On the Inverse Correlation of Protein and Oil: Examining the Effects of Altered Central Carbon Metabolism on Seed Composition Using Soybean Fast Neutron Mutants

 , , and
, , and
Abstract
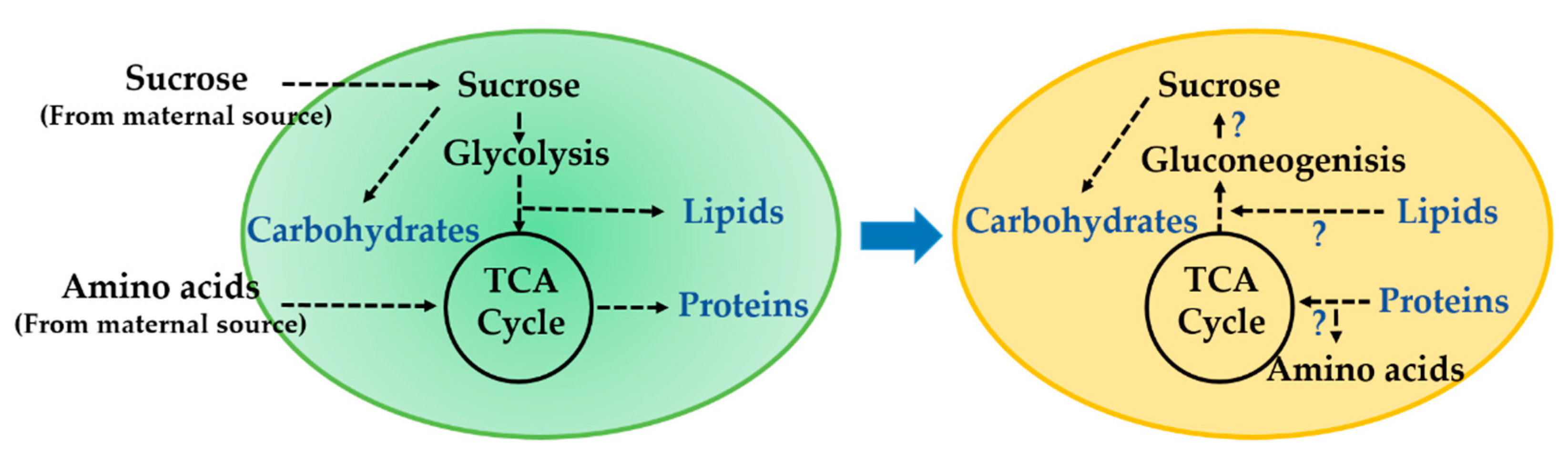
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Extraction of Lipids, Protein, and Soluble Sugars
2.3. FAME Analysis for the Quantification of Lipids
2.4. Quantification of Sugars using HILIC-MS/MS
2.5. Linear Modeling and Statistical Analyses
3. Results
3.1. Identification of FN Lines with Alterations in Central Carbon Metabolic Genes
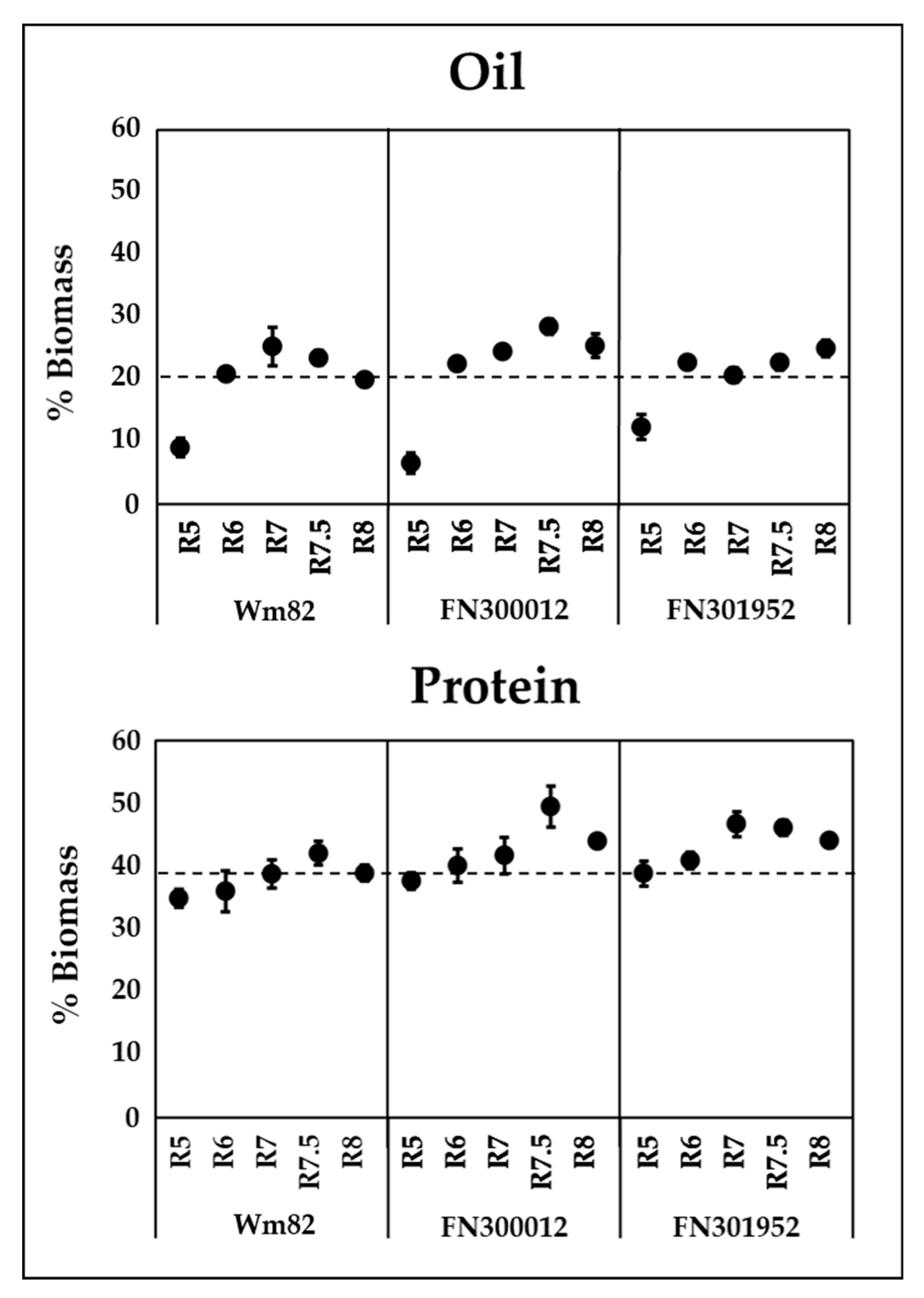
3.2. FN300012 and FN301952 Accumulated High Oil without a Decline in Their Total Protein Content
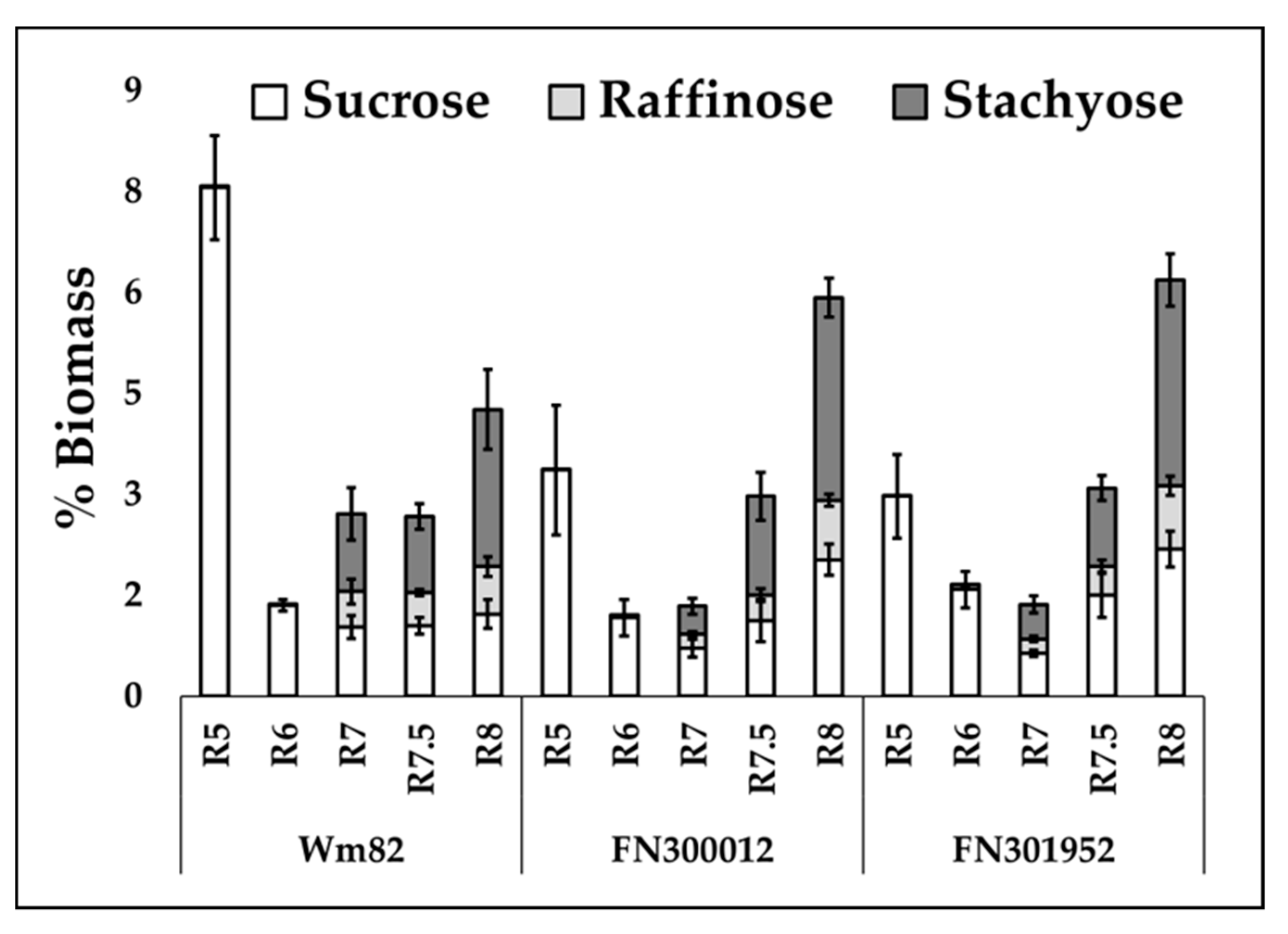
3.3. No Decrease in RFOs Was Observed in the Final Composition of Selected Lines
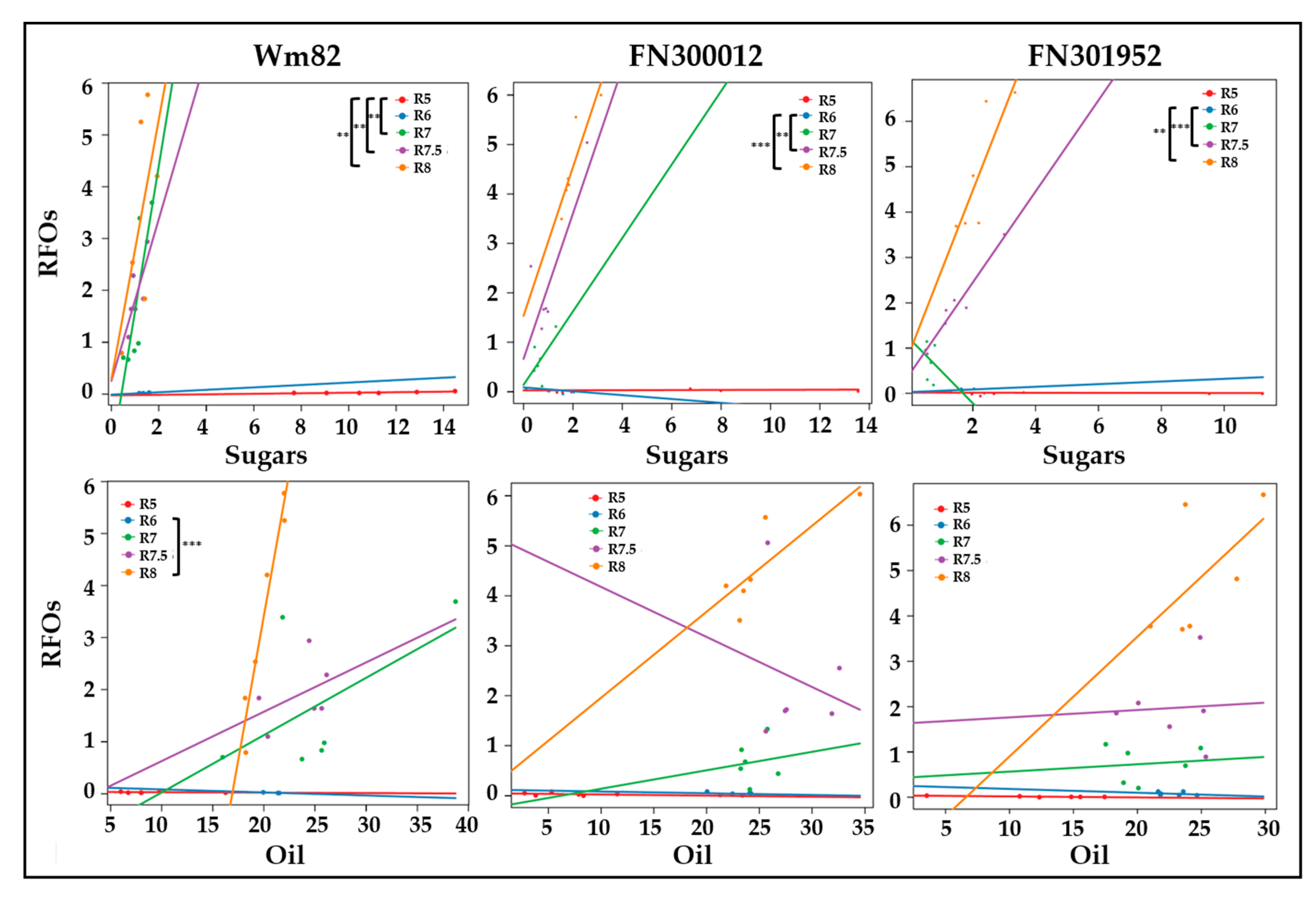
3.4. A Delayed Switch towards RFO Accumulation Leads to Extended Oil and Protein Accumulation
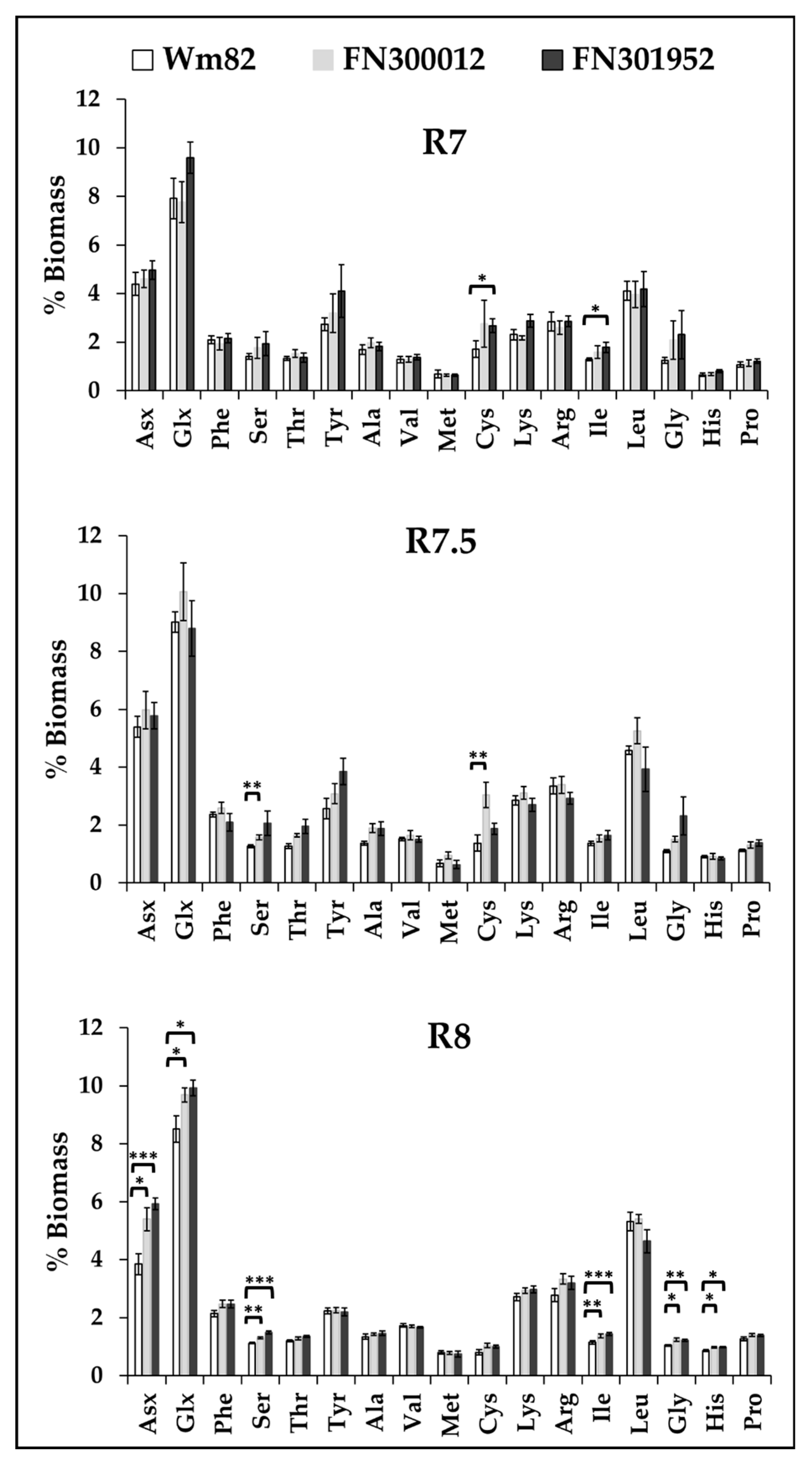
3.5. Cysteine Content That Contributes to Protein Quality Peaked Earlier in Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2016, 68, 827–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdier, J.; Lalanne, D.; Pelletier, S.; Torres-Jerez, I.; Righetti, K.; Bandyopadhyay, K.; Leprince, O.; Chatelain, E.; Vu, B.L.; Gouzy, J.; et al. A regulatory network-based approach dissects late maturation processes related to the acquisition of desiccation tolerance and longevity of medicago truncatula seeds. Plant Physiol. 2013, 163, 757–774. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Rani, A.; Goyal, L.; Dixit, A.K.; Manjaya, J.G.; Dev, J.; Swamy, M. Sucrose and raffinose family oligosaccharides (RFOs) in soybean seeds as influenced by genotype and growing location. J. Agric. Food Chem. 2010, 58, 5081–5085. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, S.J.; Hadley, H.H. Maternal effects on sugar content in soybean seeds. Crop Sci. 1978, 18, 581–584. [Google Scholar] [CrossRef]
