Vitamin D Metabolism and Profiling in Veterinary Species


Abstract
:1. Introduction
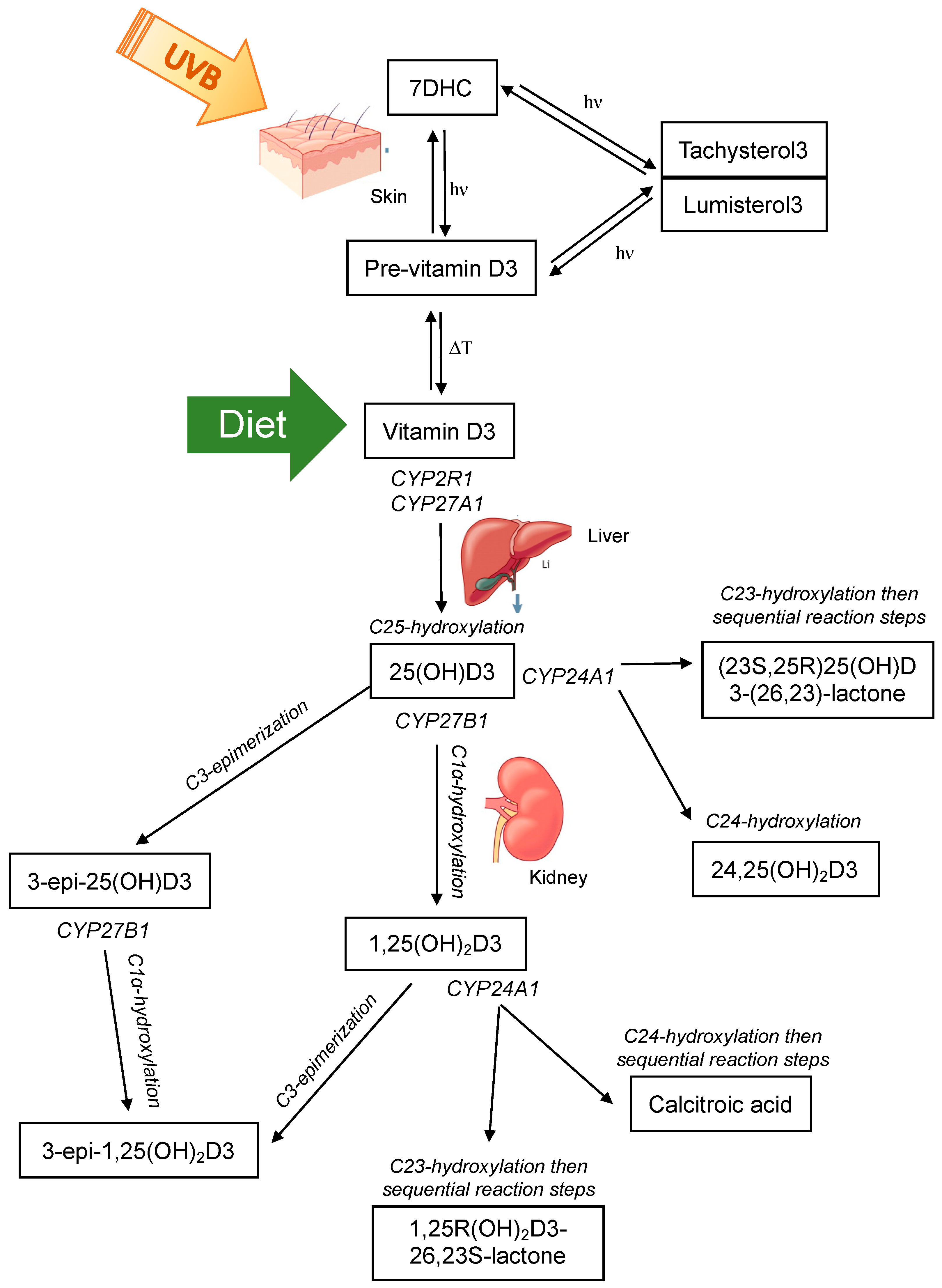
2. Fundamentals of Vitamin D Metabolism
3. Vitamin D Metabolism in Veterinary Species
3.1. Comparative Differences in Vitamin D Metabolism
3.1.1. Dogs and Cats
3.1.2. Horses
3.1.3. Sheep and Goats
3.1.4. Cattle
3.1.5. Pigs
3.1.6. Poultry
3.1.7. Llamas and Alpacas
3.1.8. Nonhuman Primates
3.2. Profiling Vitamin D beyond 25-Hydoxyvitamin-D in Veterinary Species
3.2.1. Measuring 1α,25-Dihydroxyvitamin-D
3.2.2. 24,25-Dihydroxyvitamin D and Vitamin D Metabolite Ratios
3.2.3. C3-Epimers
3.2.4. Free Vitamin D and the Vitamin D Binding Protein
4. Analytical Challenges in Profiling Vitamin D Metabolites in Veterinary Species
4.1. Analytical Challenges of Vitamin D Analysis
4.2. Standardization of Vitamin D Analysis
4.3. Challenges in Veterinary Vitamin D Analysis
4.3.1. Sample Acquisition and Preparation
4.3.2. Assay Modification and Validation for Use in Veterinary Species
4.4. Analysis of Vitamin D from Non-Invasive Biological Matrices
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-sommeregger, S.; Köstenberger, M.; Tmava, A.; Gennaro, B.; Stefan, M.; Oliver, P. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Fraser, W.D.; Tang, J.C.Y.; Dutton, J.J.; Schoenmakers, I. Vitamin D Measurement, the Debates Continue, New Analytes Have Emerged, Developments Have Variable Outcomes. Calcif. Tissue Int. 2020, 106, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, I.R.; Bolland, M.J. Controversies in medicine: The role of calcium and vitamin D supplements in adults. Med. J. Aust. 2019, 211, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Bolland, M.J.; Grey, A.; Avenell, A. Effects of vitamin D supplementation on musculoskeletal health: A systematic review, meta-analysis, and trial sequential analysis. Lancet Diabetes Endocrinol. 2018, 6, 847–858. [Google Scholar] [CrossRef] [Green Version]
- Scientific Advisory Committee on Nutrition. Vitamin D and Health; Public Health England: London, UK, 2016.
