
In Vitro Efficacy of Hungarian Propolis against Bacteria, Yeast, and Trichomonas gallinae Isolated from Pigeons—A Possible Antibiotic Alternative?



Abstract
:1. Introduction
2. Materials and Methods
2.1. The Origin and Extraction of Propolis
2.2. Antibacterial and Antifungal Studies
2.3. Antiprotozoal Studies
2.4. Statistical Analysis
3. Results
3.1. Efficacy of Propolis against Bacteria and Fungi
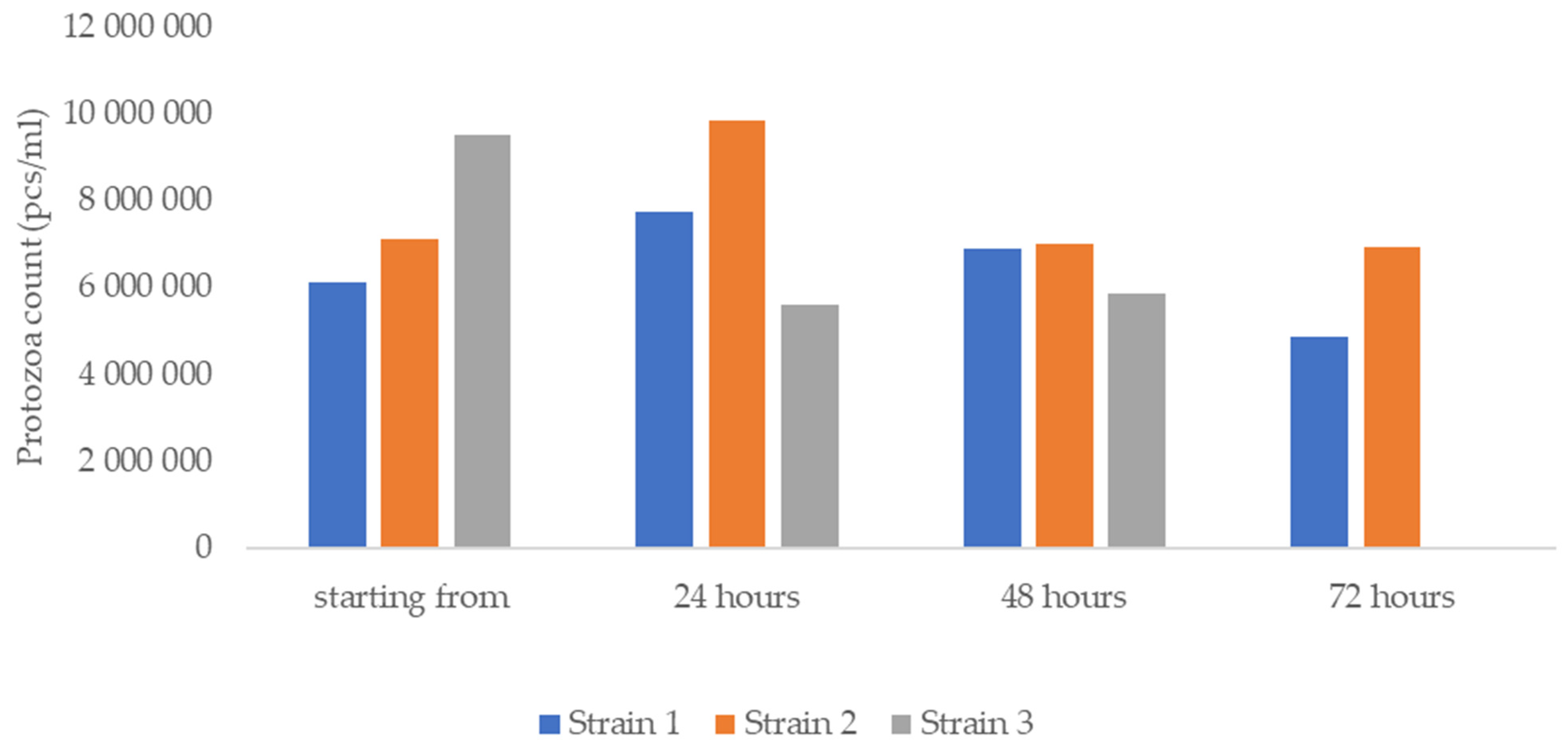
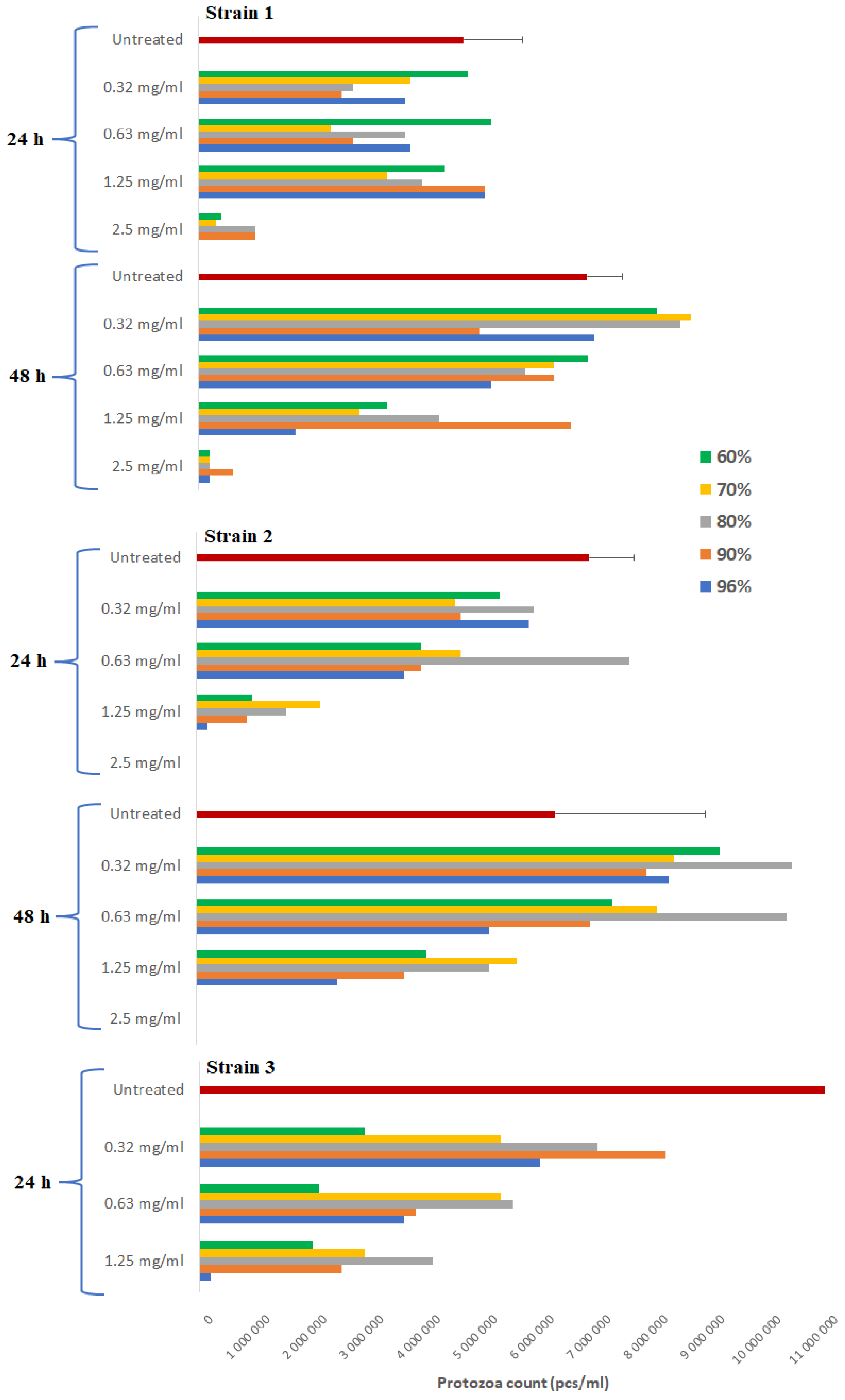
3.2. Viability of Protozoa and Activity of Propolis against T. gallinae
3.3. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, L.M.; Fonseca, M.S.; Sokolonski, A.R.; Deegan, K.R.; Araújo, R.P.; Umsza-Guez, M.A.; Barbosa, J.D.; Portela, R.D.; Machado, B.A. Propolis: Types, composition, biological activities, and veterinary product patent prospecting. J. Sci. Food Agric. 2020, 100, 1369–1382. [Google Scholar] [CrossRef]
- Daneshmand, A.; Sadeghi, G.H.; Karimi, A.; Vaziry, A.; Ibrahim, S.A. Evaluating complementary effects of ethanol extract of propolis with the probiotic on growth performance, immune response and serum metabolites in male broiler chickens. Livest. Sci. 2015, 178, 195–201. [Google Scholar] [CrossRef]
- Aygun, A.; Sert, D. Effects of prestorage application of propolis and storage time on eggshell microbial activity, hatchability, and chick performance in Japanese quail (Coturnix coturnix japonica) eggs. Poult. Sci. 2013, 92, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Matsumiya, T.; Hayakari, R.; Shiba, Y.; Kawaguchi, S.; Seya, K.; Ueno, K.; Imaizumi, T. Daily Brazilian green propolis intake elevates blood artepillin C levels in humans. J. Sci. Food Agric. 2021, 101, 4855–4861. [Google Scholar] [CrossRef]
- Almuhayawi, M.S. Propolis as a novel antibacterial agent. Saudi J. Biol. Sci. 2020, 27, 3079–3086. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed]
- Kowacz, M.; Pollack, G.H. Propolis-induced exclusion of colloids: Possible new mechanism of biological action. Colloid Interface Sci. Commun. 2020, 38, 100307. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Yañez, N.; Rivera-Yañez, C.R.; Pozo-Molina, G.; Méndez-Catalá, C.F.; Reyes-Reali, J.; Mendoza-Ramos, M.I.; Méndez-Cruz, A.R.; Nieto-Yañez, O. Effects of Propolis on Infectious Diseases of Medical Relevance. Biology 2021, 10, 428. [Google Scholar] [CrossRef]
- Silva-Carvalho, R.; Baltazar, F.; Almeida-Aguiar, C. Propolis: A Complex Natural Product with a Plethora of Biological Activities That Can Be Explored for Drug Development. Evid.-Based Complement. Altern. Med. 2015, 2015, 206439. [Google Scholar] [CrossRef]
- Regueira-Neto, M.d.S.; Tintino, S.R.; Rolón, M.; Coronal, C.; Vega, M.C.; de Queiroz Balbino, V.; de Melo Coutinho, H.D. Antitrypanosomal, antileishmanial and cytotoxic activities of Brazilian red propolis and plant resin of Dalbergia ecastaphyllum (L) Taub. Food Chem. Toxicol. 2018, 119, 215–221. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Gómez-Romero, M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Advances in the analysis of phenolic compounds in products derived from bees. J. Pharm. Biomed. Anal. 2006, 41, 1220–1234. [Google Scholar] [CrossRef]
- Farnesi, A.P.; Aquino-Ferreira, R.; De Jong, D.; Bastos, J.K.; Soares, A.E.E. Effects of stingless bee and honey bee propolis on four species of bacteria. Genet. Mol. Res. 2009, 8, 635–640. [Google Scholar] [CrossRef]