- Dierking, E.C.; Bilyeu, K.D. Association of a soybean raffinose synthase gene with low raffinose and stachyose seed phenotype. Plant Genome 2008, 1, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Coon, C.N.; Leske, K.L.; Akavanichan, O.; Cheng, T.K. Effect of oligosaccharide-free soybean meal on true metabolizable energy and fiber digestion in adult roosters1. Poult. Sci. 1990, 69, 787–793. [Google Scholar] [CrossRef]
- Koster, K.L.; Leopold, A.C. Sugars and desiccation tolerance in seeds. Plant Physiol. 1988, 88, 829–832. [Google Scholar] [CrossRef] [Green Version]
- Horbowicz, M.; Obendorf, R.L. Seed desiccation tolerance and storability: Dependence on flatulence-producing oligosaccharides and cyclitols—Review and survey. Seed Sci. Res. 2008, 4, 385–405. [Google Scholar] [CrossRef]
- Peterbauer, T.; Richter, A. Biochemistry and physiology of raffinose family oligosaccharides and galactosyl cyclitols in seeds. Seed Sci. Res. 2001, 11, 185–197. [Google Scholar]
- Blöchl, A.; Peterbauer, T.; Richter, A. Inhibition of raffinose oligosaccharide breakdown delays germination of pea seeds. J. Plant Physiol. 2007, 164, 1093–1096. [Google Scholar] [CrossRef]
- Gangl, R.; Tenhaken, R. Raffinose family oligosaccharides act as galactose stores in seeds and are required for rapid germination of arabidopsis in the dark. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hincha, D.K.; Zuther, E.; Heyer, A.G. The preservation of liposomes by raffinose family oligosaccharides during drying is mediated by effects on fusion and lipid phase transitions. Biochim. Biophys. Acta-Biomembr. 2003, 1612, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Neus, J.D.; Fehr, W.R.; Schnebly, S.R. Agronomic and seed characteristics of soybean with reduced raffinose and stachyose. Crop Sci. 2005, 45, 589–592. [Google Scholar] [CrossRef]
