Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Experimental Setup
2.2. Biomethane Production Experiments and Data Analyses
2.2.1. Batch Digestion Test 1
2.2.2. Batch Digestion Test 2
2.2.3. Batch Digestion Test 3
2.2.4. Batch Digestion Test 4
2.3. Biomass and Data Analysis
3. Results
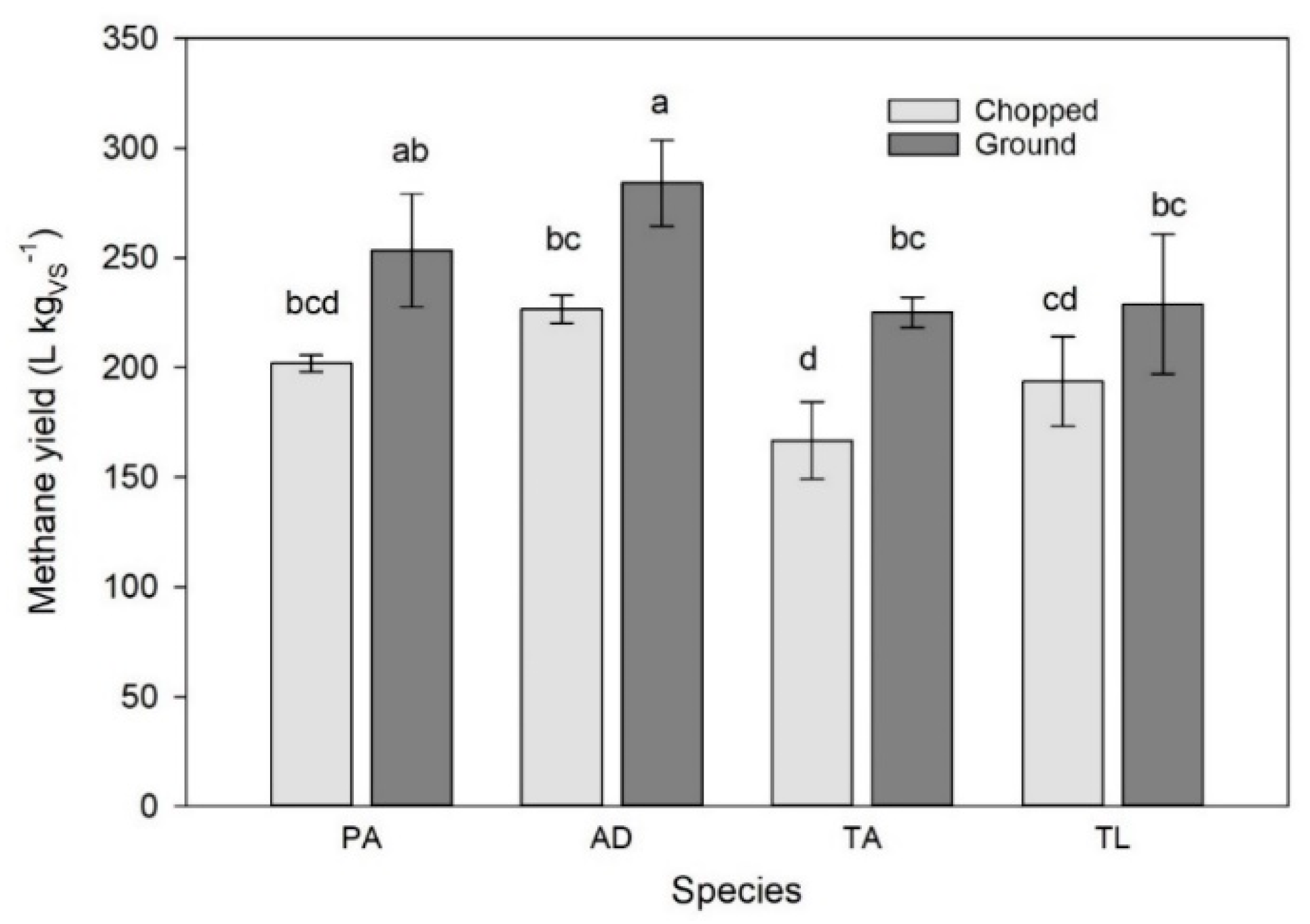
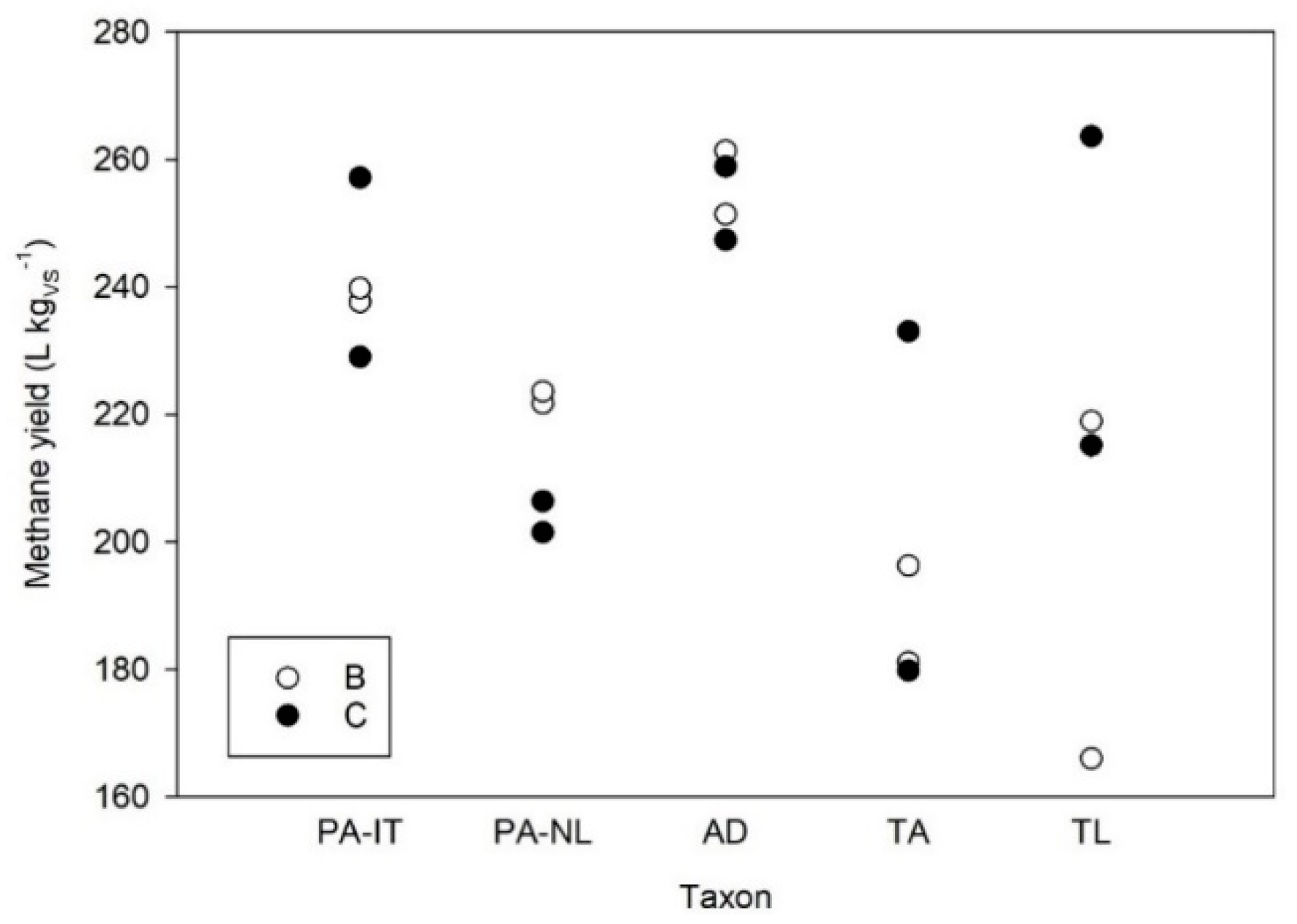
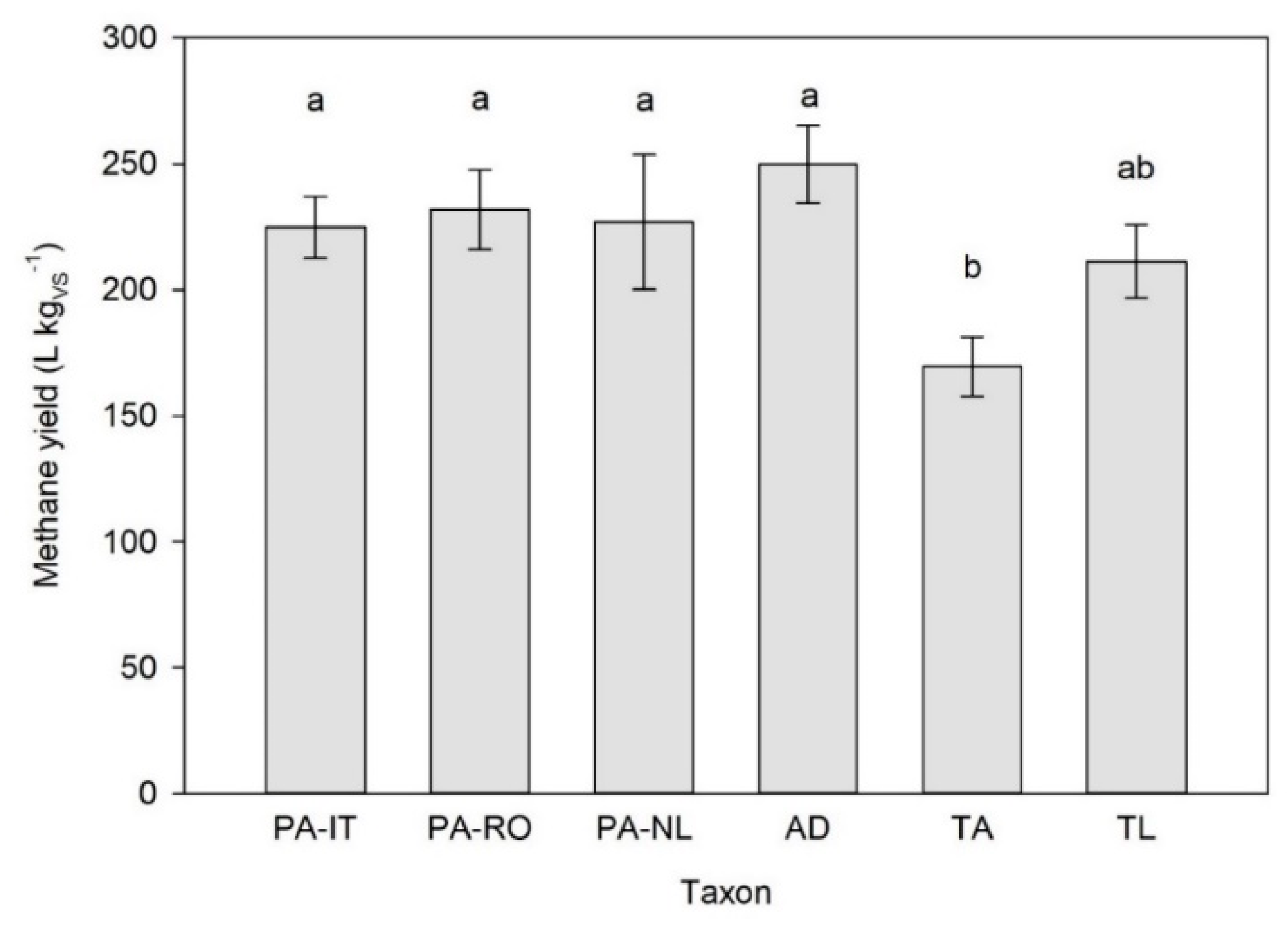
3.1. Batch Digestion Test 1
3.2. Batch Digestion Test 2
3.3. Batch Digestion Test 3
3.4. Batch Digestion Test 4
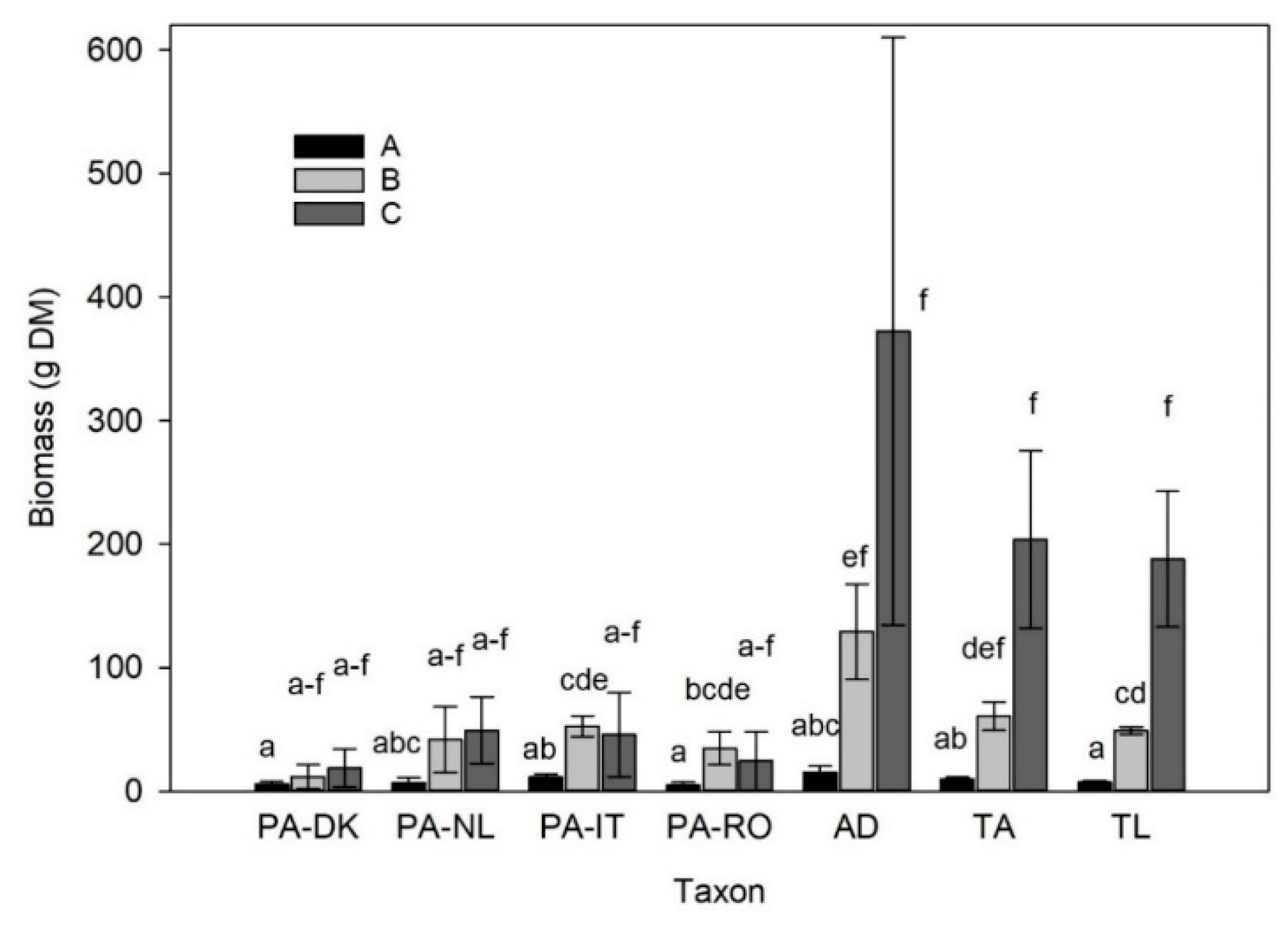
3.5. Biomass
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ward, A.J.; Hobbs, P.J.; Holliman, P.J.; Jones, D.L. Optimisation of the anaerobic digestion of agricultural resources. Bioresour. Technol. 2008, 99, 7928–7940. [Google Scholar] [CrossRef]
- Karlsson, N.P.E.; Halila, F.; Mattsson, M.; Hoveskog, M. Success factors for agricultural biogas production in Sweden: A case study of business model innovation. J. Clean. Prod. 2017, 142, 2925–2934. [Google Scholar] [CrossRef]
- Verotti, M.; Servadio, P.; Bergonzoli, S. Biogas upgrading and utilization from ICEs towards stationary molten carbonate fuel cell systems. Int. J. Green Energy 2016, 13, 655–664. [Google Scholar] [CrossRef]
- Karki, S.; Elsgaard, L.; Lærke, P.E. Effect of reed canary grass cultivation on greenhouse gas emission from peat soil at controlled rewetting. Biogeosciences 2015, 12, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Wichtmann, W.; Schröder, C.; Joosten, H. Paludiculture—Productive Use of Wet Peatlands; Schweizerbart Science Publishers: Stuttgart, Germany, 2016. [Google Scholar]
- Weisner, S.E.B.; Johannesson, K.; Thiere, G.; Svengren, H.; Ehde, P.M.; Tonderski, K.S. National large-scale wetland creation in agricultural areas—Potential versus realized effects on nutrient transports. Water 2016, 8, 544. [Google Scholar] [CrossRef]