- Liberio, S.A.; Pereira, A.L.A.; Dutra, R.P.; Reis, A.S.; Araújo, M.J.A.M.; Mattar, N.S.; Silva, L.A.; Ribeiro, M.N.S.; Nascimento, F.R.F.; Guerra, R.N.M.; et al. Antimicrobial activity against oral pathogens and immunomodulatory effects and toxicity of geopropolis produced by the stingless bee Melipona fasciculata Smith. BMC Complement. Altern. Med. 2011, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Al-Khalaifah, H.; Ibrahim, M.S.; Al-Hamid, A.E.A.; Al-Harthi, M.A.; El-Naggar, A. Blood Hematological and Biochemical Constituents, Antioxidant Enzymes, Immunity and Lymphoid Organs of Broiler Chicks Supplemented with Propolis, Bee Pollen and Mannan Oligosaccharides Continuously or Intermittently. Poult. Sci. 2017, 96, 4182–4192. [Google Scholar] [CrossRef] [PubMed]
- Kasote, D.; Bankova, V.; Viljoen, A.M. Propolis: Chemical diversity and challenges in quality control. Phytochem. Rev. 2022, 21, 1887–1911. [Google Scholar] [CrossRef]
- Állatállomány, 1 December 2019. Available online: https://www.ksh.hu/docs/hun/xftp/stattukor/allat/2019/index.html (accessed on 8 October 2021).
- Ghanbarpour, R.; Aflatoonian, M.R.; Askari, A.; Abiri, Z.; Naderi, Z.; Bagheri, M.; Jajarmi, M.; Shobeiri, S.; Molaei, R.; Askari, N. Domestic and game pigeons as reservoirs for Escherichia coli harbouring antimicrobial resistance genes. J. Glob. Antimicrob. Resist. 2020, 22, 571–577. [Google Scholar] [CrossRef]
- Alhababi, D.A.; Eltai, N.O.; Nasrallah, G.K.; Farg, E.A.; Al Thani, A.A.; Yassine, H.M. Antimicrobial Resistance of Commensal Escherichia coli Isolated from Food Animals in Qatar. Microb. Drug Resist. 2020, 26, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Antić, N.; Dakić, I.; Svabić-Vlahović, M. In vitro antimicrobial activity of propolis and synergism between propolis and antimicrobial drugs. Microbiol. Res. 2003, 158, 353–357. [Google Scholar] [CrossRef]
- Zulhendri, F.; Chandrasekaran, K.; Kowacz, M.; Ravalia, M.; Kripal, K.; Fearnley, J.; Perera, C.O. Antiviral, Antibacterial, Antifungal, and Antiparasitic Properties of Propolis: A Review. Foods 2021, 10, 1360. [Google Scholar] [CrossRef]
- Mirzoeva, O.K.; Grishanin, R.N.; Calder, P.C. Antimicrobial action of propolis and some of its components: The effects on growth, membrane potential and motility of bacteria. Microbiol. Res. 1997, 152, 239–246. [Google Scholar] [CrossRef]
- Kerek, Á.; Csanády, P.; Jerzsele, Á. Antibacterial efficiency of propolis—Part 1. Magy. Állatorvosok Lapja 2022, 144, 285–298. [Google Scholar]
- Gucwa, K.; Kusznierewicz, B.; Milewski, S.; Van Dijck, P.; Szweda, P. Antifungal Activity and Synergism with Azoles of Polish Propolis. Pathogens 2018, 7, 56. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Górecki, M.; Rzepecka-Stojko, A.; Balwierz, R.; Stojko, J. Bee Products in Dermatology and Skin Care. Molecules 2020, 25, 556. [Google Scholar] [CrossRef]
- Anjum, S.I.; Ullah, A.; Khan, K.A.; Attaullah, M.; Khan, H.; Ali, H.; Bashir, M.A.; Tahir, M.; Ansari, M.J.; Ghramh, H.A.; et al. Composition and functional properties of propolis (bee glue): A review. Saudi J. Biol. Sci. 2019, 26, 1695–1703. [Google Scholar] [CrossRef]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Ádám, K.; Péter, C.; Ákos, J. Antiprotozoal and antifungal efficiency of propolis—Part 2. Magy. Állatorvosok Lapja 2022, 144, 691–704. [Google Scholar]
