1. Introduction
Baldness or androgenic alopecia (AGA) does not have many physically harmful effects to human health; however, it has a great impact on a person’s self-respect, mental health, and overall quality of life [
1,
2]. AGA is caused by aberrant hair follicle cycling (anagen to telogen stages) and miniaturization of hair follicles, which depends on the androgenic hormones (testosterone and dihydrotestosterone, DHT) [
3]. DHT is a male hormone product converted from testosterones by 5α-reductase that is more active than testosterone. Its androgenic effect can be reduced by blocking the conversion of testosterone to DHT. Therefore, anti-androgenic substances, which inhibit the 5α-reductase enzyme or bind between the androgen receptor and DHT, may be useful for preventing AGA [
3,
4]. With respect to the treatment of AGA, minoxidil (as a topical medication) and finasteride have been used. These synthetic medications, however, have a number of limitations. Their safety and effectiveness are questionable, and the amount of time needed to achieve their effect is long [
2,
3,
4]. Therefore, there is an interest in finding an alternative or new compound against hair loss, especially from natural plants and/or their active ingredients.
The ideal treatment for hair loss is a combination of 5α-reductase inhibitors with hair growth-promoting substances [
3]. Currently, natural extracts from several plants have been used for hair growth-promoting treatment in terms of 5α-reductase inhibitors, e.g.,
Carthamus tinctorius,
Clitoria ternatea,
Citrus hystrix,
Ipomoea aquatica,
Cassia siamea, and
Rhinacanthus nasutus, and in terms of hair growth-promoting substances, such as
Ginkgo biloba and
Lawsonai inermis [
4,
5]. Moreover, naturally derived compounds have been reported to impact hair growth improvement, including fatty acid or linolenic acid, phytoestrogens, and proanthocyanidins [
4]. Proanthocyanidins or procyanidin oligomers can promote proliferation of hair follicle cells and possess remarkable hair-cycle-converting activity from the telogen phase to the anagen phase in C3H mice [
6,
7], and have exhibited hair growth effects in clinical studies [
8,
9]. These substances can be found in various plants including grape seed, apple, barley, and pigmented rice [
6,
7,
10,
11].
Oryza sativa L. is consumed as a staple food by over half of the world’s population. Thailand is one of the world’s major producers of rice, with more than 93 rice varieties including pigmented and non-pigmented strains [
12]. The phytochemicals responsible for pigmentation in rice pericarp are mainly anthocyanins for black rice and proanthocyanidins for red or brown rice, which are considered a main resource in the context of natural health promotion. Rice is used in several traditional Asian treatments for cardiovascular disease, diabetes, inflammation, and as a skin and hair treatment [
12,
13,
14]. Historically, rice water obtained from the washing of rice has been used for traditional hair treatment in Asia, including Thailand, because it exhibits effects on hair, such as reducing surface friction, increasing hair elasticity, and inducing hair growth [
15]. According to these traditional Asian wisdoms for hair care, Thai pigmented rice containing proanthocyanidin is one of the important natural sources of active ingredients for hair-growth promoting product. However, the benefits of rice for hair are supported by scarce scientific studies and there is a lack of scientific literature unveiling the hair growth effects largely claimed by cosmetic and other related manufacturers. Therefore, the present study aimed to investigate the hair growth-promoting activities of Thai native red (Sang-Yod and Mun-Poo) and black (Black glutinous and Hom-Nil) pigmented rice extracts including in vitro 5α-reductase inhibition, hair growth-promoting activity in human hair matrix cells and in vivo hair-cycle-converting activity in C3H mice. Moreover, the safety of these extracts is evaluated via cytotoxicity and in vivo irritation tests in this study.
2. Materials and Methods
2.1. Plant Materials
Four native pigmented rice varieties (Oryza sativa L.) including two red rice varieties (Sang-Yod: SY, Mun-Poo: MP) and two black rice varieties (Black Glutinous: BG, Hom-Nil: HN) were used. MP, BG, and HN rice were procured from rice farmers in Northern Thailand whereas SY rice was harvested from Phatthalung, Southern Thailand. The voucher specimens of these rice (SY: MFLU00622, MP: MFLU00623, BG: MFLU00621, HN: MFLU00624) were deposited at a herbarium at Mae Fah Luang University Botanical Garden, Chiang Rai, Thailand. These grains were rinsed with absolute ethanol (Merck, Darmstadt, Germany) and dried with a hot air oven (Memmert, UFE600, Germany) at 45 °C for 48 h.