- Dierking, E.C.; Bilyeu, K.D. Raffinose and stachyose metabolism are not required for efficient soybean seed germination. J. Plant. Physiol. 2009, 166, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Valentine, M.F.; De Tar, J.R.; Mookkan, M.; Firman, J.D.; Zhang, Z.J. Silencing of soybean raffinose synthase gene reduced raffinose family oligosaccharides and increased true metabolizable energy of poultry feed. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, T.E.; Cahoon, E.B. Soybean Oil: Genetic Approaches for modification of functionality and total content. Plant Physiol. 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, J.; Patil, G.B.; Sonah, H.; Deshmukh, R.K.; Vuong, T.D.; Valliyodan, B.; Nguyen, H.T. Expanding omics resources for improvement of soybean seed composition traits. Front. Plant. Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Patil, G.; Mian, R.; Vuong, T.; Pantalone, V.; Song, Q.; Chen, P.; Shannon, G.J.; Carter, T.C.; Nguyen, H.T. Molecular mapping and genomics of soybean seed protein: A review and perspective for the future. Theor. Appl. Genet. 2017, 130, 1975–1991. [Google Scholar] [CrossRef] [Green Version]
- Rincker, K.; Nelson, R.; Specht, J.; Sleper, D.; Cary, T.; Cianzio, S.R.; Casteel, S.; Conley, S.; Chen, P.; Davis, V.; et al. Genetic improvement of U.S. soybean in maturity groups II, III, and IV. Crop Sci. 2014, 54, 1419–1432. [Google Scholar] [CrossRef]