- Joosten, H.; Tanneberger, F.; Moen, A. (Eds.) Mires and Peatlands of Europe—Status, Distribution and Conservation; Schweitzerbart Science Publishers: Stuttgart, Germany, 2017; 780p. [Google Scholar]
- Wichtmann, W. Use of biomass from rewetted peatlands (in German). Greifswald. Geogr. Arb. 2003, 31, 43–54. [Google Scholar]
- Gniffke, P.; Strogies, M. (Eds.) National Inventory Report for the German Greenhouse Gas Inventory 1990–2014; Submission under the United Nations Framework Convention on Climate Change and the Kyoto Protocol; Umweltbundesamt: Dessau-Roßlau, Germany, 2016.
- Lambertini, C.; Sorrell, B.K.; Riis, T.; Olesen, B.; Brix, H. Exploring the borders of European Phragmites within a cosmopolitan genus. AoB Plants 2012, pls020. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.Y.; Lambertini, C.; Li, X.Z.; Meyerson, L.A.; Brix, H. Invasion of old world Phragmites australis in the new world: Precipitation and temperature patterns combined with human influences redesign the invasive niche. Glob. Chang. Biol. 2013, 19, 3406–3422. [Google Scholar] [PubMed]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.Y.; Guo, X.; Hazelton, E.L.G.; Kettenring, K.M.; et al. Cosmopolitan Species as Models for Ecophysiological Responses to Global Change: The Common Reed Phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef] [Green Version]
- Brix, H. Do macrophytes play a role in constructed treatment wetlands? Water Sci. Technol. 1997, 35, 11–17. [Google Scholar] [CrossRef]
- Vymazal, J.; Kröpfelova, L. Growth of Phragmites australis and Phalaris arundinacea in constructed wetlands for wastewater treatment in the Czech Republic. Ecol. Eng. 2005, 25, 606–621. [Google Scholar] [CrossRef]