2.2. Laboratory Animals and Ethics Committee
Three adult male Spague-Dawley rats (Rattus norvegicus: aged ~8 weeks) and six adult male C3H/HeMlac mice (Mus musculus: aged ~8 weeks) from the National Laboratory Animal Center, Mahidol University were used. These animals were acclimatized to the laboratory environment for one week, where they were maintained in a controlled environment at a temperature of 25 °C, and with a diurnal period characterized by a 12:12 h light and dark cycle, with ad libitum food and water, at the Laboratory Animal Service Center, Prince of Songkla University until further study. The animal test in this study was guided by the principle of the 3Rs (replacement, reduction and refinement). Prior to the animal study, the study protocol was filed with the animal ethic committee for approval. All of the investigated methods used with animals that had been filed in the study protocol were approved by the ethics committee of the Prince of Songkla University with the registered approval no. MOE 0521.11/285.
2.3. Rice Extraction
Four native rice were extracted with 70% ethanol (Merck, Darmstadt, Germany) at a rice:solvent ratio of 1:10 (w/v) using the shaking method (Julabo, SW22, Darmstadt, Germany) at 150 rpm at room temperature for 6 h. All rice extracts were filtered with filter papers and solvents were removed with a rotary evaporator (Eyela, 0SB-2100, Beijing, China) at 45 °C. The samples were freeze-dried (Labconco, Freezone 6 plus, Kansas City, MO, USA) and kept at −20 °C.
2.4. Active Principle Contents
2.4.1. Total Phenolic Content (TPC)
Total phenolic content was determined using the Folin–Ciocalteu assay [
10]. Briefly, rice extracts were mixed with Folin–Ciocalteu reagent (Merck, Darmstadt, Germany) and 7.5% Na
2CO
3 (Merck, Darmstadt, Germany) for a 30-min incubation at ambient temperature. Absorbencies were determined at 765 nm using a microplate reader (Biotek, Epoch, Santa Clara, CA, USA). Gallic acid (Sigma, Livonia, MI, USA) was used as a reference standard. The
TPC was expressed as milligram gallic acid equivalents per gram extract (mg GAE/g extract).
2.4.2. Total Proanthocyanidin Content (TPAC)
Epicatechin (Sigma, Livonia, MI, USA) concentrated to 30–150 µg/mL was prepared in DMSO (RCI Labscan, Bangkok, Thailand) to generate a calibration curve (
r > 0.999).
TPAC was measured based on the vanillin–sulfuric acid assay [
10]. Prior to the absorbance reading at 500 nm, the sample solution was mixed with 1% vanillin (Sigma, USA) and 25% H
2SO
4 (Merck, Darmstadt, Germany) in MeOH (Merck, Darmstadt, Germany) for 15 min. Each sample was measured in triplicate. The
TPAC was expressed as milligram epicatechin equivalents per gram extract (mg ECE/g extract).
2.5. 5α-Reductase Inhibitory Activity
5α-reductase inhibitory activity was determined using the HPLC method as described by Kumar et al. [
5], with some modification. Briefly, a rat microsomal suspension was prepared by using the livers of Spague-Dawley rats. Livers were minced and homogenized in a buffer solution (1 mM dithiothreitol (Sigma, USA) and 0.32 M sucrose (Sigma, USA) in 0.02 M phosphate buffer (Merck, Darmstadt, Germany) at pH 6.5. The solution was centrifuged (Hermle, Z326K, Wehingen, Germany) at 4500×
g at 0 °C for 30 min (2 times). Then, the supernatants were collected and kept at −20 °C until they were used.