- Siheri, W.; Ebiloma, G.U.; Igoli, J.O.; Gray, A.I.; Biddau, M.; Akrachalanont, P.; Alenezi, S.; Alwashih, M.A.; Edrada-Ebel, R.; Muller, S.; et al. Isolation of a Novel Flavanonol and an Alkylresorcinol with Highly Potent Anti-Trypanosomal Activity from Libyan Propolis. Molecules 2019, 24, 1041. [Google Scholar] [CrossRef]
- Torres-Santos, E.C.; Sampaio-Santos, M.I.; Buckner, F.S.; Yokoyama, K.; Gelb, M.; Urbina, J.A.; Rossi-Bergmann, B. Altered sterol profile induced in Leishmania amazonensis by a natural dihydroxymethoxylated chalcone. J. Antimicrob. Chemother. 2009, 63, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Antwi, C.A.; Amisigo, C.M.; Adjimani, J.P.; Gwira, T.M. In vitro activity and mode of action of phenolic compounds on Leishmania donovani. PLoS Negl. Trop. Dis. 2019, 13, e0007206. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Silva, F.; Canto-Cavalheiro, M.M.; Menna-Barreto, R.F.S.; Almeida-Amaral, E.E. Effect of Apigenin on Leishmania amazonensis Is Associated with Reactive Oxygen Species Production Followed by Mitochondrial Dysfunction. J. Nat. Prod. 2015, 78, 880–884. [Google Scholar] [CrossRef]
- De Pablos, L.M.; González, G.; Rodrigues, R.; García Granados, A.; Parra, A.; Osuna, A. Action of a Pentacyclic Triterpenoid, Maslinic Acid, against Toxoplasma gondii. J. Nat. Prod. 2010, 73, 831–834. [Google Scholar] [CrossRef]
- Maróstica Junior, M.R.; Daugsch, A.; Moraes, C.S.; Queiroga, C.L.; Pastore, G.M.; Parki, Y.K. Comparison of volatile and polyphenolic compounds in Brazilian green propolis and its botanical origin Baccharis dracunculifolia. Food Sci. Technol. 2008, 28, 178–181. [Google Scholar] [CrossRef]
- Bolaños, V.; Díaz-Martínez, A.; Soto, J.; Marchat, L.A.; Sanchez-Monroy, V.; Ramírez-Moreno, E. Kaempferol inhibits Entamoeba histolytica growth by altering cytoskeletal functions. Mol. Biochem. Parasitol. 2015, 204, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Tully, T.N.; Lawton, M.P.C.; Dorrestein, G.M. (Eds.) Avian Medicine; Butterworth-Heinemann: Oxford, UK; Boston, MA, USA, 2000; ISBN 978-0-7506-3598-1. [Google Scholar]
- Arafa, M.I.; Hassan, H.H.; Mahmoed, W.G.M.; Abdel-Rahman, M.F. Study the effect of aequeous extract of propolis on Trichomonas gallinae, in vitro. Assiut Vet. Med. J. 2016, 62, 82–88. [Google Scholar] [CrossRef]
- Petrilla, J.; Mátis, G.; Molnár, A.; Jerzsele, Á.; Pál, L.; Gálfi, P.; Neogrády, Z.; Dublecz, K. A butirát antibakteriális hatékonyságának in vitro vizsgálata különféle Campylobacter jejuni törzseken. Magy. Állatorvosok Lapja 2021, 143, 57–64. [Google Scholar]
- Pomothy, J.M.; Barna, R.F.; Gere, E. The Effects of the Rosmarinic Acid in Livestock Animals: Literature Review. Magy. Állatorvosok Lapja 2020, 142, 567–576. [Google Scholar]
- Skoskiewicz-Malinowska, K.; Kaczmarek, U.; Malicka, B.; Walczak, K.; Zietek, M. Application of Chitosan and Propolis in Endodontic Treatment: A Review. Mini Rev. Med. Chem. 2017, 17, 410–434. [Google Scholar] [CrossRef] [PubMed]
- Kizerwetter-Świda, M.; Chrobak-Chmiel, D.; Rzewuska, M.; Antosiewicz, A.; Dolka, B.; Ledwoń, A.; Czujkowska, A.; Binek, M. Genetic characterization of coagulase-positive staphylococci isolated from healthy pigeons. Pol. J. Vet. Sci. 2015, 18, 627–634. [Google Scholar] [CrossRef] [PubMed]