- Bandillo, N.; Jarquin, D.; Song, Q.; Nelson, R.; Cregan, P.; Specht, J.; Lorenz, A. A population structure and genome-wide association analysis on the USDA soybean germplasm collection. Plant Genome 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Brzostowski, L.F.; Diers, B.W. Agronomic evaluation of a high protein allele from PI407788A on chromosome 15 across two soybean backgrounds. Crop Sci. 2017, 57, 2972–2978. [Google Scholar] [CrossRef]
- Allen, D.K.; Young, J.D. Carbon and nitrogen provisions alter the metabolic flux in developing soybean embryos. Plant Physiol. 2013, 161, 1458–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, D.K.; Ohlrogge, J.B.; Shachar-Hill, Y. The role of light in soybean seed filling metabolism. Plant J. 2009, 58, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Truong, Q.; Koch, K.; Yoon, J.M.; Everard, J.D.; Shanks, J.V. Influence of carbon to nitrogen ratios on soybean somatic embryo (cv. Jack) growth and composition. J. Exp. Bot. 2013, 64, 2985–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotundo, J.L.; Borrás, L.; Westgate, M.E.; Orf, J.H. Relationship between assimilate supply per seed during seed filling and soybean seed composition. Field Crop. Res. 2009, 112, 90–96. [Google Scholar] [CrossRef]
- Fait, A.; Angelovici, R.; Less, H.; Ohad, I.; Urbanczk-Wochniak, E.; Fernie, A.R.; Galili, G. Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 2006, 142, 839–854. [Google Scholar] [CrossRef] [Green Version]
- Galili, G.; Avin-Wittenberg, T.; Angelovici, R.; Fernie, A.R. The role of photosynthesis and amino acid metabolism in the energy status during seed development. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef]
- Collakova, E.; Aghamirzaie, D.; Fang, Y.; Klumas, C.; Tabataba, F.; Kakumanu, A.; Myers, E.; Heath, L.S.; Grene, R. Metabolic and transcriptional reprogramming in developing soybean (Glycine max) embryos. Metab. 2013, 3, 347–372. [Google Scholar] [CrossRef]