Rice extracts (0.2 mL in 50% ethanol) were mixed with 1.0 mL of 0.02 mM phosphate buffer (pH 6.5), 0.3 mL of 500 ppm testosterone (Sigma, USA) in 50% ethanol, and 1.0 mL of rat microsomal suspension. The mixture was reacted with 0.5 mL of 0.77 mg/mL NADPH (Sigma, USA) in phosphate buffer and then incubated at 37 °C for 30 min. Then, the reaction was stopped by adding 5.0 mL of dichloromethane (Merck, Germany) followed by adding 0.5 mL of 100 ppm propyl p-hydroxybenzoate (Sigma, USA) in 50% ethanol (as an internal standard). The organic phase was evaporated with a rotary evaporator. The residues were redissolved in 5.0 mL of methanol. An aliquot of 10 µL was injected into the HPLC system.
HPLC was performed on a Waters Alliance 2625 separations module. Chromatographic separation was performed using an AltimaTM column (C18; 250 mm × 4.6 mm; particle size, 5 µM) maintained at 40 °C. The mobile phase consisted of methanol and deionized water (65:35) with a flow rate of 1.0 mL/min. A UV detector at 242 nm was used to collect data.
To determine inhibitory activity, two special reactions must be completed: firstly, a complete reaction (rxn) containing 0.2% of 50% ethanol instead of the extract; secondly, an enzyme blank (ctrl) that receives 5.0 mL dichloromethane before the addition of NADPH, so that the conversion of testosterone into DHT does not occur. The percent inhibition was calculated using the peak area ratio (r) of testosterone relative to the internal standard following the equation: % Inhibition = [(rsample − rrxn)/(rctrl − rrxn)] × 100. The standard enzyme inhibitor was finasteride (Sigma, USA). With respect to the 5α-reductase inhibitory activity of finasteride, its IC50 was calculated as 0.32 μM. The correlation between the inhibitory activity (% Inhibition) and concentration of finasteride was expressed as: y = 147.2x + 2.2 (R2 = 0.999) with y representing percent inhibition and x representing the concentration of finasteride. All results were expressed as finasteride equivalent 5α-reductase inhibitory activity (FEA)/g extract.
2.6. Cytotoxicity
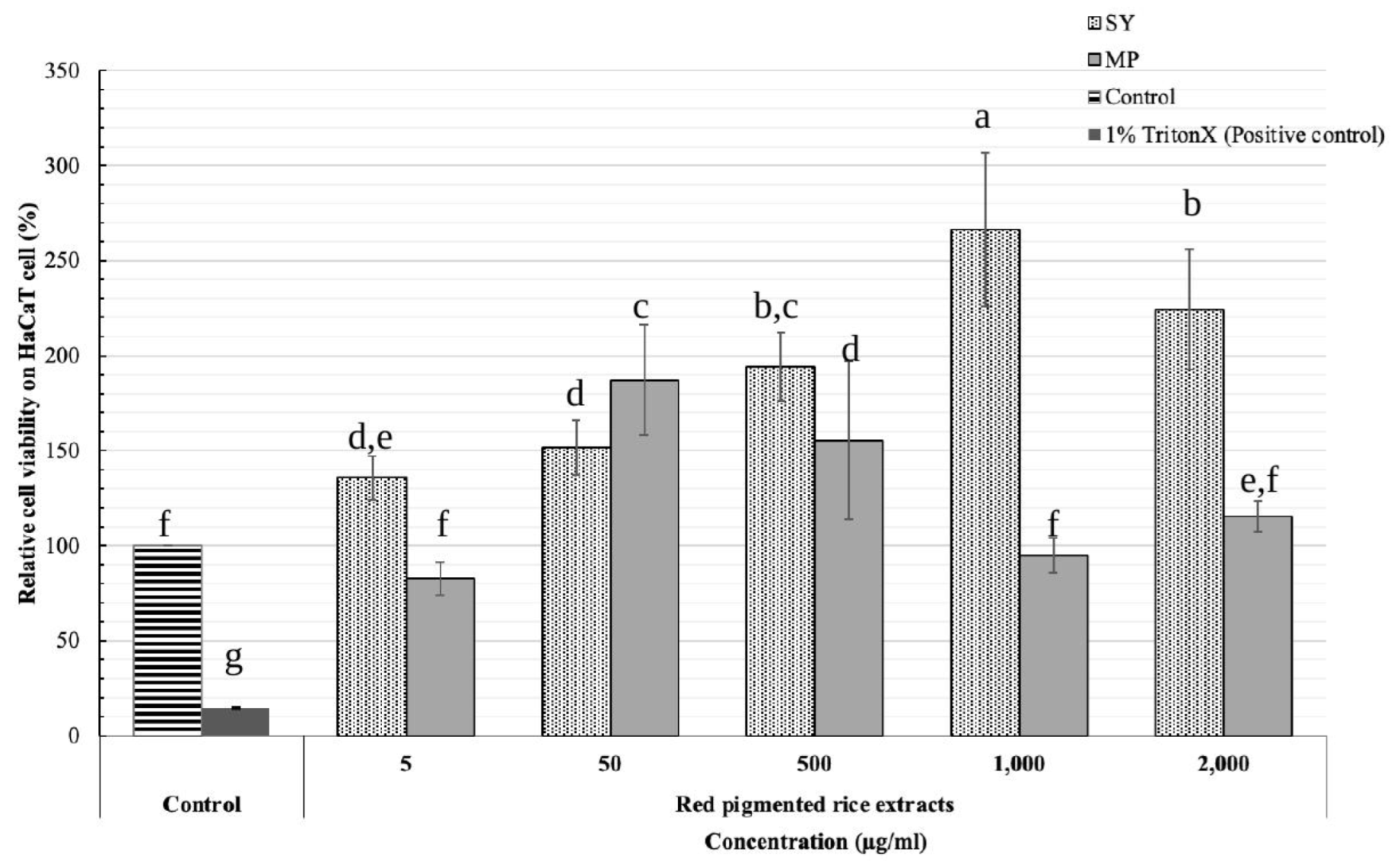