- De Vasconcellos, H.V.G.; Silva, K.F.B.; Montenegro, H.; Miguel, C.B.; Tizioto, P.; Agostinho, F.; Araújo, M.C.; Ribas, R.M.; da Silva, M.V.; Soares, S.d.C.; et al. Staphylococcus aureus and Enterococcus faecium isolated from pigeon droppings (Columba livia) in the external environment close to hospitals. Rev. Soc. Bras. Med. Trop. 2022, 55, e0353. [Google Scholar] [CrossRef]
- Karim, S.J.I.; Islam, M.; Sikder, T.; Rubaya, R.; Halder, J.; Alam, J. Multidrug-resistant Escherichia coli and Salmonella spp. isolated from pigeons. Vet. World 2020, 13, 2156–2165. [Google Scholar] [CrossRef]
- Stenkat, J.; Krautwald-Junghanns, M.-E.; Schmidt, V. Causes of morbidity and mortality in free-living birds in an urban environment in Germany. EcoHealth 2013, 10, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Pasmans, F.; Baert, K.; Martel, A.; Bousquet-Melou, A.; Lanckriet, R.; De Boever, S.; Van Immerseel, F.; Eeckhaut, V.; de Backer, P.; Haesebrouck, F. Induction of the carrier state in pigeons infected with Salmonella enterica subspecies enterica serovar typhimurium PT99 by treatment with florfenicol: A matter of pharmacokinetics. Antimicrob. Agents Chemother. 2008, 52, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Stabler, R.M. Trichomonas gallinae: A review. Exp. Parasitol. 1954, 3, 368–402. [Google Scholar] [CrossRef] [PubMed]
- Cunha, I.B.S.; Sawaya, A.C.H.F.; Caetano, F.M.; Shimizu, M.T.; Marcucci, M.C.; Drezza, F.T.; Povia, G.S.; Carvalho, P.d.O. Factors that influence the yield and composition of Brazilian propolis extracts. J. Braz. Chem. Soc. 2004, 15, 964–970. [Google Scholar] [CrossRef]
- Galeotti, F.; Maccari, F.; Fachini, A.; Volpi, N. Chemical Composition and Antioxidant Activity of Propolis Prepared in Different Forms and in Different Solvents Useful for Finished Products. Foods 2018, 7, 41. [Google Scholar] [CrossRef]
- Da Silva, L.A.L.; Pezzini, B.R.; Soares, L. Spectrophotometric determination of the total flavonoid content in Ocimum basilicum L. (Lamiaceae) leaves. Pharmacogn. Mag. 2015, 11, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Melvin, P. CLSI: Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Köt. CLSI Standards M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- James, H.J. CLSI: Methods for Determining Bactericidal Activity of Antimicrobial Agents, 1st ed.; Köt. CLSI Standards M26; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 1999. [Google Scholar]
- Gunetti, M.; Castiglia, S.; Rustichelli, D.; Mareschi, K.; Sanavio, F.; Muraro, M.; Signorino, E.; Castello, L.; Ferrero, I.; Fagioli, F. Validation of analytical methods in GMP: The disposable Fast Read 102® device, an alternative practical approach for cell counting. J. Transl. Med. 2012, 10, 112. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Ye, S.-R.; Ting, C.; Yu, Y.-H. Antibacterial activity of propolins from Taiwanese green propolis. J. Food Drug Anal. 2018, 26, 761–768. [Google Scholar] [CrossRef]
- Wojtyczka, R.D.; Dziedzic, A.; Idzik, D.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Smoleń-Dzirba, J.; Stojko, J.; Sajewicz, M.; Wąsik, T.J. Susceptibility of Staphylococcus aureus clinical isolates to propolis extract alone or in combination with antimicrobial drugs. Molecules 2013, 18, 9623–9640. [Google Scholar] [CrossRef] [PubMed]
