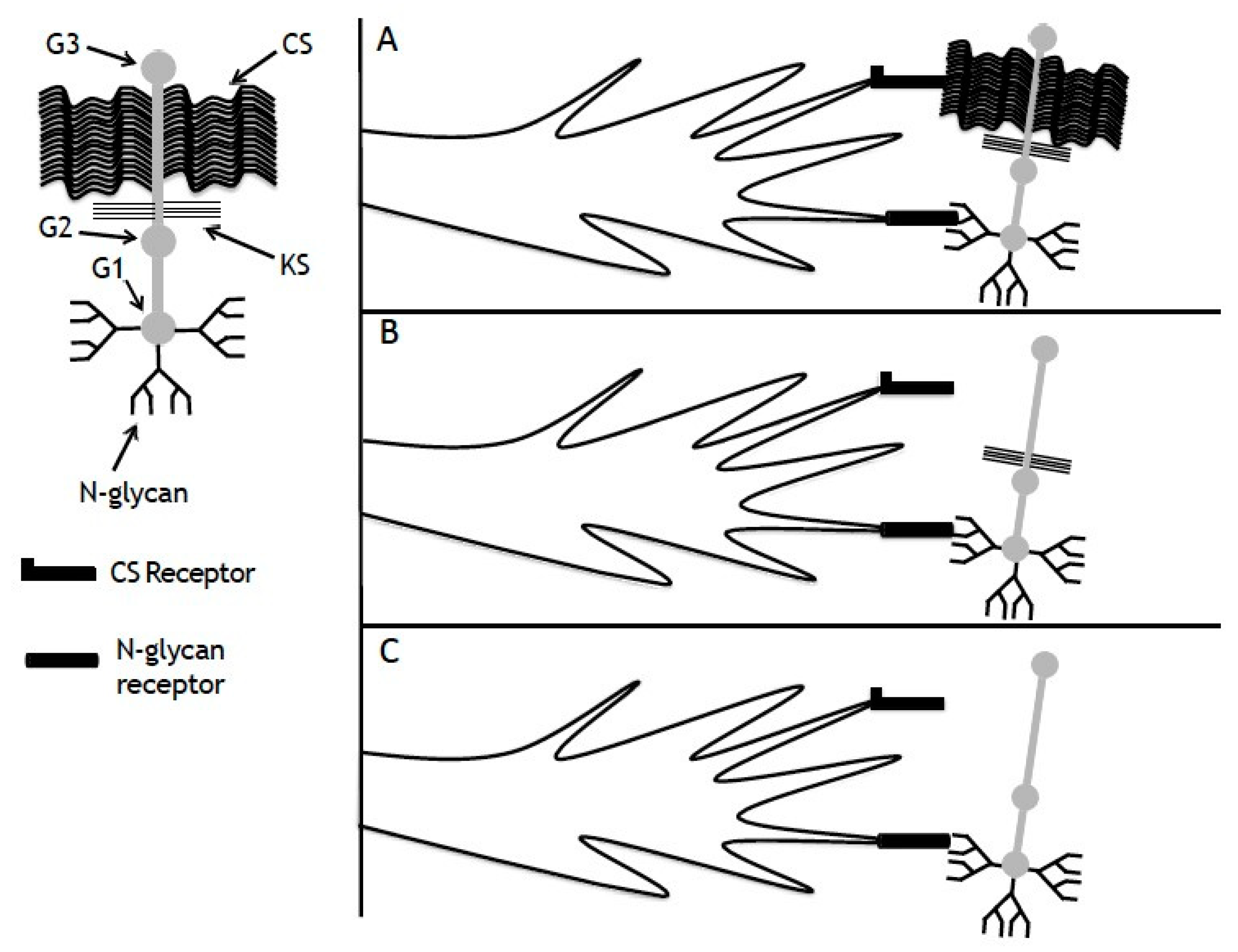
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kresse, H.; Schonherr, E. Proteoglycans of the extracellular matrix and growth control. J. Cell. Physiol. 2001, 189, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Beller, J.A.; Kulengowski, B.; Kobraei, E.M.; Curinga, G.; Calulot, C.M.; Bahrami, A.; Hering, T.M.; Snow, D.M. Comparison of sensory neuron growth cone and filopodial responses to structurally diverse aggrecan variants, In Vitro. Exp. Neurol. 2013, 247, 143–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, D.M.; Lemmon, V.; Carrino, D.A.; Caplan, A.I.; Silver, J. Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth In Vitro. Exp. Neurol. 1990, 109, 111–130. [Google Scholar] [CrossRef]
- Snow, D.M.; Watanabe, M.; Letourneau, P.C.; Silver, J. A chondroitin sulfate proteoglycan may influence the direction of retinal ganglion cell outgrowth. Development 1991, 113, 1473–1485. [Google Scholar]
- Zuo, J.; Neubauer, D.; Dyess, K.; Ferguson, T.A.; Muir, D. Degradation of chondroitin sulfate proteoglycan enhances the neurite-promoting potential of spinal cord tissue. Exp. Neurol. 1998, 154, 654–662. [Google Scholar] [CrossRef]
- Massey, J.M.; Amps, J.; Viapiano, M.S.; Matthews, R.T.; Wagoner, M.R.; Whitaker, C.M.; Alilain, W.; Yonkof, A.L.; Khalyfa, A.; Cooper, N.G.; et al. Increased chondroitin sulfate proteoglycan expression in denervated brainstem targets following spinal cord injury creates a barrier to axonal regeneration overcome by chondroitinase ABC and neurotrophin-3. Exp. Neurol. 2008, 209, 426–445. [Google Scholar] [CrossRef] [Green Version]
- Emerling, D.E.; Lander, A.D. Inhibitors and promoters of thalamic neuron adhesion and outgrowth in embryonic neocortex: Functional association with chondroitin sulfate. Neuron 1996, 17, 1089–1100. [Google Scholar] [CrossRef] [Green Version]
- Faissner, A.; Clement, A.; Lochter, A.; Streit, A.; Mandl, C.; Schachner, M. Isolation of a neural chondroitin sulfate proteoglycan with neurite outgrowth promoting properties. J. Cell. Biol. 1994, 126, 783–799. [Google Scholar] [CrossRef]
- Johnson, W.E.; Caterson, B.; Eisenstein, S.M.; Hynds, D.L.; Snow, D.M.; Roberts, S. Human intervertebral disc aggrecan inhibits nerve growth In Vitro. Arthritis Rheum. 2002, 46, 2658–2664. [Google Scholar] [CrossRef]
- Condic, M.L.; Snow, D.M.; Letourneau, P.C. Embryonic neurons adapt to the inhibitory proteoglycan aggrecan by increasing integrin expression. J. Neurosci. 1999, 19, 10036–10043. [Google Scholar] [CrossRef]
- Kwok, J.C.; Carulli, D.; Fawcett, J.W. In Vitro modeling of perineuronal nets: Hyaluronan synthase and link protein are necessary for their formation and integrity. J. Neurochem. 2010, 114, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Kwok, J.C.; Dick, G.; Wang, D.; Fawcett, J.W. Extracellular matrix and perineuronal nets in CNS repair. Dev. Neurobiol. 2011, 71, 1073–1089. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Giger, R.J. A new role for RPTPsigma in spinal cord injury: Signaling chondroitin sulfate proteoglycan inhibition. Sci. Signal. 2010, 3, pe6. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Tenney, A.P.; Busch, S.A.; Horn, K.P.; Cuascut, F.X.; Liu, K.; He, Z.; Silver, J.; Flanagan, J.G. PTPsigma is a receptor for chondroitin sulfate proteoglycan, an inhibitor of neural regeneration. Science 2009, 326, 592–596. [Google Scholar] [CrossRef] [Green Version]