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 Report on Dietary Reference Intakes for Calcium and Vitamin D from the Institute of Medicine: What Clinicians Need to Know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- Holick, M.F.; Binkley, N.C.; Bischoff-ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, C. The vitamin D metabolome: An update on analysis and function. Cell Biochem. Funct. 2019, 37, 408–423. [Google Scholar] [CrossRef]
- Tuckey, R.C.; Tang, E.K.Y.; Maresse, S.R.; Delaney, D.S. Catalytic properties of 25-hydroxyvitamin D3 3-epimerase in rat and human liver microsomes. Arch. Biochem. Biophys. 2019, 666, 16–21. [Google Scholar] [CrossRef]
- Zafalon, R.V.A.; Risolia, L.W.; Pedrinelli, V.; Vendramini, T.H.A.; Rodrigues, R.B.A.; Amaral, A.R.; Kogika, M.M.; Brunetto, M.A. Vitamin D metabolism in dogs and cats and its relation to diseases not associated with bone metabolism. J. Anim. Physiol. Anim. Nutr. (Berl.) 2019, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Parker, V.J.; Rudinsky, A.J.; Chew, D.J. Vitamin D metabolism in canine and feline medicine. J. Am. Vet. Med. Assoc. 2017, 250, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, K.E.; Thompson, K.G. Vitamin D Metabolism and Rickets in Domestic Animals: A Review. Vet. Pathol. 2011, 48, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Mellanby, R.J. Beyond the skeleton: The role of vitamin D in companion animal health. J. Small Anim. Pract. 2016, 57, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Corbee, R.J.; Vaandrager, A.B.; Kik, M.J.; Molenaar, M.R.; Hazewinkel, H.A.W. Cutaneous vitamin D synthesis in carnivorous species. Eur. Soc. Vet. Comp. Nutr. 2008, 96, 1. [Google Scholar]
- Morris, J.G. Ineffective vitamin D synthesis in cats is reversed by an inhibitor of 7-dehydrocholestrol-delta7-reductase. J. Nutr. 1999, 129, 903–908. [Google Scholar] [CrossRef] [Green Version]
- How, K.L.; Hazewinkel, H.A.W.; Mol, J.A. Photosynthesis of vitamin d in the skin of dogs, cats, and rats. Vet. Q. 1995, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- How, K.L.; Hazewinkel, H.A.W.; Mol, J.A. Dietary vitamin D dependence of cat and dog due to inadequate cutaneous synthesis of vitamin D. Gen. Comp. Endocrinol. 1994, 96, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Allenspach, K.; Rizzo, J.; Jergens, A.E.; Chang, Y.M. Hypovitaminosis D is associated with negative outcome in dogs with protein losing enteropathy: A retrospective study of 43 cases. BMC Vet. Res. 2017, 13, 96. [Google Scholar] [CrossRef] [Green Version]
- Titmarsh, H.; Gow, A.G.; Kilpatrick, S.; Sinclair, J.; Hill, T.; Milne, E.; Philbey, A.; Berry, J.; Handel, I.; Mellanby, R.J. Association of Vitamin D Status and Clinical Outcome in Dogs with a Chronic Enteropathy. J. Vet. Intern. Med. 2015, 29, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Titmarsh, H.F.; Lalor, S.M.; Tasker, S.; Barker, E.N.; Berry, J.; Gunn-More, D.; Mellanby, R.J. Vitamin D status in cats with feline immunodeficiency virus. Vet. Med. Sci. 2015, 1, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Titmarsh, H.F.; Gow, A.G.; Kilpatrick, S.; Cartwright, J.A.; Milne, E.M.; Philbey, A.W.; Berry, J.; Handel, I.; Mellanby, R.J. Low Vitamin D status is associated with systemic and gastrointestinal inflammation in dogs with a chronic enteropathy. PLoS ONE 2015, 10, e137377. [Google Scholar] [CrossRef] [Green Version]
- Gow, A.G.; Else, R.; Evans, H.; Berry, J.L.; Herrtage, M.E.; Mellanby, R.J. Hypovitaminosis D in dogs with inflammatory bowel disease and hypoalbuminaemia. J. Small Anim. Pract. 2011, 52, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Chacar, F.; Kogika, M.; Sanches, T.R.; Caragelasco, D.; Martorelli, C.; Rodrigues, C.; Capcha, J.M.C.; Chew, D.; Andrade, L. Urinary Tamm-Horsfall protein, albumin, vitamin D-binding protein, and retinol-binding protein as early biomarkers of chronic kidney disease in dogs. Physiol. Rep. 2017, 5, e13262. [Google Scholar] [CrossRef]
- Parker, V.J.; Harjes, L.M.; Dembek, K.; Young, G.S.; Chew, D.J.; Toribio, R.E. Association of Vitamin D Metabolites with Parathyroid Hormone, Fibroblast Growth Factor-23, Calcium, and Phosphorus in Dogs with Various Stages of Chronic Kidney Disease. J. Vet. Intern. Med. 2017, 31, 791–798. [Google Scholar] [CrossRef]
- Osuga, T.; Nakamura, K.; Morita, T.; Lim, S.Y.; Nisa, K.; Yokoyama, N.; Sasaki, N.; Morishita, K.; Ohta, H.; Takiguchi, M. Vitamin D Status in Different Stages of Disease Severity in Dogs with Chronic Valvular Heart Disease. J. Vet. Intern. Med. 2015, 29, 1518–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, M.S.; Rassnick, K.M.; Wakshlag, J.J.; Gelzer, A.R.M.; Waxman, A.S.; Struble, A.M.; Refsal, K. Relation of Vitamin D status to congestive heart failure and cardiovascular events in dogs. J. Vet. Intern. Med. 2014, 28, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidner, N.; Woods, J.P.; Conlon, P.; Meckling, K.A.; Atkinson, J.L.; Bayle, J.; Makowski, A.J.; Horst, R.L.; Verbrugghe, A. Influence of Various Factors on Circulating 25(OH) Vitamin D Concentrations in Dogs with Cancer and Healthy Dogs. J. Vet. Intern. Med. 2017, 31, 1796–1803. [Google Scholar] [CrossRef]
- Titmarsh, H.F.; Cartwright, J.A.; Kilpatrick, S.; Gaylor, D.; Milne, E.M.; Berry, J.L.; Bommer, N.X.; Gunn-Moore, D.; Reed, N.; Handel, I.; et al. Relationship between vitamin D status and leukocytes in hospitalised cats. J. Feline Med. Surg. 2017, 19, 364–369. [Google Scholar] [CrossRef]
- Handel, I.; Watt, K.A.; Pilkington, J.G.; Pemberton, J.M.; MacRae, A.; Scott, P.; McNeilly, T.N.; Berry, J.L.; Clements, D.N.; Nussey, D.H.; et al. Vitamin D status predicts reproductive fitness in a wild sheep population. Sci. Rep. 2016, 6, 18986. [Google Scholar] [CrossRef] [Green Version]
- Tuckey, R.C.; Cheng, C.Y.S.; Slominski, A.T. The serum vitamin D metabolome: What we know and what is still to discover. J. Steroid Biochem. Mol. Biol. 2019, 186, 4–21. [Google Scholar] [CrossRef]
- Kaufmann, M.; Sepiashvili, L.; Singh, R.J. Volume One: Biochemistry, Physiology and Diagnostics. In Vitamin D, 4th ed.; Feldman, D., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 909–923. ISBN 9780128099667. [Google Scholar]
- Yin, S.; Yang, Y.; Wu, L.; Li, Y.; Sun, C. Recent advances in sample preparation and analysis methods for vitamin D and its analogues in different matrices. Trends Anal. Chem. 2019, 110, 204–220. [Google Scholar] [CrossRef]