The safety of red rice extract for cosmetic application was determined on a Human immortalised non-tumorigenic keratinocyte cell line (HaCaT) (CLS Cell Lines Service, Eppelheim, Germany) using the MTT assay. SY and MP red pigmented rice extracts with high 5α-reductase inhibitory activity were cytotoxically tested at concentrations of 5, 50, 500, 1000 and 2000 µg/mL. An untreated sample (control) and 1% Triton X (positive control) (Sigma, USA) were used as standard reference. Each sample was determined in triplicate. Results were expressed as relative cell viability (%).
2.7. In Vitro Hair Growth-Promoting Activity on Human Hair Germinal Matrix Cells (HHGMC)
2.7.1. Cell Culture
Human hair germinal matrix cells (HHGMC) (ScienceCell Research Laboratories, Carlsbad, CA, USA) were used for the determination of hair growth-promoting activity. HHGMC cells were cultured in mesenchymal stem cell medium with 5% fetal bovine serum, 5% mesenchymal supplement, and 1% penicillin-streptomycin (Gibco, Gaithersburg, MD, USA) and were maintained at 37 °C in a humidified 5% CO2 incubator (Thermo Fisher Scientific, Forma II, Waltham, MA, USA).
2.7.2. In Vitro Hair Growth-Promoting Activity on HHGMC Cell Culture
For measuring the effects of rice extract on HHGMC proliferation, cells were seeded into a 96-well plate at 2 × 103 cells/well, treated with rice extracts (1 mg/mL of each SY, MP, BG, HN extracts), and incubated for 72 h at 37 °C in a CO2 incubator. Catechin (Sigma, USA), Epicatechin (Sigma, USA), Oryzanol (Sigma, USA), and Procyanidin B2 (Sigma, USA) were used as reference standards; 1% Finasteride and 1% Minoxidil (Sigma, USA) were used as positive controls. The percentage of living cells was determined using the MTT assay and the Trypan blue staining assay.
For the MTT proliferation methods, the treated and control culture cells were mixed with 100 µL of MTT reagent (0.5 mg/mL) and incubated for 4 h. Formazan was solubilized by adding 200 µL DMSO for 30 min. The medium was a centrifuge and absorbencies were recorded at 570 nm using a microplate reader. The relative cell proliferation (%) compared to control cells was calculated for the MTT assay using the following equation:
For the Trypan blue staining assay, culture cells were trypsinized, stained with Trypan Blue (Sigma, USA) at cell:staining dye ratio of 1:1 (
v/
v) and counted using a hemocytometer (Marienfeld, Germany). Results were expressed as the relative percentage of the total cell number using the following equation:
The ratio of living cells to dead cells from the total cell number of each sample were determined.