- Dickendesher, T.L.; Baldwin, K.T.; Mironova, Y.A.; Koriyama, Y.; Raiker, S.J.; Askew, K.L.; Wood, A.; Geoffroy, C.G.; Zheng, B.; Liepmann, C.D.; et al. NgR1 and NgR3 are receptors for chondroitin sulfate proteoglycans. Nat. Neurosci. 2012, 15, 703–712. [Google Scholar] [CrossRef]
- Brown, J.M.; Xia, J.; Zhuang, B.; Cho, K.S.; Rogers, C.J.; Gama, C.I.; Rawat, M.; Tully, S.E.; Uetani, N.; Mason, D.E.; et al. A sulfated carbohydrate epitope inhibits axon regeneration after injury. Proc. Natl. Acad. Sci. USA 2012, 109, 4768–4773. [Google Scholar] [CrossRef] [Green Version]
- Hering, T.M.; Kollar, J.; Huynh, T.D. Complete coding sequence of bovine aggrecan: Comparative structural analysis. Arch. Biochem. Biophys. 1997, 345, 259–270. [Google Scholar] [CrossRef]
- Monnier, P.P.; Sierra, A.; Schwab, J.M.; Henke-Fahle, S.; Mueller, B.K. The Rho/ROCK pathway mediates neurite growth-inhibitory activity associated with the chondroitin sulfate proteoglycans of the CNS glial scar. Mol. Cell. Neurosci. 2003, 22, 319–330. [Google Scholar] [CrossRef]
- Snow, D.M.; Mullins, N.; Hynds, D.L. Nervous system-derived chondroitin sulfate proteoglycans regulate growth cone morphology and inhibit neurite outgrowth: A light, epifluorescence, and electron microscopy study. Microsc. Res. Tech. 2001, 54, 273–286. [Google Scholar] [CrossRef]
- Lingor, P.; Teusch, N.; Schwarz, K.; Mueller, R.; Mack, H.; Bahr, M.; Mueller, B.K. Inhibition of Rho kinase (ROCK) increases neurite outgrowth on chondroitin sulphate proteoglycan In Vitro and axonal regeneration in the adult optic nerve in vivo. J. Neurochem. 2007, 103, 181–189. [Google Scholar] [CrossRef]
- Oohira, A.; Matsui, F.; Katoh-Semba, R. Inhibitory effects of brain chondroitin sulfate proteoglycans on neurite outgrowth from PC12D cells. J. Neurosci. 1991, 11, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, D.M.; Brown, E.M.; Letourneau, P.C. Growth cone behavior in the presence of soluble chondroitin sulfate proteoglycan (CSPG), compared to behavior on CSPG bound to laminin or fibronectin. Int. J. Dev. Neurosci. 1996, 14, 331–349. [Google Scholar] [CrossRef]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yick, L.W.; Cheung, P.T.; So, K.F.; Wu, W. Axonal regeneration of Clarke’s neurons beyond the spinal cord injury scar after treatment with chondroitinase ABC. Exp. Neurol. 2003, 182, 160–168. [Google Scholar] [CrossRef]
- Barritt, A.W.; Davies, M.; Marchand, F.; Hartley, R.; Grist, J.; Yip, P.; McMahon, S.B.; Bradbury, E.J. Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury. J. Neurosci. 2006, 26, 10856–10867. [Google Scholar] [CrossRef] [PubMed]