- Jenkinson, C.; Taylor, A.E.; Hassan-smith, Z.K.; Adams, J.S.; Stewart, P.M.; Hewison, M.; Keevil, B.G. High throughput LC – MS / MS method for the simultaneous analysis of multiple vitamin D analytes in serum. J. Chromatogr. B 2016, 1014, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, G.; Kaufmann, M. Vitamin D metabolite profiling using liquid chromatography–tandem mass spectrometry (LC–MS/MS). J. Steroid Biochem. Mol. Biol. 2016, 164, 110–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.J.; Volmer, D.A. Mass spectrometric profiling of vitamin D metabolites beyond 25-Hydroxyvitamin D. Clin. Chem. 2015, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardwell, G.; Bornman, J.F.; James, A.P.; Black, L.J. A Review of Mushrooms as a Potential Source of Dietary Vitamin D. Nutrients 2018, 10, 1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, L.J.; Lucas, R.M.; Sherriff, J.L.; Björn, L.O.; Bornman, J.F. In Pursuit of Vitamin D in Plants. Nutrients 2017, 9, 136. [Google Scholar] [CrossRef] [Green Version]
- Jäpelt, R.B.; Jakobsen, J. Vitamin D in plants: A review of occurrence, analysis, and biosynthesis. Front. Plant Sci. 2013, 4, 136. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D Metabolism, Mechanism of Action, and Clinical Applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.R. Who, what, where and when—Influences on cutaneous vitamin D synthesis. Prog. Biophys. Mol. Biol. 2006, 92, 17–25. [Google Scholar] [CrossRef]
- Bouillon, R.; Eisman, J.; Garabedian, M.; Holick, M.; Kleinschmidt, J.; Suda, T.; Terenetskaya, I.; Webb, A. Action spectrum for the production of previtamin D3 in human skin. UDC 2006, 612, 481–506. [Google Scholar]
- Holick, M.F.; Maclaughlin, J.; Clark, M.; Holick, S.; Potts, J.; Anderson, R.; Blank, I.; Parrish, J.; Elias, P. Photosynthesis of Previtamin D3 in Human Skin and the Physiologic Consequences. Science 1980, 210, 203–205. [Google Scholar] [CrossRef]
- Holick, M.F.; Richtand, N.M.; McNeill, S.C.; Holick, S.A.; Frommer, J.E.; Henley, J.W.; John, T.; Potts, J. Isolation and identification of previtamin D3 from the skin of rats exposed to ultraviolet irradiation. Biochemistry 1979, 18. [Google Scholar] [CrossRef] [PubMed]
- MacLaughlin, J.A.; Anderson, R.R.; Holick, M.F. Spectral character of sunlight modulates photosynthesis of previtamin D3 and its photoisomers in human skin. Science 1982, 216, 1001–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuprykova, O.; Chenc, X.; Hocherc, C.-F.; Skobloa, R.; Yinc, L.; Hocherb, B. Why should we measure free 25 (OH) vitamin D? J. Steroid Biochem. Mol. Biol. 2018, 180, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Farrell, C.L.; Pusceddu, I.; Fabregat-cabello, N. Assessment of vitamin D status—A changing landscape. Clin. Chemsitry Lab. Med. 2017, 55, 3–26. [Google Scholar] [CrossRef]
- Bikle, D.D.; Gee, E.; Halloran, B.; Kowalski, M.A.N.N.; Ryzen, E.; Haddad, J.G. Assessment of the Free Fraction of 25-Hydroxyvitamini n Serum and Its Regulation by Albumin and the Vitamin D-Binding Protein. J. Clin. Endocrinol. Metab. 1986, 63, 954–959. [Google Scholar] [CrossRef]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.G.; Ochalek, J.T.; Kaufmann, M.; Jones, G.; Deluca, H.F. CYP2R1 is a major, but not exclusive, contributor to 25-hydroxyvitamin D production in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 15650–15655. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.B.; Levine, M.A.; Bell, N.H.; Mangelsdorf, D.J.; Russell, D.W. Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase. Proc. Natl. Acad. Sci. USA 2004, 101, 7711–7715. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, D.; Bland, R.; Walker, E.A.; Bradwell, A.R.; Howie, A.J.; Hewison, M.; Stewart, P.M. Expression of 25-Hydroxyvitamin D 3-1 alpha-Hydroxylase in the Human Kidney. J. Am. Soc. Nephrol. 1999, 10, 2465–2473. [Google Scholar]
- Adams, J.S.; Hewison, M. Extrarenal expression of the 25-hydroxyvitamin D-1-hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewison, M.; Burke, F.; Evans, K.N.; Lammas, D.A.; Sansom, D.M.; Liu, P.; Modlin, R.L.; Adams, J.S. Extra-renal 25-hydroxyvitamin D 3-1 alpha-hydroxylase in human health and disease. J. Steroid Biochem. Mol. Biol. 2007, 103, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, D.; Bland, R.; Williams, M.C.; Ninch, R.W.M.C.; Howie, A.J.; Stewart, P.M.; Hewison, M. Extrarenal Expression of 25-Hydroxyvitamin D 3-1 alpha-Hydroxylase. J. Clin. Endocrinol. Metab. 2001, 86, 888–894. [Google Scholar] [PubMed] [Green Version]
- Haussler, M.R.; Jurutka, P.W.; Mizwicki, M.; Norman, A.W. Vitamin D receptor (VDR)-mediated actions of 1a,25(OH)2vitamin D3: Genomic and non-genomic mechanisms. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 543–559. [Google Scholar] [CrossRef]
- Brenza, H.L.; Deluca, H.F. Regulation of 25-Hydroxyvitamin D 3 1 a-Hydroxylase Gene Expression by Parathyroid Hormone and. Arch. Biochem. Biophys. 2000, 381, 143–152. [Google Scholar] [CrossRef]
- Murayama, A.; Takeyama, K.-I.; Kitanaka, S.; Kodera, Y.; Kawaguchi, Y.; Hosoya, T.; Kato, S. Positive and Negative Regulations of the Renal 25-Hydroxyvitamin D3 1a-Hydroxylase Gene by Parathyroid Hormone, Calcitonin, and 1a,25(OH)2D3 in Intact Animals. Endocrinology 1999, 140, 2224–2231. [Google Scholar] [CrossRef]
- Murayama, A.; Takeyama, K.; Kitanaka, S.; Kodera, Y.; Hosoya, T.; Kato, S. The Promoter of the Human 25-Hydroxyvitamin D3 1a-Hydroxylase Gene Confers Positive and Negative Responsiveness to PTH, Calcitonin, and 1a,25(OH)2D3. Biochem. Biophys. Res. Commun. 1998, 16, 11–16. [Google Scholar] [CrossRef]
- Shimada, T.; Kakitani, M.; Yamazaki, Y.; Hasegawa, H.; Takeuchi, Y.; Fujita, T.; Fukumoto, S.; Tomizuka, K. Targeted ablation of Fgf23 demonstrates an essential physiological role of FGF23 in phosphate and vitamin D metabolism. J. Clin. Investig. 2004, 113. [Google Scholar] [CrossRef]
- Boyle, I.T.; Gray, R.W.; Deluca, H.F. Regulation by Calcium of In Vivo Synthesis of 1,25-Dihydroxycholecalciferol. Proc. Natl. Acad. Sci. USA 1971, 68, 2131–2134. [Google Scholar] [CrossRef] [Green Version]
- Prosser, D.E.; Jones, G. Enzymes involved in the activation and inactivation of vitamin D. Trends Biochem. Sci. 2004, 29. [Google Scholar] [CrossRef]