- Suleman, T.; van Vuuren, S.; Sandasi, M.; Viljoen, A.M. Antimicrobial activity and chemometric modelling of South African propolis. J. Appl. Microbiol. 2015, 119, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Almeida, E.T.; da Silva, M.C.D.; Oliveira, J.M.D.S.; Kamiya, R.U.; Arruda, R.E.D.S.; Vieira, D.A.; Silva, V.d.C.; Escodro, P.B.; Basílio-Júnior, I.D.; do Nascimento, T.G. Chemical and microbiological characterization of tinctures and microcapsules loaded with Brazilian red propolis extract. J. Pharm. Anal. 2017, 7, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Devequi-Nunes, D.; Machado, B.A.S.; Barreto, G.d.A.; Rebouças Silva, J.; da Silva, D.F.; da Rocha, J.L.C.; Brandão, H.N.; Borges, V.M.; Umsza-Guez, M.A. Chemical characterization and biological activity of six different extracts of propolis through conventional methods and supercritical extraction. PLoS ONE 2018, 13, e0207676. [Google Scholar] [CrossRef]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The Anti-Staphylococcal Potential of Ethanolic Polish Propolis Extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.P.D.; Machado, B.A.S.; Barreto, G.d.A.; Costa, S.S.; Andrade, L.N.; Amaral, R.G.; Carvalho, A.A.; Padilha, F.F.; Barbosa, J.D.V.; Umsza-Guez, M.A. Antioxidant, antimicrobial, antiparasitic, and cytotoxic properties of various Brazilian propolis extracts. PLoS ONE 2017, 12, e0172585. [Google Scholar] [CrossRef]
- El Menyiy, N.; Bakour, M.; El Ghouizi, A.; El Guendouz, S.; Lyoussi, B. Influence of Geographic Origin and Plant Source on Physicochemical Properties, Mineral Content, and Antioxidant and Antibacterial Activities of Moroccan Propolis. Int. J. Food Sci. 2021, 2021, 5570224. [Google Scholar] [CrossRef]
- Touzani, S.; Imtara, H.; Katekhaye, S.; Mechchate, H.; Ouassou, H.; Alqahtani, A.S.; Noman, O.M.; Nasr, F.A.; Fearnley, H.; Fearnley, J.; et al. Determination of Phenolic Compounds in Various Propolis Samples Collected from an African and an Asian Region and Their Impact on Antioxidant and Antibacterial Activities. Molecules 2021, 26, 4589. [Google Scholar] [CrossRef]
- Nazeri, R.; Ghaiour, M.; Abbasi, S. Evaluation of Antibacterial Effect of Propolis and its Application in Mouthwash Production. Front. Dent. 2019, 16, 1–12. [Google Scholar] [CrossRef]
- Jafarzadeh Kashi, T.S.; Kasra Kermanshahi, R.; Erfan, M.; Vahid Dastjerdi, E.; Rezaei, Y.; Tabatabaei, F.S. Evaluating the In-vitro Antibacterial Effect of Iranian Propolis on Oral Microorganisms. Iran. J. Pharm. Res. IJPR 2011, 10, 363–368. [Google Scholar] [PubMed]
- Mavri, A.; Abramovič, H.; Polak, T.; Bertoncelj, J.; Jamnik, P.; Smole Možina, S.; Jeršek, B. Chemical Properties and Antioxidant and Antimicrobial Activities of Slovenian Propolis. Chem. Biodivers. 2012, 9, 1545–1558. [Google Scholar] [CrossRef]
- Ristivojević, P.; Dimkić, I.; Trifković, J.; Berić, T.; Vovk, I.; Milojković-Opsenica, D.; Stanković, S. Antimicrobial Activity of Serbian Propolis Evaluated by Means of MIC, HPTLC, Bioautography and Chemometrics. PLoS ONE 2016, 11, e0157097. [Google Scholar] [CrossRef]
- Nina, N.; Feresin, G.; Giménez, A.; Capusiri, E.; Schmeda-Hirschmann, G. Antibacterial and Leishmanicidal Activity of Bolivian Propolis. Lett. Appl. Microbiol. 2016, 62, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Nina, N.; Quispe, C.; Jiménez-Aspee, F.; Theoduloz, C.; Feresín, G.E.; Lima, B.; Leiva, E.; Schmeda-Hirschmann, G. Antibacterial Activity, Antioxidant Effect and Chemical Composition of Propolis from the Región del Maule, Central Chile. Molecules 2015, 20, 18144–18167. [Google Scholar] [CrossRef]