- Baud, S.; Boutin, J.-P.; Miquel, M.; Lepiniec, L.; Rochat, C. An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 2002, 40, 151–160. [Google Scholar] [CrossRef]
- Chia, T.Y.P.; Pike, M.J.; Rawsthorne, S. Storage oil breakdown during embryo development of Brassica napus (L.). J. Exp. Bot. 2005, 56, 1285–1296. [Google Scholar] [CrossRef]
- Angelovici, R.; Galili, G.; Fernie, A.R.; Fait, A. Seed desiccation: A bridge between maturation and germination. Trends Plant Sci. 2010, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Pipolo, A.E.; Sinclair, T.R.; Camara, G.M.S. Protein and oil concentration of soybean seed cultured in vitro using nutrient solutions of differing glutamine concentration. Ann. Appl. Biol. 2004, 144, 223–227. [Google Scholar] [CrossRef]
- Campbell, B.W.; Hofstad, A.N.; Sreekanta, S.; Fu, F.; Kono, T.J.Y.; O’Rourke, J.A.; Vance, C.P.; Muehlbauer, G.J.; Stupar, R.M. Fast neutron-induced structural rearrangements at a soybean NAP1 locus result in gnarled trichomes. Theor. Appl. Genet. 2016, 129, 1725–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stacey, M.G.; Cahoon, R.E.; Nguyen, H.T.; Cui, Y.; Sato, S.; Nguyen, C.T.; Phoka, N.; Clark, K.M.; Liang, Y.; Forrester, J.; et al. Identification of homogentisate dioxygenase as a target for vitamin E biofortification in oilseeds. Plant Physiol. 2016, 172, 1506–1518. [Google Scholar] [CrossRef] [Green Version]
- Dobbels, A.A.; Michno, J.-M.; Campbell, B.W.; Virdi, K.S.; Stec, A.O.; Muehlbauer, G.J.; Naeve, S.L.; Stupar, R.M. An induced chromosomal translocation in soybean disrupts a KASI ortholog and is associated with a high-sucrose and low-oil seed phenotype. G3 Genes Genomes Genet. 2017, 7, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Men, A.E.; Laniya, T.S.; Searle, I.R.; Iturbe-Ormaetxe, I.; Gresshoff, I.; Jiang, Q.; Carroll, B.J.; Gresshoff, P.M. Fast neutron mutagenesis of soybean (Glycine soja L.) produces a supernodulating mutant containing a large deletion in linkage group H. Genome Lett. 2002, 1, 147–155. [Google Scholar] [CrossRef]