- Beckman, M.J.; Tadikonda, P.; Werner, E.; Prahl, J.; Yamada, S.; DeLuca, H.F. Human 25-Hydroxyvitamin D3-24-Hydroxylase, a Multicatalytic Enzyme. Biochemistry 1996, 35, 8465–8472. [Google Scholar] [CrossRef] [PubMed]
- Prosser, D.E.; Kaufmann, M.; O’Leary, B.; Byford, V.; Jones, G. Single A326G mutation converts human CYP24A1 from 25-OH-D3-24-hydroxylase into -23-hydroxylase, generating 1α,25-(OH)2D3-26,23-lactone. Proc. Natl. Acad. Sci. USA 2007, 104, 12673–12678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, G.S.; Tserng, K.Y. Calcitroic acid, end product of renal metabolism of 1,25-dihydroxyvitamin D3 through the C-24 oxidation pathway. Biochemistry 1989, 28, 1763–1769. [Google Scholar] [CrossRef]
- St-Arnaud, R.; Jones, G. CYP24A1: Structure, Function, and Physiological Role. In Vitamin D: Fourth Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2017; Volume 1, pp. 81–95. ISBN 9780128099667. [Google Scholar]
- Martineau, C.; Jones, G.; St-arnaud, R.; Martineau, C.; Naja, R.P.; Husseini, A.; Hamade, B.; Kaufmann, M.; Akhouayri, O.; Arabian, A.; et al. Optimal bone fracture repair requires 24R,25-dihydroxyvitamin D 3 and its effector molecule Find the latest version: Optimal bone fracture repair requires 24R,25-dihydroxyvitamin D 3 and its effector molecule FAM57B2. J. Clin. Investig. 2018, 128, 3546–3557. [Google Scholar] [CrossRef] [Green Version]
- Boyan, B.D.; Hyzy, S.L.; Pan, Q.; Scott, K.M.; Coutts, R.D.; Healey, R.; Schwartz, Z. 24R,25-Dihydroxyvitamin D3 Protects against Articular Cartilage Damage following Anterior Cruciate Ligament Transection in Male Rats. PLoS ONE 2016, 11, e161782. [Google Scholar] [CrossRef] [Green Version]
- Boyan, B.D.; Hurst-kennedy, J.; Denison, T.A.; Schwartz, Z. 24R,25-Dihydroxyvitamin D3 [24R,25(OH)2D3] controls growth plate development by inhibiting apoptosis in the reserve zone and stimulating response to 1_,25(OH)2D3 in hypertrophic cells. J. Steroid Biochem. Mol. Biol. 2010, 121, 212–216. [Google Scholar] [CrossRef]
- St-arnaud, R. CYP24A1-deficient mice as a tool to uncover a biological activity for vitamin D metabolites hydroxylated at position 24. J. Steroid Biochem. Mol. Biol. 2010, 121, 254–256. [Google Scholar] [CrossRef]
- Engstrom, G.W.; Reinhardt, T.A.; Horst, R.L. 25-Hydroxyvitamin D3-23-hydroxylase, a renal enzyme in several animal species. Arch. Biochem. Biophys. 1986, 250, 86–93. [Google Scholar] [CrossRef]
- Pedersen, J.I.; Hagenfeldt, Y.; Björkhem, I. Assay and properties of 25-hydroxyvitamin D3 23-hydroxylase. Evidence that 23,25-dihydroxyvitamin D3 is a major metabolite in 1,25-dihydroxyvitamin D3-treated or fasted guinea pigs. Biochem. J. 1988, 250, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.; Prosser, D.E.; Kaufmann, M. 25-Hydroxyvitamin D-24-hydroxylase (CYP24A1): Its important role in the degradation of vitamin D. Arch. Biochem. Biophys. 2012, 523, 9–18. [Google Scholar] [CrossRef]
- Hamamoto, H.; Kusudo, T.; Urushino, N.; Masuno, H.; Yamamoto, K.; Yamada, S.; Kamakura, M.; Ohta, M.; Inouye, K.; Sakaki, T. Structure-Function Analysis of Vitamin D 24-Hydroxylase (CYP24A1) by Site-Directed Mutagenesis: Amino Acid Residues Responsible for Species-Based Difference of CYP24A1 between Humans and Rats. Mol. Pharmacol. 2006, 70, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, N.; Saikatsu, S.; Akeno, N.; Abe, M.; Kimura, S.; Yamada, S. Synthesis of 25-Hydroxyvitamin D3-26,23-Lactone but not 24,25-Dihydroxyvitamin D3 from 25-Hydroxyvitamin D3 in Opossum Kidney Cells Treated with 1α,25-Dihydroxyvitamin D3. Horm. Metab. Res. 1995, 27, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Simboli-Campbell, M.; Jones, G. Dietary Phosphate Deprivation Increases Renal Synthesis and Decreases Renal Catabolism of 1,25-Dihydroxycholecalciferol in Guinea Pigs. J. Nutr. 1991, 121, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Uhl, E.W. The pathology of vitamin D deficiency in domesticated animals: An evolutionary and comparative overview. Int. J. Paleopathol. 2018, 23, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Suda, T. Vitamin D: Calcium and bone homeostasis during evolution. Bonekey Rep. 2014, 3, 480. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.; Veljkovic, K.; Yazdanpanah, M.; Adeli, K. Analytical measurement and clinical relevance of vitamin D 3 C3-epimer. Clin. Biochem. 2012, 46, 190–196. [Google Scholar] [CrossRef]
- Hurst, E.A.; Homer, N.Z.; Gow, A.G.; Clements, D.N.; Evans, H.; Gaylor, D.; Campbell, S.; Handel, I.; Mellanby, R.J. Vitamin D status is seasonally stable in northern European dogs. Vet. Clin. Pathol. 2020. [Google Scholar] [CrossRef]
- Hurst, E.A.; Homer, N.Z.; Denham, S.G.; MacFarlane, E.; Campbell, S.; Boswinkel, M.; Mellanby, R.J. Development and application of a LC–MS/MS assay for simultaneous analysis of 25-hydroxyvitamin-D and 3-epi-25-hydroxyvitamin-D metabolites in canine serum. J. Steroid Biochem. Mol. Biol. 2020, 199. [Google Scholar] [CrossRef]
- Groth, E.M.; Lulich, J.P.; Chew, D.J.; Parker, V.J. Vitamin D metabolism in dogs with and without hypercalciuric calcium oxalate urolithiasis. J. Vet. Intern. Med. 2019, 33, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Mick, P.J.; Peng, S.A.; Loftus, J.P.; Elisa, C. Serum Vitamin D Metabolites and CXCL10 Concentrations Associate with Survival in Dogs with Immune Mediated Disease. Front. Vet. Sci. 2019, 6, 247. [Google Scholar] [CrossRef] [Green Version]
- Fritz, C.A.; Navetta, K.A.; Wolford, D.P.; Colangelo, J.L. Assessment of endogenous 25-hydroxyvitamin D serum concentrations by liquid chromatography-tandem mass spectrometry in various animal species. Vet. Clin. Pathol. 2017, 46, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprinkle, M.C.; Hooper, S.E.; Backus, R.C. Previously undescribed vitamin D C-3 epimer occurs in substantial amounts in the blood of cats. J. Feline Med. Surg. 2017. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.L.; Hammett-Stabler, C.; Hauck, M.L. Serum 25-hydroxyvitamin D concentrations in dogs with osteosarcoma do not differ from those of age- and weight-matched control dogs. Vet. J. 2016, 217, 132–133. [Google Scholar] [CrossRef] [Green Version]
- Spoo, J.W.; Downey, R.L.; Griffitts, C.; Horst, R.J.; Levine, C.B.; Childs, R.M.; Wakshlag, J.J. Plasma Vitamin D Metabolites and C-Reactive Protein in Stage-Stop Racing Endurance Sled Dogs. J. Vet. Intern. Med. 2015, 29, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Azarpeykan, S.; Dittmer, K.E.; Gee, E.K.; Marshall, J.C.; Wallace, J.; Elder, P.; Acke, E.; Thompson, K.G. Influence of blanketing and season on vitamin D and parathyroid hormone, calcium, phosphorus, and magnesium concentrations in horses in New Zealand. Domest. Anim. Endocrinol. 2016, 56, 75–84. [Google Scholar] [CrossRef]