2.8. In Vivo Preliminary Rat Skin Irritation
The acute dermal irritation test was modified from methods of Reungpatthanaphong et al. [
16]. Two adult male Spague-Dawley rats were used after acclimatization in the laboratory. One day before experimentation, an area of dorsal-lumbar skin approximately 6 × 6 cm
2 in area was gently shaved with an electrical shaver and cleaned with normal saline. A single application closed patch test was preliminary performed on the rat skin for 24 h. Finn chambers (8 mM) (Smartpractice, McDowell, WV, USA) were used for observation of skin irritation (20 µL) on the shaved rats’ skin. The skin irritation severity was graded 0–4 according to the Draize dermal irritation scoring system (erythema and edema formation). An erythema/edema score ≤1 was interpreted as no irritation. For the skin irritation test, the samples were: (1) Untreated skin, (2) Normal saline as the negative control, (3) 1 mg/mL of each rice extracts (SY, MP, BG, and MP extracts).
2.9. In Vivo Hair Growth-Promoting Activity in C3H/HeMlac Mice
SY and MP red pigmented rice extracts that showed high 5α-reductase inhibitory activity with no cytotoxicity were tested on adult male C3H/HeMlac mice (
Mus musculus) with modified methods [
6,
7,
17]. An area of 8 cm
2 from the mouse (9 weeks of age) dorsal area was gently shaved using an electrical shaver. A total of 0.5 mL of each rice extract (concentration 1 mg/mL) was applied on the animal’s shaved skin one time per day for 28 days. Each sample was determined in duplicate. Changes in hair growth of each mouse were recorded and photographed. Areas of extensive hair growth with visible hair (re-growth light gray hair to mature dark brown hair) were determined. DI water (Deionized water) and 2% minoxidil (as a topical anti-hair loss drug) were used as a reference control. Moreover, the histological change in the dorsal skin at day 28 was determined with paraffin sections [