- Bolon, Y.-T.; Haun, W.J.; Xu, W.W.; Grant, D.; Stacey, M.G.; Nelson, R.T.; Gerhardt, D.J.; Jeddeloh, J.A.; Stacey, G.; Muehlbauer, G.J.; et al. Phenotypic and genomic analyses of a fast neutron mutant population resource in soybean. Plant Physiol. 2011, 156, 240–253. [Google Scholar] [CrossRef] [Green Version]
- Bolon, Y.-T.; Stec, A.O.; Michno, J.-M.; Roessler, J.; Bhaskar, P.B.; Ries, L.; Dobbels, A.A.; Campbell, B.W.; Young, N.P.; Anderson, J.E.; et al. Genome resilience and prevalence of segmental duplications following fast neutron irradiation of soybean. Genetics 2014, 198, 967–981. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.J.; Kim, M.Y.; Kang, Y.J.; Shim, S.; Stacey, M.G.; Stacey, G.; Lee, S.-H. Genome-wide analysis of mutations in a dwarf soybean mutant induced by fast neutron bombardment. Euphytica 2015, 203, 399–408. [Google Scholar] [CrossRef]
- Islam, N.; Stupar, R.M.; Qijian, S.; Luthria, D.L.; Garrett, W.; Stec, A.O.; Roessler, J.; Natarajan, S.S. Genomic changes and biochemical alterations of seed protein and oil content in a subset of fast neutron induced soybean mutants. BMC Plant Biol. 2019, 19, 420. [Google Scholar] [CrossRef]
- Prenger, E.M.; Ostezan, A.; Mian, M.A.R.; Stupar, R.M.; Glenn, T.; Li, Z. Identification and characterization of a fast-neutron-induced mutant with elevated seed protein content in soybean. Theor. Appl. Genet. 2019, 132, 2965–2983. [Google Scholar] [CrossRef] [PubMed]
- Naeve, S.L. Soybean Growth Stages. Available online: https://extension.umn.edu/growing-soybean/soybean-growth-stages (accessed on 31 October 2019).