- Weisner, S.E.B.; Thiere, G. Effects of vegetation state on biodiversity and nitrogen retention in created wetlands: A test of the biodiversity–ecosystem functioning hypothesis. Freshw. Biol. 2010, 55, 387–396. [Google Scholar] [CrossRef]
- Ren, L.; Eller, F.; Lambertini, C.; Guo, W.Y.; Sorrell, B.K.; Brix, H. Minimum Fe requirement and toxic tissue concentration of Fe in Phragmites australis: A tool for alleviating Fe-deficiency in constructed wetlands. Ecol. Eng. 2018, 118, 152–160. [Google Scholar] [CrossRef]
- Tho, B.T.; Sorrell, B.K.; Lambertini, C.; Eller, F.; Brix, H. Phragmites australis: How do genotypes of different phylogeographic origins differ from their invasive genotypes in growth, nitrogen allocation and gas exchange? Biol. Invasions 2016, 18, 2563–2576. [Google Scholar] [CrossRef]
- Eller, F.; Brix, H. Different genotypes of Phragmites australis show distinct phenotypic plasticity in response to nutrient availability and temperature. Aquat. Bot. 2012, 103, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Eller, F.; Lambertini, C.; Nguyen, L.X.; Achenbach, L.; Brix, H. Interactive effects of elevated temperature and CO2 on two phylogeographically distinct clones of common reed (Phragmites australis). AoB Plants 2013, 5, pls051. [Google Scholar] [CrossRef]
- Nguyen, L.X.; Lambertini, C.; Sorrell, B.K.; Eller, F.; Achenbach, L.; Brix, H. Photosynthesis of co-existing Phragmites haplotypes in their non-native range: Are characteristics determined by adaptations derived from their native origin? AoB Plants 2013, 5, plt016. [Google Scholar] [CrossRef] [Green Version]
- Brix, H.; Sorrell, B.K.; Orr, P.T. Internal Pressurization and Convective Gas-Flow in Some Emergent Fresh-Water Macrophytes. Limnol. Oceanogr. 1992, 37, 1420–1433. [Google Scholar] [CrossRef]