- Akyuz, N.; Rost, S.; Mehanna, A.; Bian, S.; Loers, G.; Oezen, I.; Mishra, B.; Hoffmann, K.; Guseva, D.; Laczynska, E.; et al. Dermatan 4-O-sulfotransferase1 ablation accelerates peripheral nerve regeneration. Exp. Neurol. 2013, 247, 517–530. [Google Scholar] [CrossRef]
- Imagama, S.; Sakamoto, K.; Tauchi, R.; Shinjo, R.; Ohgomori, T.; Ito, Z.; Zhang, H.; Nishida, Y.; Asami, N.; Takeshita, S.; et al. Keratan sulfate restricts neural plasticity after spinal cord injury. J. Neurosci. 2011, 31, 17091–17102. [Google Scholar] [CrossRef] [Green Version]
- Ito, Z.; Sakamoto, K.; Imagama, S.; Matsuyama, Y.; Zhang, H.; Hirano, K.; Ando, K.; Yamashita, T.; Ishiguro, N.; Kadomatsu, K. N-acetylglucosamine 6-O-sulfotransferase-1-deficient mice show better functional recovery after spinal cord injury. J. Neurosci. 2010, 30, 5937–5947. [Google Scholar] [CrossRef]
- Beller, J.A.; Hering, T.M.; Snow, D.M. High-throughput quantitative assay for analyzing neurite outgrowth on a uniform substratum: The cell substratum assay. In Neuromethods: Extracellular Matrix; Leach, J.B., Powell, E.M., Eds.; Humana Press: New York, NY, USA, 2014. [Google Scholar]
- Gopalakrishnan, S.M.; Teusch, N.; Imhof, C.; Bakker, M.H.; Schurdak, M.; Burns, D.J.; Warrior, U. Role of Rho kinase pathway in chondroitin sulfate proteoglycan-mediated inhibition of neurite outgrowth in PC12 cells. J. Neurosci. Res. 2008, 86, 2214–2226. [Google Scholar] [CrossRef]
- Heinegard, D. Extraction, fractionation and characterization of proteoglycans from bovine tracheal cartilage. Biochim. Biophys. Acta 1972, 285, 181–192. [Google Scholar] [CrossRef]
- Rosenberg, L.; Choi, H.U.; Tang, L.H.; Pal, S.; Johnson, T.; Lyons, D.A.; Laue, T.M. Proteoglycans of bovine articular cartilage. The effects of divalent cations on the biochemical properties of link protein. J. Biol. Chem. 1991, 266, 7016–7024. [Google Scholar] [PubMed]
- Hering, T.M.; Beller, J.A.; Calulot, C.M.; Centers, A.; Snow, D.M. Proteoglycans of reactive rat cortical astrocyte cultures: Abundance of N-unsubstituted glucosamine-enriched heparan sulfate. Matrix Biol. 2015, 41, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Pool, M.; Thiemann, J.; Bar-Or, A.; Fournier, A.E. NeuriteTracer: A novel ImageJ plugin for automated quantification of neurite outgrowth. J. Neurosci. Methods 2008, 168, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.M.; Letourneau, P.C. Neurite outgrowth on a step gradient of chondroitin sulfate proteoglycan (CS-PG). J. Neurobiol. 1992, 23, 322–336. [Google Scholar] [CrossRef]
- Greene, L.A. Nerve growth factor prevents the death and stimulates the neuronal differentiation of clonal PC12 pheochromocytoma cells in serum-free medium. J. Cell. Biol. 1978, 78, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, E.J.; Moon, L.D.; Popat, R.J.; King, V.R.; Bennett, G.S.; Patel, P.N.; Fawcett, J.W.; McMahon, S.B. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 2002, 416, 636–640. [Google Scholar] [CrossRef]
- Lee, H.; McKeon, R.J.; Bellamkonda, R.V. Sustained delivery of thermostabilized chABC enhances axonal sprouting and functional recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2010, 107, 3340–3345. [Google Scholar] [CrossRef] [Green Version]