- Licht, M. Soybean Growth and Development. Available online: https://store.extension.iastate.edu/product/6451 (accessed on 31 October 2019).
- Ma, F.; Jazmin, L.J.; Young, J.D.; Allen, D.K. Isotopically nonstationary metabolic flux analysis (INST-MFA) of photosynthesis and photorespiration in plants. In Photorespiration: Methods and Protocols; Fernie, A.R., Bauwe, H., Weber, A.P.M., Eds.; Springer: New York, NY, USA, 2017; pp. 167–194. [Google Scholar]
- Kambhampati, S.; Li, J.; Evans, B.S.; Allen, D.K. Accurate and efficient amino acid analysis for protein quantification using hydrophilic interaction chromatography coupled tandem mass spectrometry. Plant Methods 2019, 15, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, A.H.; Zhang, X.; Jastrebski, S.; Lamont, S.J.; Singh, A.; Schmidt, C.J. Identifying mechanisms of regulation to model carbon flux during heat stress and generate testable hypotheses. PLoS ONE 2018, 13, e0205824. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B. Engineering soybean for enhanced sulfur amino acid content. Crop Sci. 2005, 45, 454–461. [Google Scholar] [CrossRef]
- Krishnan, H.B.; Jez, J.M. Review: The promise and limits for enhancing sulfur-containing amino acid content of soybean seed. Plant Sci. 2018, 272, 14–21. [Google Scholar] [CrossRef]
- Li, L.; Hur, M.; Lee, J.-Y.; Zhou, W.; Song, Z.; Ransom, N.; Demirkale, C.Y.; Nettleton, D.; Westgate, M.; Arendsee, Z.; et al. A systems biology approach toward understanding seed composition in soybean. BMC Genom. 2015, 16, S9. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-S.; Jez, J.M.; Krishnan, H.B. Effects of proteome rebalancing and sulfur nutrition on the accumulation of methionine rich δ-zein in transgenic soybeans. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.A.; Barbazuk, W.B.; Sandford, M.; May, G.; Song, Z.; Zhou, W.; Nikolau, B.J.; Herman, E.M. Silencing of soybean seed storage proteins results in a rebalanced protein composition preserving seed protein content without major collateral changes in the metabolome and transcriptome. Plant Physiol. 2011, 156, 330–345. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zheng, W.; Zhu, Y.; Ye, H.; Tang, B.; Arendsee, Z.W.; Jones, D.; Li, R.; Ortiz, D.; Zhao, X.; et al. QQS orphan gene regulates carbon and nitrogen partitioning across species via NF-YC interactions. Proc. Natl. Acad. Sci. USA 2015, 112, 14734. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wurtele, E.S. The QQS orphan gene of Arabidopsis modulates carbon and nitrogen allocation in soybean. Plant Biotechnol. J. 2015, 13, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.F. Seed composition. In Soybeans: Improvement, Production, and Uses; Boerma, H.R., Specht, J.E., Eds.; American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America: Madison, WI, USA, 2004; pp. 621–677. [Google Scholar]
- Rotundo, J.L. Linking assimilate supply and seed developmental processes that determine soybean seed composition. Eur. J. Agron. 2011, 35, 184–191. [Google Scholar] [CrossRef]
- Grant, D.; Nelson, R.T.; Cannon, S.B.; Shoemaker, R.C. SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Res. 2009, 38, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Litterer, L.A.; Plaisance, K.L.; Schnurr, J.A.; Storey, K.K.; Jung, H.-J.G.; Gronwald, J.W.; Somers, D.A. Biosynthesis of UDP-glucuronic acid in developing soybean embryos: Possible role of UDP-sugar pyrophosphorylase. Physiol. Plant. 2006, 128, 200–211. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef] [Green Version]