- Allott, B.S.; Dittmer, K.E.; Kenyon, A.G.; Elder, P.A. Preliminary investigation of the effect of treating sheep during pregnancy with a vitamin A, D, E formulation on the incidence of vaginal prolapse. N. Z. Vet. J. 2020, 68, 193–197. [Google Scholar] [CrossRef]
- Dittmer, K.E.; Surendran, A.; Elder, P.A.; Hoyle, T.J. The effect of serum 25-hydroxyvitamin D concentration on fecundity of ewes on a New Zealand sheep farm. Small Rumin. Res. 2020, 190, 106149. [Google Scholar] [CrossRef]
- Zhou, P.; Mcevoy, T.G.; Andrew, C.G.; Lambe, N.R.; Morgan-davies, C.R.; Hurst, E.; Sargison, N.D.; Mellanby, R.J. Investigation of relationship between vitamin D status and reproductive fitness in Scottish hill sheep. Sci. Rep. 2019, 9, 1162. [Google Scholar] [CrossRef] [Green Version]
- Celi, P.; Williams, S.; Engstrom, M.; Mcgrath, J.; La, J. Safety evaluation of dietary levels of 25-hydroxyvitamin D 3 in growing calves. Food Chem. Toxicol. 2018, 111, 641–649. [Google Scholar] [CrossRef]
- Guo, J.; Jones, A.K.; Givens, D.I.; Lovegrove, J.A.; Kliem, K.E. Effect of dietary vitamin D 3 and 25-hydroxyvitamin D 3 supplementation on plasma and milk 25-hydroxyvitamin D 3 concentration in dairy cows. J. Dairy Sci. 2018, 101, 3545–3553. [Google Scholar] [CrossRef]
- Alexander, B.M.; Ingold, B.C.; Young, J.L.; Fensterseifer, S.R.; Wechsler, P.J.; Austin, K.J.; Larson-Meyer, D.E. Sunlight exposure increases vitamin D sufficiency in growing pigs fed a diet formulated to exceed requirements. Domest. Anim. Endocrinol. 2017, 59, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Kühn, J.; Schutkowski, A.; Hirche, F.; Baur, A.C.; Mielenz, N.; Stangl, G.I. Non-linear increase of vitamin D content in eggs from chicks treated with increasing exposure times of ultraviolet light. J. Steroid Biochem. Mol. Biol. 2015, 148, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Schutkowski, A.; Krämer, J.; Kluge, H.; Hirche, F.; Krombholz, A.; Theumer, T.; Stangl, G.I. UVB Exposure of Farm Animals: Study on a Food-Based Strategy to Bridge the Gap between Current Vitamin D Intakes and Dietary Targets. PLoS ONE 2013, 8, e69418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, T.E.; Kapoor, A.; Hedman, C.J.; Binkley, N.; KEMNITZ, J.W. Measurement of 25-Hydroxyvitamin D2&3 and 1,25-Dihydroxyvitamin D2&3 by Tandem Mass Spectrometry: A Primate Multispecies Comparison. Am. J. Primatol. 2015, 77, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, T.E.; Kapoor, A.; Binkley, N.C.; Rice, K.S.; Rogers, J.; Phillips-conroy, C.J.J.J.E. Comparison of vitamin D metabolites in wild and captive baboons. Am. J. Primatol. 2018, 25, 11935. [Google Scholar] [CrossRef]
- Griffiths, P.; Fairney, A. Vitamin D metabolites in polar vertebrates. Comp. Biochem. Physiol. 1988, 91, 511–516. [Google Scholar]
- Laing, C.J.; Malik, R.; Wigney, D.I.; Fraser, D.R. Seasonal vitamin D status of Greyhounds in Sydney. Aust. Vet. J. 1999, 77, 35–38. [Google Scholar] [CrossRef]
- Morris, J.G. Idiosyncratic nutrient requirements of cats appear to be diet-induced evolutionary adaptations. Nutr. Res. Rev. 2002, 15, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, K.; Rumbeiha, W.K. Pet Food Recalls and Pet Food Contaminants in Small Animals: An Update. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Crossley, V.J.; Bovens, C.P.V.; Pineda, C.; Hibbert, A.; Finch, N.C. Vitamin D toxicity of dietary origin in cats fed a natural complementary kitten food. J. Feline Med. Surg. Open Reports 2017. [Google Scholar] [CrossRef]
- Mellanby, R.J.; Mee, A.P.; Berry, J.L.; Herrtage, M.E. Hypercalcaemia in two dogs caused by excessive dietary supplementation. J. Small Anim. Pract. 2005, 46, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Kritikos, G.; Weidner, N.; Atkinson, J.L.; Bayle, J.; van Hoek, I.; Verbrugghe, A. Quantification of vitamin D 3 in commercial dog foods and comparison with Association of American and manufacturer-reported concentrations. J. Am. Vet. Med. Assoc. 2018, 252, 1521–1526. [Google Scholar] [CrossRef] [PubMed]
- Young, L.R.; Backus, R.C. Oral vitamin D supplementation at five times the recommended allowance marginally affects serum 25-hydroxyvitamin D concentrations in dogs. J. Nutr. Sci. 2016, 25, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA U.S. Recalls, Market Withdrawels and Safety Alerts. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts (accessed on 9 January 2020).
- Morris, B.J.G. Cats discriminate between cholecalciferol and ergocalciferol. J. Anim. Physiol. Anim. Nutr. Anim. Nutr. 2002, 86, 229–238. [Google Scholar] [CrossRef]
- Sih, T.R.; Morris, J.G.; Hickman, M.A. Chronic ingestion of high concentrations of cholecalciferol in cats. Am. J. Vet. Res. 2001, 62, 1500–1506. [Google Scholar] [CrossRef]
- El Shorafa, W.M.; Feaster, J.P.; Ott, E.A.; Asquith, R. K effect of vitamin d and sunlight on growth and bone development of young ponies. J. Anim. Sci. 1979, 48, 882–886. [Google Scholar] [CrossRef] [Green Version]
- Breidenbach, A.; Schlumbohm, C.; Harmeyer, J. Peculiarities of vitamin D and of the calcium and phosphate homeostatic system in horses. Vet. Res. 1998, 29, 173–186. [Google Scholar]
- Maenpaa, P.H.; Koskinen, T.; Koskinen, E. Serum profiles of vitamins A, E and D in mares and foals during different seasons. J. Anim. Sci. 1988, 66, 1418–1423. [Google Scholar] [CrossRef] [Green Version]
- Pozza, M.E.; Kaewsakhorn, T.; Trinarong, C.; Inpanbutr, N.; Toribio, R.E. Serum vitamin D, calcium, and phosphorus concentrations in ponies, horses and foals from the United States and Thailand. Vet. J. 2014, 199, 451–456. [Google Scholar] [CrossRef]
- Wilkens, M.R.; Marholt, L.; Eigendorf, N.; Muscher-banse, A.S.; Feige, K.; Schröder, B.; Breves, G.; Cehak, A. Trans- and paracellular calcium transport along the small and large intestine in horses. Comp. Biochem. Physiol. Part A 2017, 204, 157–163. [Google Scholar] [CrossRef]
- Toribio, R.E.; Kohn, C.W.; Rourke, K.M.; Levine, A.L.; Rosol, T.J. Effects of hypercalcemia on serum concentrations of magnesium, potassium, and phosphate and urinary excretion of electrolytes in horses. Am. J. Vet. Res. 2007, 68, 543–554. [Google Scholar] [CrossRef]