- Sorrell, B.K.; Brix, H. Effects of water vapour pressure deficit and stomatal conductance on photosynthesis, internal pressurization and convective flow in three emergent wetland plants. Plant Soil 2003, 253, 71–79. [Google Scholar] [CrossRef]
- Jennewein, S.P.; Gilbert, R.A.; Rowland, D.L.; Wright, A.L.; Glaz, B.; Bennett, J.M. Four Biofuel Species’ Responses to Periodic Flooding and High Water Tables on a Florida Histosol. BioEnergy Res. 2017, 10, 688–699. [Google Scholar] [CrossRef]
- de Gannes, V.; Eudoxie, G.; Hickey, W.J. Feedstock carbon influence on compost biochemical stability and maturity. Compost. Sci. Util. 2018, 26, 59–70. [Google Scholar] [CrossRef]
- Dioha, I.J.; Ikeme, C.; Nafi, T.; Soba, N.I.; Mbs, Y. Effect of carbon to nitrogen ratio on biogas production. Int. Res. J. Nat. Sci. 2013, 1, 1–10. [Google Scholar]
- Giannini, V.; Oehmke, C.; Silvestri, N.; Wichtmann, W.; Dragoni, F.; Bonari, E. Combustibility of biomass from perennial crops cultivated on a rewetted Mediterranean peatland. Ecol. Eng. 2016, 97, 157–169. [Google Scholar] [CrossRef]
- Kupryś-Caruk, M.; Podlaski, S.; Kotyrba, D. Influence of double-cut harvest system on biomass yield, quality and biogas production from C4 perennial grasses. Biomass Bioenergy 2019, 130, 105376. [Google Scholar] [CrossRef]
- Lu, C.; Tian, H. Global nitrogen and phosphorus fertilizer use for agriculture production in the past half century: Shifted hot spots and nutrient imbalance. Earth Syst. Sci. Data 2017, 9, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Akritas, M.G. The rank transformation method in some two-factor designs. J. Am. Stat. Assoc. 1990, 85, 73–78. [Google Scholar] [CrossRef]