18]. Skin tissue was preserved in neutral buffer formalin (NBF) (Sigma, USA) and embedded in paraffin (Thermo Scientific, Histostar, USA). The paraffin was sectioned at 6 µm thickness (Leica Biosystem, RM2123, Germany) and stained with hematoxylin (Sigma, USA) and eosin (Sigma, USA) dyes. Skin morphology was determined with a microscope (Nikon, Eclipse TS100F, Tokyo, Japan).
2.10. Statistical Analysis
Data were reported as the mean ± SD. The difference between groups in each parameter was statistically analyzed by an analysis of variance (ANOVA) test with Duncan’s multiple range test (IBM SPSS version 21.0). The level of significance was at p < 0.05.
4. Discussion
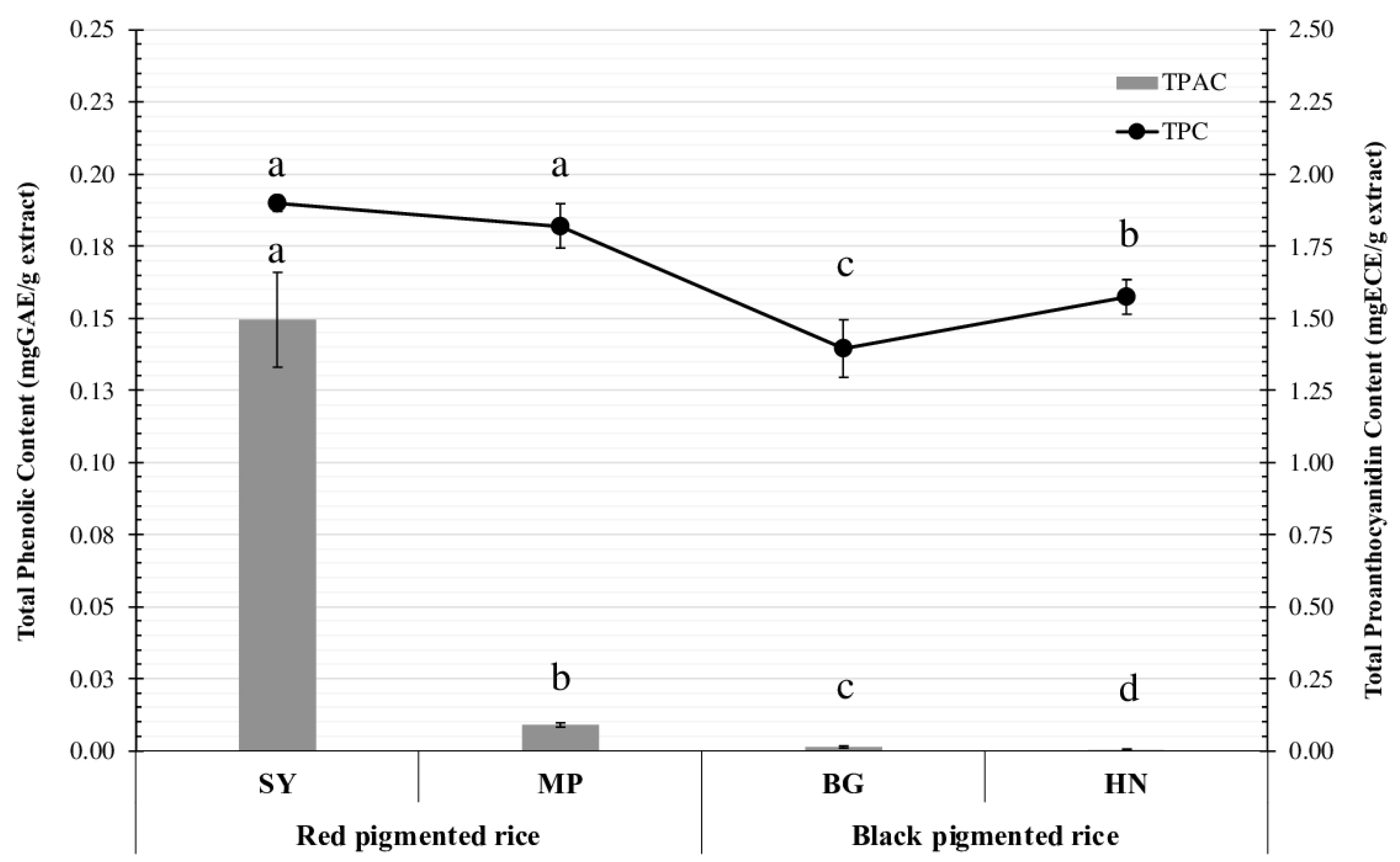
Ethanolic extracts from red pigmented rice (especially Sangyod rice) exhibited significantly higher bioactive contents including
TPAC than the extracts of black pericarp rice (BG and HN) (
p < 0.05). The
TPAC was reported as being important in cosmetics for hair growth treatment [
6,
7]. Proanthocyanins are oligomers and polymers of flavan-3-ols that make up the bulk of the phenolic compounds found in red rice, being responsible for the red pigmentation of the pericarp [
14]. Proanthocyanidins have not been detected in white rice, while some black or purple pericarp rice varieties have been reported to contain them [
19,
20,
21]. Moreover, proanthocyanidins (especially procyanidin B2) in some plants, e.g., grape seed and apple, are reported to be natural hair growth-inducing agents that can promote proliferation of in vitro mouse hair follicle cells and in vivo hair-cycle-converting activity in C3H mice [
6,
7], as well as human hair growth, as supported by a clinical trial that studied the topical application of proanthocyanidins [
8,
9].
There are several biological mechanisms for hair growth promotion and this study focused on in vitro 5αR inhibitory activity, in vitro cell proliferation of human hair germinal matrix (HHGMC) cells, and in vivo hair growth in C3H/HeMlac mice. Red SY pericarp extract had high potential for hair growth-promoting activity according to the results of these in vitro and in vivo tests.