- Massey, J.M.; Hubscher, C.H.; Wagoner, M.R.; Decker, J.A.; Amps, J.; Silver, J.; Onifer, S.M. Chondroitinase ABC digestion of the perineuronal net promotes functional collateral sprouting in the cuneate nucleus after cervical spinal cord injury. J. Neurosci. 2006, 26, 4406–4414. [Google Scholar] [CrossRef]
- Huang, W.C.; Kuo, W.C.; Cherng, J.H.; Hsu, S.H.; Chen, P.R.; Huang, S.H.; Huang, M.C.; Liu, J.C.; Cheng, H. Chondroitinase ABC promotes axonal re-growth and behavior recovery in spinal cord injury. Biochem. Biophys. Res. Commun. 2006, 349, 963–968. [Google Scholar] [CrossRef]
- Nakamae, T.; Tanaka, N.; Nakanishi, K.; Kamei, N.; Sasaki, H.; Hamasaki, T.; Yamada, K.; Yamamoto, R.; Mochizuki, Y.; Ochi, M. Chondroitinase ABC promotes corticospinal axon growth in organotypic cocultures. Spinal Cord 2009, 47, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Garwood, J.; Schnadelbach, O.; Clement, A.; Schutte, K.; Bach, A.; Faissner, A. DSD-1-proteoglycan is the mouse homolog of phosphacan and displays opposing effects on neurite outgrowth dependent on neuronal lineage. J. Neurosci. 1999, 19, 3888–3899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidler, P.S.; Schuette, K.; Asher, R.A.; Dobbertin, A.; Thornton, S.R.; Calle-Patino, Y.; Muir, E.; Levine, J.M.; Geller, H.M.; Rogers, J.H.; et al. Comparing astrocytic cell lines that are inhibitory or permissive for axon growth: The major axon-inhibitory proteoglycan is NG2. J. Neurosci. 1999, 19, 8778–8788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, G.J.; McCabe, C.F. Identification of a developmentally regulated keratan sulfate proteoglycan that inhibits cell adhesion and neurite outgrowth. Neuron 1991, 7, 1007–1018. [Google Scholar] [CrossRef]
- Mace, K.; Saxod, R.; Feuerstein, C.; Sadoul, R.; Hemming, F.J. Chondroitin and keratan sulfates have opposing effects on attachment and outgrowth of ventral mesencephalic explants in culture. J. Neurosci. Res 2002, 70, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Charvet, I.; Hemming, F.J.; Feuerstein, C.; Saxod, R. Mosaic distribution of chondroitin and keratan sulphate in the developing rat striatum: Possible involvement of proteoglycans in the organization of the nigrostriatal system. Brain Res. Dev. Brain. Res. 1998, 109, 229–244. [Google Scholar] [CrossRef]
- Helenius, A.; Aebi, M. Intracellular functions of N-linked glycans. Science 2001, 291, 2364–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, J.W. N-linked oligosaccharide processing and tumor cell biology. Semin. Cancer Biol. 1991, 2, 411–420. [Google Scholar]
- Varki, A. Biological roles of oligosaccharides: All of the theories are correct. Glycobiology 1993, 3, 97–130. [Google Scholar] [CrossRef]
- Takahashi, M.; Tsuda, T.; Ikeda, Y.; Honke, K.; Taniguchi, N. Role of N-glycans in growth factor signaling. Glycoconj. J. 2004, 20, 207–212. [Google Scholar] [CrossRef]