- Ragaglini, G.; Dragoni, F.; Simone, M.; Bonari, E. Suitability of giant reed (Arundo donax L.) for anaerobic digestion: Effect of harvest time and frequency on the biomethane yield potential. Bioresour. Technol. 2014, 152, 107–115. [Google Scholar] [CrossRef]
- Nkemka, V.N.; Gilroyed, B.; Yanke, J.; Gruninger, R.; Vedres, D.; McAllister, T.; Hao, X. Bioaugmentation with an anaerobic fungus in a two-stage process for biohydrogen and biogas production using corn silage and cattail. Bioresour. Technol. 2015, 185, 79–88. [Google Scholar] [CrossRef]
- Lizasoain, J.; Rincón, M.; Theuretzbacher, F.; Enguídanos, R.; Nielsen, P.J.; Potthast, A.; Zweckmair, T.; Gronauer, A.; Bauer, A. Biogas production from reed biomass: Effect of pretreatment using different steam explosion conditions. Biomass Bioenergy 2016, 95, 84–91. [Google Scholar] [CrossRef]
- Boyd, C.E. Further Studies on Productivity, Nutrient and Pigment Relationships in Typha latifolia Populations. Bull. Torrey Bot. Club 1971, 98, 144–150. [Google Scholar] [CrossRef]
- Gacia, E.; Bernal, S.; Nikolakopoulou, M.; Carreras, E.; Morgado, L.; Ribot, M.; Isnard, M.; Sorolla, A.; Sabater, F.; Martí, E. The role of helophyte species on nitrogen and phosphorus retention from wastewater treatment plant effluents. J. Environ. Manag. 2019, 252, 109585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbachová-Vojtíšková, L.; Tylová, E.; Soukup, A.; Novická, H.; Votrubová, O.; Lipavská, H.; Čížková, H. Influence of nutrient supply on growth, carbohydrate, and nitrogen metabolic relations in Typha angustifolia. Environ. Exp. Bot. 2006, 57, 246–257. [Google Scholar] [CrossRef]
- Ren, L.; Eller, F.; Lambertini, C.; Guo, W.Y.; Brix, H.; Sorrell, B.K. Assessing nutrient responses and biomass quality for selection of appropriate paludiculture crops. Sci. Total Environ. 2019, 664, 1150–1161. [Google Scholar] [CrossRef] [PubMed]