- Toribio, R.E.; Kohn, C.W.; Chew, D.J.; Sams, R.A.; Rosol, T.J. Comparison of serum parathyroid hormone and ionized calcium and magnesium concentrations and fractional urinary clearance of calcium and phosphorus in healthy horses and horses with enterocolitis. Am. J. Vet. Res. 2001, 62, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.; Brownlow, M.; Hutchins, D. Indices of renal function: Reference values in normal horses. Aust. Vet. J. 1989, 66, 60–63. [Google Scholar] [CrossRef]
- Rourke, K.M.; Coe, S.; Kohn, C.W.; Rosol, T.J.; Mendoza, F.J.; Toribio, R.E. Cloning, comparative sequence analysis and mRNA expression of calcium-transporting genes in horses. Gen. Comp. Endocrinol. 2010, 167, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Azarpeykan, S.; Dittmer, K.E.; Marshall, J.C.; Perera, K.C.; Gee, K.; Acke, E.; Thompson, K.G. Evaluation and Comparison of Vitamin D Responsive Gene Expression in Ovine, Canine and Equine Kidney. PLoS ONE 2016, 11, e0162598. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.; Wright, H. Relative contributions of diet and sunshine to the overall vitamin D status of the grazing ewe. Vet. Rec. 1984, 115, 537–538. [Google Scholar] [CrossRef]
- Piccione, G.; Assenza, A.; Fazio, F.; Bergero, D.; Caola, G. Daily Rhythms of Serum Vitamin D—Metabolites, Calcium and Phosphorus in Horses. Acta Vet. Brun. 2008, 77, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, L.Y.; Wortsman, J.; Dannenberg, M.J.; Hollis, B.W.; Lu, Z.; Holick, M.F. Clothing prevents ultraviolet-B radiation-dependent photosynthesis of vitamin D3. J. Clin. Endocrinol. Metab. 1992, 75, 1099–1103. [Google Scholar] [CrossRef]
- Salih, F.M. Effect of clothing varieties on solar photosynthesis of previtamin D3: An in vitro study. Photodermatol. Photoimmunol. Photomed. 2004, 20, 53–58. [Google Scholar] [CrossRef]
- Hymøller, L.; Jensen, S.K. Vitamin D3 synthesis in the entire skin surface of dairy cows despite hair coverage. J. Dairy Sci. 2010, 93, 2025–2029. [Google Scholar] [CrossRef] [Green Version]
- Clemens, T.L.; Henderson, S.L.; Adams, J.S.; Holick, M.F. Increased skin pigment reduces the capacity of skin to synthesise vitamin D3. Lancet 1982, 319, 74–76. [Google Scholar] [CrossRef]
- Azarpeykan, S.; Dittmer, K.E.; Gee, E.K.; Marshall, J.C.; Elder, P.; Acke, E.; Thompson, K.G. Circadian rhythm of calciotropic hormones, serum calcium, phosphorus and magnesium during the shortest and longest days of the year in horses in New Zealand. J. Anim. Physiol. Anim. Nutr. (Berl.) 2016, 100, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, E.; Dede, S.; Deger, Y.; Yoruk, I. Investigation of the effects of carrying heavy load on prooxidation / antioxidation status and vitamin D3 in healthy horses. Asian J. Anim. Vet. Adv. 2009, 4, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Harmeyer, J.; Schlumbohm, C. Effects of pharmacological doses of Vitamin D3 on mineral balance and profiles of plasma Vitamin D3 metabolites in horses. J. Steroid Biochem. Mol. Biol. 2004, 90, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Kamr, A.M.; Dembek, K.A.; Reed, S.M.; Slovis, N.M.; Zaghawa, A.; Rosol, T.J.; Toribio, R.E. Vitamin D Metabolites and Their Association with Calcium, Phosphorus, and PTH Concentrations, Severity of Illness, and Mortality in Hospitalized Equine Neonates. PLoS ONE 2015, 25, e0127684. [Google Scholar] [CrossRef] [Green Version]
- Kamr, A.M.; Dembek, K.A.; Hildreth, B.E.; Morresey, P.R.; Rathgeber, R.A.; Burns, T.A.; Zaghawa, A.A.; TORIBIO, R.E. The FGF-23/klotho axis and its relationship with phosphorus, calcium, vitamin D, PTH, aldosterone, severity of disease, and outcome in hospitalised foals. Equine Vet. J. 2018, 50, 739–746. [Google Scholar] [CrossRef]
- Nemeth, M.V.; Wilkens, M.R.; Liesegang, A. Vitamin D status in growing dairy goats and sheep: Influence of ultraviolet B radiation on bone metabolism and calcium homeostasis. J. Dairy Sci. 2017, 100, 8072–8086. [Google Scholar] [CrossRef] [Green Version]
- Mearns, R.; Scholes, S.F.E.; Wessels, M.; Whitaker, K.; Strugnell, B. Rickets in sheep flocks in northern England. Veterninary Rec. 2008, 162, 98–99. [Google Scholar] [CrossRef]
- Smith, B.S.; Wright, H. Seasonal variation in serum 25-hydroxyvitamin D concentrations in sheep. Vet. Rec. 1981, 109, 139–141. [Google Scholar] [CrossRef]
- Smith, B.S.; Wright, H.; Brown, K.G. Effect of vitamin D supplementation during pregnancy on the vitamin D status of ewes and their lambs. Vet. Rec. 1987, 120, 199–201. [Google Scholar] [CrossRef]
- Herm, G.; Breves, G.; Schröder, B.; Wilkens, M.R. Renal mechanisms of calcium homeostasis in sheep and goats. J. Anim. Sci. 2015, 1608–1621. [Google Scholar] [CrossRef] [PubMed]
- Kohler, M.; Leiber, F.; Willems, H.; Merbold, L.; Liesegang, A. Influence of altitude on vitamin D and bone metabolism of lactating sheep and goats. J. Anim. Sci. 2013, 91, 5259–5268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, K.G.; Dittmer, K.E.; Blair, H.T.; Fairley, R.A.; Sim, D.F.W. An outbreak of rickets in Corriedale sheep: Evidence for a genetic aetiology An outbreak of rickets in Corriedale sheep: Evidence for a genetic aetiology. N. Z. Vet. J. 2007, 55, 137–142. [Google Scholar] [CrossRef]
- Dittmer, K.E.; Thompson, K.G.; Blair, H.T. Pathology of Inherited Rickets in Corriedale Sheep. J. Comp. Pathol. 2009, 141, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dittmer, K.E.; Blair, H.T.; Thompson, K.G.; Rothschild, M.F.; Garrick, D.J. A Novel Nonsense Mutation in the DMP1 Gene Identified by a Genome-Wide Association Study Is Responsible for Inherited Rickets in Corriedale Sheep. PLoS ONE 2011, 6, e21739. [Google Scholar] [CrossRef]
- Saun, R.J. Van Vitamin D-responsive rickets in neonatal lambs. Can. Vet. J. 2004, 45, 841–844. [Google Scholar] [PubMed]
- Yao, X.; Ei-samahy, M.A.; Yang, H.; Feng, X.; Li, F.; Mengb, F.; Niea, H.; Wanga, F. Age-associated expression of vitamin D receptor and vitamin D-metabolizing enzymes in the male reproductive tract and sperm of Hu sheep. Anim. Reprod. Sci. 2018, 190, 27–38. [Google Scholar] [CrossRef]
- Goyal, R.; Billings, T.L.; Mansour, T.; Martin, C.; Baylink, D.J.; Longo, L.D.; Pearce, W.J.; Mata-greenwood, E. Vitamin D status and metabolism in an ovine pregnancy model: Effect of long-term, high-altitude hypoxia. Am. J. Physiol. Endocrinol. Metab. 2016, 310. [Google Scholar] [CrossRef] [Green Version]