- Parkkinen, J.; Raulo, E.; Merenmies, J.; Nolo, R.; Kajander, E.O.; Baumann, M.; Rauvala, H. Amphoterin, the 30-kDa protein in a family of HMG1-type polypeptides. Enhanced expression in transformed cells, leading edge localization, and interactions with plasminogen activation. J. Biol. Chem. 1993, 268, 19726–19738. [Google Scholar]
- Srikrishna, G.; Huttunen, H.J.; Johansson, L.; Weigle, B.; Yamaguchi, Y.; Rauvala, H.; Freeze, H.H. N -Glycans on the receptor for advanced glycation end products influence amphoterin binding and neurite outgrowth. J. Neurochem. 2002, 80, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Guo, H.B.; Kamar, M.; Abbott, K.; Troupe, K.; Lee, J.K.; Alvarez-Manilla, G.; Pierce, M. N-acetylglucosaminyltranferase VB expression enhances beta1 integrin- dependent PC12 neurite outgrowth on laminin and collagen. J. Neurochem. 2006, 97, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Horstkorte, R.; Schachner, M.; Magyar, J.P.; Vorherr, T.; Schmitz, B. The fourth immunoglobulin-like domain of NCAM contains a carbohydrate recognition domain for oligomannosidic glycans implicated in association with L1 and neurite outgrowth. J. Cell. Biol. 1993, 121, 1409–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.R.; Fawcett, J.W. Combination treatment with chondroitinase ABC in spinal cord injury--breaking the barrier. Neurosci. Bull. 2013, 29, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Hilton, B.J.; Lang, B.T.; Cregg, J.M. Keratan sulfate proteoglycans in plasticity and recovery after spinal cord injury. J. Neurosci. 2012, 32, 4331–4333. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.L.; Tuszynski, M.H. Spinal cord injury elicits expression of keratan sulfate proteoglycans by macrophages, reactive microglia, and oligodendrocyte progenitors. J. Neurosci. 2002, 22, 4611–4624. [Google Scholar] [CrossRef] [Green Version]
- McAdams, B.D.; McLoon, S.C. Expression of chondroitin sulfate and keratan sulfate proteoglycans in the path of growing retinal axons in the developing chick. J. Comp. Neurol 1995, 352, 594–606. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hering, T.M.; Beller, J.A.; Calulot, C.M.; Snow, D.M. Contributions of Chondroitin Sulfate, Keratan Sulfate and N-linked Oligosaccharides to Inhibition of Neurite Outgrowth by Aggrecan. Biology 2020, 9, 29. https://doi.org/10.3390/biology9020029
Hering TM, Beller JA, Calulot CM, Snow DM. Contributions of Chondroitin Sulfate, Keratan Sulfate and N-linked Oligosaccharides to Inhibition of Neurite Outgrowth by Aggrecan. Biology. 2020; 9(2):29. https://doi.org/10.3390/biology9020029
Chicago/Turabian StyleHering, Thomas M., Justin A. Beller, Christopher M. Calulot, and Diane M. Snow. 2020. "Contributions of Chondroitin Sulfate, Keratan Sulfate and N-linked Oligosaccharides to Inhibition of Neurite Outgrowth by Aggrecan" Biology 9, no. 2: 29. https://doi.org/10.3390/biology9020029
APA StyleHering, T. M., Beller, J. A., Calulot, C. M., & Snow, D. M. (2020). Contributions of Chondroitin Sulfate, Keratan Sulfate and N-linked Oligosaccharides to Inhibition of Neurite Outgrowth by Aggrecan. Biology, 9(2), 29. https://doi.org/10.3390/biology9020029