- Dragoni, F.; Giannini, V.; Ragaglini, G.; Bonari, E.; Silvestri, N. Effect of Harvest Time and Frequency on Biomass Quality and Biomethane Potential of Common Reed (Phragmites australis) Under Paludiculture Conditions. BioEnergy Res. 2017, 10, 1066–1078. [Google Scholar] [CrossRef]
- Günther, A.; Huth, V.; Jurasinski, G.; Glatzel, S. The effect of biomass harvesting on greenhouse gas emissions from a rewetted temperate fen. Glob. Chang. Biol. Bioenergy 2015, 7, 1092–1106. [Google Scholar] [CrossRef]
- Liu, D.; Wu, X.; Chang, J.; Gu, B.; Min, Y.; Ge, Y.; Shi, Y.; Xue, H.; Peng, C.; Wu, J. Constructed wetlands as biofuel production systems. Nat. Clim. Chang. 2012, 2, 190–194. [Google Scholar] [CrossRef]
- Granéli, W.; Weisner, S.E.B.; Sytsma, M.D. Rhizome dynamics and resource storage in Phragmites australis. Wetl. Ecol. Manag. 1992, 1, 239–247. [Google Scholar] [CrossRef]
- Baute, K.; Van Eerd, L.L.; Robinson, D.E.; Sikkema, P.H.; Mushtaq, M.; Gilroyed, B.H. Comparing the biomass yield and biogas potential of Phragmites australis with Miscanthus × giganteus and Panicum virgatum grown in Canada. Energies 2018, 11, 2198. [Google Scholar] [CrossRef] [Green Version]
- Paepatung, N.; Nopharatana, A.; Songkasiri, W. Bio-Methane Potential of Biological Solid Materials and Agricultural Wastes. Asian J. Energy Environ. 2009, 10, 19–27. [Google Scholar]
- Jagadabhi, P.S.; Kaparaju, P.; Rintala, J. Two-stage anaerobic digestion of tomato, cucumber, common reed and grass silage in leach-bed reactors and upflow anaerobic sludge blanket reactors. Bioresour. Technol. 2011, 102, 4726–4733. [Google Scholar] [CrossRef]
- Nkemka, V.N.; Murto, M. Two-stage anaerobic dry digestion of blue mussel and reed. Renew. Energy 2013, 50, 359–364. [Google Scholar] [CrossRef]