- Regueira, M.S.; Tintino, S.R.; da Silva, A.R.P.; Costa, M.d.S.; Boligon, A.A.; Matias, E.F.F.; de Queiroz Balbino, V.; Menezes, I.R.A.; Melo Coutinho, H.D. Seasonal variation of Brazilian red propolis: Antibacterial activity, synergistic effect and phytochemical screening. Food Chem. Toxicol. 2017, 107, 572–580. [Google Scholar] [CrossRef]
- Popova, M.; Dimitrova, R.; Al-Lawati, H.T.; Tsvetkova, I.; Najdenski, H.; Bankova, V. Omani propolis: Chemical profiling, antibacterial activity and new propolis plant sources. Chem. Cent. J. 2013, 7, 158. [Google Scholar] [CrossRef]
- Sforcin, J.M.; Fernandes, A.; Lopes, C.A.; Bankova, V.; Funari, S.R. Seasonal effect on Brazilian propolis antibacterial activity. J. Ethnopharmacol. 2000, 73, 243–249. [Google Scholar] [CrossRef]
- Campos, J.F.; dos Santos, U.P.; dos Santos da Rocha, P.; Damião, M.J.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Estevinho, L.M.; de Picoli Souza, K.; dos Santos, E.L. Antimicrobial, Antioxidant, Anti-Inflammatory, and Cytotoxic Activities of Propolis from the Stingless Bee Tetragonisca fiebrigi (Jataí). Evid.-Based Complement. Alternat. Med. 2015, 2015, 296186. [Google Scholar] [CrossRef]
- Hegazi, A.G.; Hady, F.K.A.E.; Allah, F.A.M.A. Chemical Composition and Antimicrobial Activity of European Propolis. Z. Naturforschung C 2000, 55, 70–75. [Google Scholar] [CrossRef]
- Al-Waili, N.; Al-Ghamdi, A.; Ansari, M.J.; Al-Attal, Y.; Salom, K. Synergistic effects of honey and propolis toward drug multi-resistant Staphylococcus aureus, Escherichia coli and Candida albicans isolates in single and polymicrobial cultures. Int. J. Med. Sci. 2012, 9, 793–800. [Google Scholar] [CrossRef]
- Bezerra, C.R.F.; Borges, K.R.A.; Alves, R.d.N.S.; Teles, A.M.; Rodrigues, I.V.P.; da Silva, M.A.C.N.; Nascimento, M.d.D.S.B.; Bezerra, G.F.d.B. Highly efficient antibiofilm and antifungal activity of green propolis against Candida species in dentistry materials. PLoS ONE 2020, 15, e0228828. [Google Scholar] [CrossRef] [PubMed]
- Ota, C.; Unterkircher, C.; Fantinato, V.; Shimizu, M.T. Antifungal activity of propolis on different species of Candida. Mycoses 2001, 44, 375–378. [Google Scholar] [CrossRef] [PubMed]
- AL-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial Activities of European Propolis Collected from Various Geographic Origins Alone and in Combination with Antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef]
- Domán, M.; Vásárhelyi, B.; Balka, G.; Jantyik, T.; Laukó, T.; Bányai, K.; Makrai, L. Esophageal Mycosis in Hungarian Goose and Duck Flocks. Magy. Állatorvosok Lapja 2021, 143, 667–675. [Google Scholar]
- Boisard, S.; Le Ray, A.-M.; Landreau, A.; Kempf, M.; Cassisa, V.; Flurin, C.; Richomme, P. Antifungal and Antibacterial Metabolites from a French Poplar Type Propolis. Evid.-Based Complement. Altern. Med. 2015, 2015, 319240. [Google Scholar] [CrossRef] [PubMed]
- Falcão, S.I.; Vale, N.; Cos, P.; Gomes, P.; Freire, C.; Maes, L.; Vilas-Boas, M. In Vitro Evaluation of Portuguese Propolis and Floral Sources for Antiprotozoal, Antibacterial and Antifungal Activity. Phytother. Res. 2014, 28, 437–443. [Google Scholar] [CrossRef]
- De Oliveira Dembogurski, D.S.; Silva Trentin, D.; Boaretto, A.G.; Rigo, G.V.; da Silva, R.C.; Tasca, T.; Macedo, A.J.; Carollo, C.A.; Silva, D.B. Brown propolis-metabolomic innovative approach to determine compounds capable of killing Staphylococcus aureus biofilm and Trichomonas vaginalis. Food Res. Int. 2018, 111, 661–673. [Google Scholar] [CrossRef]