- Casas, E.; Lippolis, J.D.; Kuehn, L.A.; Reinhardt, T.A. Seasonal variation in vitamin D status of beef cattle reared in the central United States. Domest. Anim. Endocrinol. 2015, 52, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.D.; Powell, J.L.; Price, D.M.; Hersom, M.J.; Yelich, J.V.; Drewnoski, M.E.; Bird, S.L.; Bridges, G.A. Assessment of serum 25-hydroxyvitamin D concentrations of beef cows and calves across seasons and geographical locations. J. Anim. Sci. 2016, 94, 3958–3965. [Google Scholar] [CrossRef]
- Weir, R.R.; Strain, J.J.; Johnston, M.; Lowis, C.; Fearon, A.M.; Ireland, N.; Bt, C. Environmental and genetic factors in fl uence the vitamin D content of cows’ milk Proceedings of the Nutrition Society Proceedings of the Nutrition Society. Proc. Nutr. Soc. 2017, 76, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holcombe, S.J.; Wisnieski, L.; Gandy, J.; Norby, B.; Sordillo, L.M. Reduced serum vitamin D concentrations in healthy early-lactation dairy cattle. J. Dairy Sci. 2018, 101, 1488–1494. [Google Scholar] [CrossRef] [PubMed]
- Merriman, K.E.; Poindexter, M.B.; Kweh, M.F.; Santos, J.E.P.; Nelson, C.D. Intramammary 1,25-dihydroxyvitamin D3 treatment increases expression of host-defense genes in mammary immune cells of lactating dairy cattle. J. Steroid Biochem. Mol. Biol. 2017, 173, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Corripio-Miyar, Y.; Mellanby, R.J.; Morrison, K.; McNeilly, T.N. 1,25-Dihydroxyvitamin D3 modulates the phenotype and function of Monocyte derived dendritic cells in cattle. BMC Vet. Res. 2017, 13, 390. [Google Scholar] [CrossRef]
- Yue, Y.; Hymøller, L.; Jensen, S.K.; Lauridsen, C.; Purup, S. Effects of vitamin D and its metabolites on cell viability and Staphylococcus aureus invasion into bovine mammary epithelial cells. Vet. Microbiol. 2017, 203, 245–251. [Google Scholar] [CrossRef]
- García-Barragán, Á.; Gutiérrez-Pabello, J.A.; Alfonseca-Silva, E. Calcitriol increases nitric oxide production and modulates microbicidal capacity against Mycobacterium bovis in bovine macrophages. Comp. Immunol. Microbiol. Infect. Dis. 2018, 59, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Waters, W.R.; Nonnecke, B.J.; Rahner, T.E.; Palmer, M.V.; Whipple, D.L.; Horst, R.L. Modulation of Mycobacterium bovis-Specific Responses of Bovine Peripheral Blood Mononuclear Cells by 1,25-Dihydroxyvitamin D3. Clin. Diagn. Lab. Immunol. 2001, 8, 1204–1212. [Google Scholar] [CrossRef] [Green Version]
- Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.A.; Nonnecke, B.J.; Nelson, C.D. Treatment of an Intramammary Bacterial Infection with 25-Hydroxyvitamin D3. PLoS ONE 2011, 6, e25479. [Google Scholar] [CrossRef]
- Sacco, R.E.; Nonnecke, B.J.; Palmer, M.V.; Waters, W.R.; Lippolis, J.D.; Reinhardt, T.A. Differential Expression of Cytokines in Response to Respiratory Syncytial Virus Infection of Calves with High or Low Circulating 25-Hydroxyvitamin D3. PLoS ONE 2012, 7, e33074. [Google Scholar] [CrossRef] [Green Version]
- Yue, Y.; Hymøller, L.; Jensen, S.K.; Lauridsen, C. Effect of vitamin D treatments on plasma metabolism and immune parameters of healthy dairy cows. Arch. Anim. Nutr. 2018, 72, 205–220. [Google Scholar] [CrossRef]
- Madson, D.M.; Ensley, S.M.; Gauger, P.C.; Schwartz, K.J.; Stevenson, G.W.; Cooper, V.L.; Janke, B.H.; Burrough, E.R.; Goff, J.P.; Horst, R.L. Rickets: Case series and diagnostic review of hypovitaminosis D in swine. J. Vet. Diagnostic Investig. 2012, 24, 1137–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolp, E.; Wilkens, M.R.; Pendl, W.; Eichenberger, B.; Liesegang, A. Vitamin D metabolism in growing pigs: Influence of UVB irradiation and dietary vitamin D supply on calcium homeostasis, its regulation and bone metabolism. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flohr, J.R.; Woodworth, J.C.; Bergstrom, J.R. Evaluating the impact of maternal vitamin D supplementation: I. Sow performance, serum vitamin metabolites, and neonatal muscle characteristics. J. Anim. Sci. 2016, 25, 4629–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flohr, J.R.; Woodworth, J.C.; Bergstrom, J.R. Evaluating the impact of maternal vitamin D supplementation on sow performance: II. Subsequent growth performance and carcass characteristics of growing pigs. J. Anim. Sci. 2016, 4643–4653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, L.S.; Serda, R.; Choe, S.; Davidi, L.; Harmeyer, J.; Omdahl, J.L. Molecular basis for pseudo vitamin D-deficiency rickets in the Hannover pig. J. Nutr. Biochem. 2003, 14, 378–385. [Google Scholar] [CrossRef]
- Kaune, R.; Harmeyer, J. Vitamin D3 metabolism in a pig strain with pseudo vitamin D-deficiency rickets, type I. Acta Endocrinol. (Copenh.) 1987, 115, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Maunder, E.M.W.; Randall, V.A.; Care, A.D. Vitamin D-dependent rickets type I in pigs. Clin. Sci. 1985, 69, 541–548. [Google Scholar] [CrossRef]
- Dinev, I. Clinical and morphological investigations on the incidence of forms of rickets and their association with other pathological states in broiler chickens. Res. Vet. Sci. 2012, 92, 273–277. [Google Scholar] [CrossRef]
- Edwards, H.M. Nutrition and Skeletal Problems in Poultry. Poult. Sci. 2000, 79, 1018–1023. [Google Scholar] [CrossRef]
- Nys, Y.; Le Roy, N. Calcium Homeostasis and Eggshell Biomineralization in Female Chicken. In Vitamin D: Fourth Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 1, pp. 361–382. ISBN 9780128099667. [Google Scholar]
- Narbaitz, R.; Tsang, C.P.W.; Grunder, A.A. Effects of Vitamin D Deficiency in the Chicken Embryo. Calcif. Tissue Int. 1987, 40, 109–113. [Google Scholar] [CrossRef]
- Nys, Y.; Van Baelen, H.; Bouillon, R. Plasma 1, 25 dihydroxycholecalciferol and its free index are potentiated by ovulation dependent factors and shell formation induced hypocalcemia in the laying hens. Domest. Anim. Endocrinol. 1992, 9, 37–47. [Google Scholar] [CrossRef]
- Abe, E.; Tanabe, R.; Suda, T.; Yoshiki, S.; Horikawa, H.; Masumura, T.; Sugahara, M. Circadian rhythm of La,25-dihydroxyvitamin D3 production in egg-laying hens. Biochem. Biophys. Res. Commun. 1979, 88, 500–507. [Google Scholar] [CrossRef]
- Sakkas, P.; Smith, S.; Hill, T.R.; Kyriazakis, I. A reassessment of the vitamin D requirements of modern broiler genotypes. Poult. Sci. 2019, 98, 330–340. [Google Scholar] [CrossRef]