- Yang, L.; Li, Y. Anaerobic digestion of giant reed for methane production. Bioresour. Technol. 2014, 171, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Baldini Baldini, M.; da Borso, F.; Ferfuia, C.; Zuliani, F.; Danuso, F. Ensilage suitability and bio-methane yield of Arundo donax and Miscanthus × giganteus. Ind. Crops Prod. 2017, 95, 264–275. [Google Scholar] [CrossRef]
- Amon, T.; Amon, B.; Kryvoruchko, V.; Machmüller, A.; Hopfner-Sixt, K.; Bodiroza, V.; Hrbek, R.; Friedel, J.; Pötsch, E.; Wagentristl, H.; et al. Methane production through anaerobic digestion of various energy crops grown in sustainable crop rotations. Bioresour. Technol. 2007, 98, 3204–3212. [Google Scholar] [CrossRef]
- Mayer, F.; Gerin, P.A.; Noo, A.; Lemaigre, S.; Stilmant, D.; Schmit, T.; Leclech, N.; Ruelle, L.; Gennen, J.; von Francken-Welz, H.; et al. Assessment of energy crops alternative to maize for biogas production in the Greater Region. Bioresour. Technol. 2014, 166, 358–367. [Google Scholar] [CrossRef]
- Dubbe, D.R.; Garver, E.G.; Pratt, D.C. Production of cattail (Typha spp.) biomass in Minnesota, USA. Biomass 1988, 17, 79–104. [Google Scholar] [CrossRef]
- Risén, E.; Gregeby, E.; Tatarchenko, O.; Blidberg, E.; Malmström, M.E.; Welander, U.; Gröndahl, F. Assessment of biomethane production from maritime common reed. J. Clean. Prod. 2013, 53, 186–194. [Google Scholar] [CrossRef]
- Huhta, A. To cut or not to cut? The relationship between Common Reed, mowing and water quality. In Read Up on Reed; Southwest Finland Regional Environment Center: Turku, Finland, 2007; pp. 30–38. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fertilization (kg N ha−1 year−1) | N (g per pot per Week) | P (g per pot per Week) | K (g per pot per Week) | Amount of Stock Solution Added Per Pot Per Week (mL) |