- Sena-Lopes, Â.; Bezerra, F.S.B.; das Neves, R.N.; de Pinho, R.B.; Silva, M.T.d.O.; Savegnago, L.; Collares, T.; Seixas, F.; Begnini, K.; Henriques, J.A.P.; et al. Chemical composition, immunostimulatory, cytotoxic and antiparasitic activities of the essential oil from Brazilian red propolis. PLoS ONE 2018, 13, e0191797. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Bacterial Strain | Number of Strains | Source of Samples |
|---|---|---|
| Staphylococcus aureus | 2 strains + 2 strains * | nasal cavity and egg |
| Staphylococcus delphini | 2 strains | respiratory tract and liver |
| Staphylococcus sciuri | 2 strains | respiratory and intestinal tract |
| Enterococcus gallinarum | 1 strain + 3 strains * | conjunctiva |
| Enterococcus columbae | 2 strains | respiratory and intestinal tract |
| Enterococcus hirae | 1 strain | respiratory tract |
| Enterococcus cecorum | 1 strain | respiratory tract |
| Escherichia coli | 7 strains + 1 ATCC strain * | conjunctiva, intestinal tract, liver |
| Salmonella enterica | 8 strains | intestinal tract, liver, joint, testis |
| Candida albicans | 8 strains * | skin |
| Strain | Ethanol Extract | 96% | 90% | 80% | 70% | 60% | |
|---|---|---|---|---|---|---|---|
| µg/mL | |||||||
| Staphylococcus spp. (n = 8) | MIC-range | µg/mL | 3.125–50 | 1.56–400 | 25–400 | 50–400 | 50–400 |
| MIC50 | 12.5 | 6.25 | 100 | 100 | 100 | ||
| MIC90 | 25 | 50 | 100 | 200 | 100 | ||
| MEC-range | 12.5–100 | 12.5–3260 | 200–3260 | 200–3260 | 200–3260 | ||
| MEC50 | 25 | 50 | 200 | 200 | 400 | ||
| MEC90 | 100 | 50 | 400 | 800 | 400 | ||
| Enterococcus spp. (n = 8) | MIC-range | 6.25–50 | 1.56–12.5 | 100–400 | 100–200 | 50–200 | |
| MIC50 | 25 | 12.5 | 200 | 200 | 200 | ||
| MIC90 | 50 | 12,5 | 400 | 200 | 200 | ||
| MEC-range | 12.5–100 | 12.5–50 | 200–800 | 100–400 | 200–400 | ||
| MEC50 | 100 | 50 | 400 | 200 | 200 | ||
| MEC90 | 100 | 50 | 800 | 400 | 400 | ||
| Candida albicans (n = 8) | MIC-range | 1.56–50 | 400–400 | 25–400 | 100–400 | 50–200 | |
| MIC50 | 25 | 400 | 100 | 100 | 100 | ||
| MIC90 | 50 | 400 | 200 | 200 | 200 | ||
| MEC-range | 3.125–50 | 400–400 | 100–800 | 100–800 | 100–400 | ||
| MEC50 | 50 | 400 | 200 | 200 | 200 | ||
| MEC90 | 50 | 400 | 400 | 200 | 200 | ||
| Ethanol Concentration | 96% | 90% | 80% | 70% | 60% | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MEC | MIC | MEC | MIC | MEC | MIC | MEC | MIC | MEC | |||
| Staphylococcus spp. | 96% | MIC | - | - | - | - | - | - | - | - | ||
| MEC | - | 0.2200 * | - | <0.0010 | - | <0.0010 | - | <0.0010 | ||||
| 90% | MIC | 0.9610 * | - | - | - | - | - | - | - | |||
| MEC | - | - | - | <0.0010 | - | <0.0010 | - | <0.0010 | ||||
| 80% | MIC | <0.0001 | - | <0.0001 | - | - | - | - | - | |||
| MEC | - | - | - | - | - | 0.9980 * | - | 0.9980 * | ||||
| 70% | MIC | <0.0001 | - | <0.0001 | - | 0.9910 * | - | - | - | |||
| MEC | - | - | - | - | - | - | - | 1.0000 * | ||||
| 60% | MIC | <0.0001 | - | <0.0001 | - | 1.0000 * | - | 0.9910 * | - | |||
| MEC | - | - | - | - | - | - | - | - | ||||
| Enterococcus spp. | 96% | MIC | - | - | - | - | - | - | - | - | ||
| MEC | - | 0.1950* | - | <0.0010 | - | <0.0010 | - | <0.0010 | ||||
| 90% | MIC | <0.0001 | - | - | - | - | - | - | - | |||