Comparing the 5αR inhibitory activity of red rice and traditional Thai plants, Sang-Yod extract had lower 5αR inhibitory activity than the flower of
Carthamus tinctorius L. (24.3 ± 1.6 mgFEA/g crude extract), however, the enzyme inhibitory activity of SY red pericarp was more potent than that of various traditional Thai plants including
Clitoria ternatea L. (15.4 ± 0.7 mgFEA/g crude extract),
Citrus hystrix DC. (13.7 ± 0.8 mgFEA/g crude extract),
Ipomoea aquatica Forssk. (13.2 ± 0.4 mgFEA/g crude extract),
Cassia siamea Lam. (12.9 ± 1.1 mgFEA/g crude extract), and
Rhinacanthus nasutus (L.) Kurz. (10.7 ± 1.0 mgFEA/g crude extract) [
5]. Furthermore, baldness, especially in men, occurs particularly under the influence of the androgenic hormones, which are important regulators of hair growth. Hair loss occurs more often in people who have a more active 5α-reductase enzyme in their hair follicles: this enzyme changes testosterone to the stronger androgen-signaling dihydrotestosterone (DHT) [
2]. Inhibition of 5α-reductase is one of three possible anti-androgen mechanisms of action for herbal hair loss treatment [
4].
The hair follicle consists of the epithelial component (matrix and outer-root sheath) and dermal components (dermal papilla at the base of the hair follicle and connective tissue sheath). Dermal papilla cells play a role in the morphogenesis and regeneration of hair growth [
22]. Moreover, normal hair follicle growth depends on the activation of hair matrix cells. Anagen duration determines the hair shaft length and is dependent on the continued proliferation and differentiation of matrix cells in the matrix region of the hair follicle. Their proliferation is an important event for hair growth [
23,
24]. Previous studies showed that procyanidin B2 (an oligomer of proanthocyanidin) from apple juice as natural hair growth promoter can strongly promote the proliferation of mouse hair epithelial cells, with 300% more proliferation (at 30 µM) relative to control (=100%) for a 5-day culture, which is higher than the proliferation rates obtained with epicatechin (as a monomer of proanthocyanidin) and minoxidil [
6,
7]. Furthermore, procyanidins are reported to have no effect on the proliferation of mouse dermal papilla cells [
7]. However, there has been little research on the efficiency of natural extracts in terms of human matrix cell (germinative keratinocyte) proliferation for hair growth. Sang-Yod red rice extract with high contents of proanthocyanidins significantly increased the proliferation of human hair matrix cells (216.2% relative cell proliferation) to a greater extent than did procyanidin B2 (159.7%), epicatechin (80.2%) and minoxidil (93.9%), although the conditions of this study were different from previous studies in terms of the cell origins (human) and incubation time (3-day culture) [
6,
7]. These results demonstrated the potential feasibility of SY red pericarp extract as a candidate for the treatment of hair loss.
These results from in vivo C3H/HeMlac mice testing demonstrated that red pigmented rice, especially SY rice extract, possesses a marked hair-growing activity that is able to induce anagen phase in vivo, according to previous data obtained for proanthocyanin and procyanidin oligomer from grape seed, apple, and barley extracts in the same animal strain [
6,
7,
11]. Previous studies on in vivo hair growth-promoting effects in another mice strain (C57BL/6 mice) showed that rice bran extracts from Korean cultivars (
Oryza sativa var.
japonica) extracted using supercritical carbon dioxide fluid can induce hair follicles to enter the anagen stage (score 4: hair growth 60–80%) [
1,
11]. The major components of these rice bran oils that act to promote hair growth are γ-oryzanol and linoleic acid [
1]. However, γ-oryzanol as a reference substance in this study had slightly low activity (<100%) in terms of human matrix cell proliferation, while procyanidin B2 can induce hair matrix cell proliferation (>150%). Therefore, the functional substances for hair growth-promoting activity in the crude extract of red pigmented rice (
Oryza sativa), including Sang-Yod rice, might be proanthocyanidins, which gave higher activity of 5α-reductase inhibition, higher HaCaT and human matrix cell proliferation, and higher hair-cycle-converting activity from telogen to anagen phase in C3H mice, as in previous studies [
6,
7,
11,
14].


 ,
, 




{kind=link}
{kind=link}
{kind=link}