|---|---|---|---|---|---|
| A | 0 | 0 | 0 | 0 | 0 |
| B | 75 | 0.1876 | 0.0224 | 0.1497 | 10 |
| C | 500 | 1.3132 | 0.1568 | 1.0479 | 70 |
| Species | Fertilization Treatment | Taxon Mean ± SD | |||
|---|---|---|---|---|---|
| A | B | C | |||
| P. australis | IT | 224.7 | 195.2 | 223.3 | 214 ± 17 a |
| NL | 200.1 | 208.0 | 236.1 | 215 ± 19 a | |
| RO | 229.6 | 233.5 | 233.1 | 232 ± 2 a | |
| DK | 223.8 | 223.9 | 243.7 | 231 ± 12 a | |
| A. donax | 245.8 | 252.1 | 256.6 | 252 ± 5 a | |
| T. angustifolia | 139.5 | 172.0 | 204.5 | 172 ± 33 b | |
| T. latifolia | 212.6 | 203.8 | 249.2 | 222 ± 24 a | |
| Fertilization treatment Mean ± SD | 211 ± 35 a | 213 ± 26 a | 235 ± 17 b | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eller, F.; Ehde, P.M.; Oehmke, C.; Ren, L.; Brix, H.; Sorrell, B.K.; Weisner, S.E.B. Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes. Resources 2020, 9, 57. https://doi.org/10.3390/resources9050057
Eller F, Ehde PM, Oehmke C, Ren L, Brix H, Sorrell BK, Weisner SEB. Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes. Resources. 2020; 9(5):57. https://doi.org/10.3390/resources9050057
Chicago/Turabian StyleEller, Franziska, Per Magnus Ehde, Claudia Oehmke, Linjing Ren, Hans Brix, Brian K. Sorrell, and Stefan E. B. Weisner. 2020. "Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes" Resources 9, no. 5: 57. https://doi.org/10.3390/resources9050057
APA StyleEller, F., Ehde, P. M., Oehmke, C., Ren, L., Brix, H., Sorrell, B. K., & Weisner, S. E. B. (2020). Biomethane Yield from Different European Phragmites australis Genotypes, Compared with Other Herbaceous Wetland Species Grown at Different Fertilization Regimes. Resources, 9(5), 57. https://doi.org/10.3390/resources9050057