| MEC | - | - | - | <0.0010 | - | <0.0010 | - | <0.0010 | ||||
| 80% | MIC | <0.0001 | - | <0.0001 | - | - | - | - | - | |||
| MEC | - | - | - | - | - | 0.0861 * | - | 0.3734 * | ||||
| 70% | MIC | <0.0001 | - | <0.0001 | - | 0.9950 * | - | - | - | |||
| MEC | - | - | - | - | - | - | - | 0.9520 * | ||||
| 60% | MIC | <0.0001 | - | <0.0001 | - | 0.7810 * | - | 0.9410 * | - | |||
| MEC | - | - | - | - | - | - | - | - | ||||
| Candida albicans | 96% | MIC | - | - | - | - | - | - | - | - | ||
| MEC | <0.0010 | - | <0.0010 | - | <0.0010 | - | <0.0010 | |||||
| 90% | MIC | <0.0010 | - | - | - | - | - | - | - | |||
| MEC | - | - | - | 0.4530* | - | 0.0583 * | - | <0.0010 | ||||
| 80% | MIC | <0.0010 | - | <0.0010 | - | - | - | - | - | |||
| MEC | - | - | - | - | - | 0.8547 * | - | 0.8547 * | ||||
| 70% | MIC | <0.0010 | - | 0.0041 | - | 0.9984 * | - | - | - | |||
| MEC | - | - | - | - | - | - | - | 1.0000 * | ||||
| 60% | MIC | <0.0010 | - | 0.0014 | - | 1.0000 * | - | 0.9984 * | - | |||
| MEC | - | - | - | - | - | - | - | - | ||||
| MEC | 96% | 90% | 80% | 70% | 60% | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 24-h | 48-h | 24-h | 48-h | 24-h | 48-h | 24-h | 48-h | 24-h | 48-h | ||
| 1 | 96% | - | <0.0010 | - | <0.0010 | - | <0.0010 | - | <0.0010 | ||
| 2 | - | <0.0010 | - | <0.0010 | - | <0.0010 | - | <0.0010 | |||
| 3 | - | - | - | - | - | - | - | - | |||
| 1 | 90% | <0.0010 | - | - | 0.9999 * | - | 0.3809 * | - | 0.0226 | ||
| 2 | <0.0010 | - | - | 0.1075 * | - | 0.3097 * | - | 0.0479 | |||
| 3 | <0.0010 | - | - | - | - | - | - | ||||
| 1 | 80% | <0.0010 | - | <0.1074 * | - | - | 0.2951 * | - | 0.0142 | ||
| 2 | <0.0010 | - | 0.1070 * | - | - | 0.9842 * | - | 0.9975 * | |||
| 3 | <0.0010 | - | 0.1074 * | - | - | - | - | - | |||
| 1 | 70% | <0.0010 | - | 0.3098 * | - | 0.9842 * | - | - | 0.7193 * | ||
| 2 | <0.0010 | - | 0.3100 * | - | 0.9840 * | - | - | 0.9125 * | |||
| 3 | <0.0010 | - | 0.3097 * | - | 0.9842 * | - | - | - | |||
| 1 | 60% | <0.0010 | - | 0.0479 | - | 0.9975 * | - | 0.9125 * | - | ||
| 2 | <0.0010 | - | 0.0480 | - | 0.9980 * | - | 0.9120 * | - | |||
| 3 | <0.0010 | - | 0.0478 | - | 0.9975 * | - | 0.9125 * | - | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerek, Á.; Csanády, P.; Tuska-Szalay, B.; Kovács, L.; Jerzsele, Á. In Vitro Efficacy of Hungarian Propolis against Bacteria, Yeast, and Trichomonas gallinae Isolated from Pigeons—A Possible Antibiotic Alternative? Resources 2023, 12, 101. https://doi.org/10.3390/resources12090101
Kerek Á, Csanády P, Tuska-Szalay B, Kovács L, Jerzsele Á. In Vitro Efficacy of Hungarian Propolis against Bacteria, Yeast, and Trichomonas gallinae Isolated from Pigeons—A Possible Antibiotic Alternative? Resources. 2023; 12(9):101. https://doi.org/10.3390/resources12090101
Chicago/Turabian StyleKerek, Ádám, Péter Csanády, Barbara Tuska-Szalay, László Kovács, and Ákos Jerzsele. 2023. "In Vitro Efficacy of Hungarian Propolis against Bacteria, Yeast, and Trichomonas gallinae Isolated from Pigeons—A Possible Antibiotic Alternative?" Resources 12, no. 9: 101. https://doi.org/10.3390/resources12090101
APA StyleKerek, Á., Csanády, P., Tuska-Szalay, B., Kovács, L., & Jerzsele, Á. (2023). In Vitro Efficacy of Hungarian Propolis against Bacteria, Yeast, and Trichomonas gallinae Isolated from Pigeons—A Possible Antibiotic Alternative? Resources, 12(9), 101. https://doi.org/10.3390/resources12090101