- van Erp, H.; Kelly, A.A.; Menard, G.; Eastmond, P.J. Multigene engineering of triacylglycerol metabolism boosts seed oil content in arabidopsis. Plant Physiol. 2014, 165, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.A.; Shaw, E.; Powers, S.J.; Kurup, S.; Eastmond, P.J. Suppression of the SUGAR-DEPENDENT1 triacylglycerol lipase family during seed development enhances oil yield in oilseed rape (Brassica napus L.). Plant Biotechnol. J. 2013, 11, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Yang, S.W.; Mao, H.-Z.; Veena, S.P.; Yin, J.-L.; Chua, N.-H. Gene silencing of sugar-dependent 1 (JcSDP1), encoding a patatin-domain triacylglycerol lipase, enhances seed oil accumulation in Jatropha curcas. Biotechnol. Biofuels 2014, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Kanai, M.; Yamada, T.; Hayashi, M.; Mano, S.; Nishimura, M. Soybean (Glycine max L.) triacylglycerol lipase GmSDP1 regulates the quality and quantity of seed oil. Sci. Rep. 2019, 9, 8924. [Google Scholar] [CrossRef]
- Müntz, K. Proteases and proteolytic cleavage of storage proteins in developing and germinating dicotyledonous seeds. J. Exp. Bot. 1996, 47, 605–622. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FN Descriptor | Descriptors Used Here | 50-Seed Weight (g) | Genes of Interest within FN Deletion or Phenotype Used as Selection Criteria |
|---|---|---|---|
| Williams 82 | WT-Wm 82 | 10.05 | Wild type |
| FN300660 | FN300660 | 11.87 | α/β hydrolase, Glyma14g09181 |
| FN300088 | FN300088 | 10.49 | α/β hydrolase, Glyma17g35990 |
| FN300068.01.01.M4 | FN300068 | 13.62 | F1,6BPase, Glyma16g28310 |
| FN300012.02.BC1F3 | FN300012 | 11.15 | UDP-Gluc-6DH, Glyma01g06970 |
| FN300026.02.01.M4 | FN300026 | 11.57 | PEPCK, Glyma04g09510 |
| FN300108 | FN300108 | 7.66 | ACX2, Glyma06g43840 |
| FN301952.01.01.M4 | FN301952 | 13.33 | Enolase, Glyma03g34830 |
| FN301653.01.01.M4 | FN301653 | 12.11 | F1,6BPase, Glyma18g41940 |
| M92-220 | WT-M92-220 | 13.65 | Wild type |
| FN0171815 | FN0171815 | 14.63 | high oil |
| FN0110332.03.M3 | FN0110332 | 11.98 | Aldolase, Glyma12g24190 |
| FN0170714.03.01.02.M5 | FN0170714 | 16.3 | F1,6BPase Glyma18g41940 |
| P1082Daar541BMN15 1 | P1082D | 10.76 | high protein |
| FN0175143 | FN0175143 | 13.8 | high oil |
| FN0171855.08.01.01.10.M6 | FN0171855 | 13.32 | PEPCK Glyma04g09510 |
| 5R16CO1Ddar117MN15 1 | 5R16CO1D | 12.92 | high oil |
| FN0170904.01.M3 | FN0170904 | 13.93 | F1,6BPase Glyma16g28310 |
| FN0171734 | FN0171734 | 9.93 | SUS1/4 Glyma09g08550 |
| PO559Caar541MN15 1 | PO559C | 12.75 | high protein + oil |
| FN0141075.03.04.10.M5 | FN0141075 | 12.29 | ACX1 Glyma01g41600 |
| 1R37C45cbdaar254MN15 1 | 1R37C45 | 13.28 | high protein + oil |
| FN0171466 | FN0171466 | 14.23 | high oil |
| FN0173708 | FN0173708 | 13.32 | high oil |
| FN0175116 | FN0175116 | 13.08 | high oil |
| FN0111996 | FN0111996 | 13.53 | PXA1 Glyma11g38160 |
| FN0173054.M2 | FN0173054 | 14.18 | PGM Glyma08g04890 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kambhampati, S.; Aznar-Moreno, J.A.; Hostetler, C.; Caso, T.; Bailey, S.R.; Hubbard, A.H.; Durrett, T.P.; Allen, D.K. On the Inverse Correlation of Protein and Oil: Examining the Effects of Altered Central Carbon Metabolism on Seed Composition Using Soybean Fast Neutron Mutants. Metabolites 2020, 10, 18. https://doi.org/10.3390/metabo10010018
Kambhampati S, Aznar-Moreno JA, Hostetler C, Caso T, Bailey SR, Hubbard AH, Durrett TP, Allen DK. On the Inverse Correlation of Protein and Oil: Examining the Effects of Altered Central Carbon Metabolism on Seed Composition Using Soybean Fast Neutron Mutants. Metabolites. 2020; 10(1):18. https://doi.org/10.3390/metabo10010018
Chicago/Turabian StyleKambhampati, Shrikaar, Jose A. Aznar-Moreno, Cooper Hostetler, Tara Caso, Sally R. Bailey, Allen H. Hubbard, Timothy P. Durrett, and Doug K. Allen. 2020. "On the Inverse Correlation of Protein and Oil: Examining the Effects of Altered Central Carbon Metabolism on Seed Composition Using Soybean Fast Neutron Mutants" Metabolites 10, no. 1: 18. https://doi.org/10.3390/metabo10010018
APA StyleKambhampati, S., Aznar-Moreno, J. A., Hostetler, C., Caso, T., Bailey, S. R., Hubbard, A. H., Durrett, T. P., & Allen, D. K. (2020). On the Inverse Correlation of Protein and Oil: Examining the Effects of Altered Central Carbon Metabolism on Seed Composition Using Soybean Fast Neutron Mutants. Metabolites, 10(1), 18. https://doi.org/10.3390/metabo10010018