- Vazquez, J.R.; Gómez, G.V.; López, C.C.; Cortés, A.C.; Diaz, A.C.; Fernández, S.R.T.; Rosales, E.M.; Avila, A.G. Effects of 25-hydroxycholecalciferol with two D3 vitamin levels on production and immunity parameters in broiler chickens. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 25, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, M.; Yalçin, S.; Koçer, B.; Tüzün, A.E.; Akşit, H.; Özkan, S.; Uygun, M.; Güven, G.; Yildiz, O. Effects of enhancing vitamin D status by 25-hydroxycholecalciferol supplementation, alone or in combination with calcium and phosphorus, on sternum mineralisation and breast meat quality in broilers. Br. Poult. Sci. 2017, 58, 452–461. [Google Scholar] [CrossRef]
- Hutton, K.C.; Vaughn, M.A.; Litta, G.; Turner, B.J.; Starkey, J.D. Effect of vitamin D status improvement with 25-hydroxycholecalciferol on skeletal muscle growth characteristics and satellite cell activity in broiler chickens. J. Anim. Sci. 2014, 3291–3299. [Google Scholar] [CrossRef] [PubMed]
- Browning, L.C.; Cowieson, A.J. Vitamin D fortification of eggs for human health. J. Sci. Food Agric. 2014, 94, 1389–1396. [Google Scholar] [CrossRef]
- Mattila, P.; Valaja, J.; Rossow, L.; Venalainen, E.; Tupasela, T. Effect of Vitamin D2- and D3-Enriched Diets on Egg Vitamin D Content, Production, and Bird Condition during an Entire Production Period. Poult. Sci. 2004, 83, 433–440. [Google Scholar] [CrossRef]
- DeLuca, H.F.; Nakada, M.; Tanaka, Y.; Sicinski, R.; Mary, P. The plasma binding protein for vitamin D is a site of discrimination against vitamin D-2 compounds by the chick. Biochim. Biophys. Acta 1988, 965, 16–21. [Google Scholar] [CrossRef]
- Kühn, J.; Schutkowski, A.; Kluge, H.; Hirche, F.; Stangl, G.I. Free-range farming: A natural alternative to produce vitamin D-enriched eggs. Nutrition 2014, 30, 481–484. [Google Scholar] [CrossRef]
- Kuhn, J.; Wassermann, C.; Ebschke, S.; Schutkowski, A.; Thamm, K.; Wensch-dorendorf, M.; Von Borell, E.; Stangl, G.I. Feasibility of artificial light regimes to increase the vitamin D content in indoor-laid eggs. Poult. Sci. 2019, 98, 5177–5187. [Google Scholar] [CrossRef] [PubMed]
- Lietzow, J.; Kluge, H.; Brandsch, C.; Seeburg, N.; Hirche, F.; Glomb, M.; Stangl, G.I. Effect of Short-Term UVB Exposure on Vitamin D Concentration of Eggs and Vitamin D Status of Laying Hens. J. Agric. Food Chem. 2012, 60, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Lumeij, J.T. Chapter 28—Birds. In Medical History and Physical Examination in Companion Animals; Rijnberk, A., van Sluijs, F.J., Belshaw, B.E., Janssen, B., Fama, J., Teske, E., Mudde, M.J.A., Schaefers, E., van Blokland, R.N., Pollak, Y.W.E.A.B.T.-M.H., Eds.; W.B. Saunders: Edinburgh, UK, 2009; pp. 247–271. ISBN 978-0-7020-2968-4. [Google Scholar]
- Mayer, J.; Donnelly, T.M.B.T.-C.V.A. Uropygial Gland Disease Conditions. In Clinical Veterinary Advisor: Birds and Exotic Pets; Mayer, J., Donnelly, T.M.B.T.-C.V.A., Eds.; W.B. Saunders: Saint Louis, MO, USA, 2013; pp. 237–239. ISBN 978-1-4160-3969-3. [Google Scholar]
- Rosenberg, H.R. The site and nature of pro vitamin D in birds. Arch. Biochem. 1953, 42, 7–11. [Google Scholar] [CrossRef]
- Apandi, M.; Edwards, H.M. Studies on the Composition of the Secretions of the Uropygial Gland of Some Avian Species1,2. Poult. Sci. 1964, 43, 1445–1462. [Google Scholar] [CrossRef]
- Salibian, A.; Montalti, D. Physiological and biochemical aspects of the avian uropygial gland. Braz. J. Biol. 2009, 69, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Van Saun, R.J. Nutritional diseases of South American camelids. Small Rumin. Res. 2006, 61, 153–164. [Google Scholar] [CrossRef]
- Smith, B.B.; Van Saun, R.J. Seasonal changes in serum calcium, phosphorus, and vitamin D concentrations in llamas and alpacas. Am. J. Vet. Res. 2001, 62, 1187–1193. [Google Scholar] [CrossRef]
- Van Saun, R.J. Evaluation of vitamin D status of llamas and alpacas with hypophosphatemic rickets. J. Am. Vet. Med. Assoc. 1996, 209, 1128–1133. [Google Scholar]
- Udson, G.J.; Partington, D.L. Factors associated with low vitamin D status of Australian alpacas. Aust. Vet. J. 2008, 86, 486–490. [Google Scholar] [CrossRef]
- Hill, F.I.; Thompson, K.G.; Grace, N.D. Rickets in alpacas (Lama paces) in New Zealand. N. Z. Vet. J. 1994, 42, 229–232. [Google Scholar] [CrossRef]
- Bosseler, L.; Bakker, J.; Duchateau, L.; Remarque, E.; Langermans, J.A.M.; Cornillie, P.; Chiers, K. 25-OH-vitamin D, parathyroid hormone, and calcium serum levels in captive common marmosets (Callithrix jacchus): Reference values and effect of age, sex, season, and closure of long bone epiphyses. J. Med. Primatol. 2018, 47, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Shinki, T.; Shiinal, Y.; Takahashil, N.; Tanioka, Y.; Koizumi, H.; Sudal, T. Extremely high circulating levels of Lu,25-dihydroxyvitamin D3 in the marmoset, a new world monkey. Biochem. Biophys. Res. Commun. 1983, 114, 452–457. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Kohno, Y.; Yamazaki, T.; Takahashi, N.; Shinki, T.; Horiuchi, N.; Suda, T.; Koizumi, H.; Tanioka, Y.; Yoshikp, S. Bone in the Marmoset: A Resemblance to Vitamin D-Dependent Rickets, Type II. Calcif. Tissue Int. 1986, 39, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Chun, R.F.; Chen, H.; Boldrick, L.; Sweet, C.; Adams, J.S. Cloning, Sequencing, and Functional Characterization of the Vitamin D Receptor in Vitamin D-Resistant New World Primates. Am. J. Primatol. 2001, 54, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Lips, P. Relative Value of 25(OH)D and 1,25(OH)2D Measurements. J. Bone Miner. Res. 2007, 22, 1668–1671. [Google Scholar] [CrossRef]
- Zalewski, A.; Ma, N.S.; Legeza, B.; Renthal, N.; Flück, C.E.; Pandey, A. V Vitamin D-Dependent Rickets Type 1 Caused by Mutations in CYP27B1 Affecting Protein Interactions with Adrenodoxin. J. Clin. Endocrinol. Metab. 2016, 101, 3409–3418. [Google Scholar] [CrossRef] [Green Version]
- Grahn, R.A.; Ellis, M.R.; Grahn, J.C.; Lyons, L.A. A novel CYP27B1 mutation causes a feline vitamin D-dependent rickets type IA. J. Feline Med. Surg. 2012, 14, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Geisen, V.; Weber, K.; Hartmann, K. Vitamin D-Dependent Hereditary Rickets Type I in a Cat. J. Vet. Intern. Med. 2009, 23, 196–199. [Google Scholar] [CrossRef]
- Johnson, K.A.; Church, D.B.; Barton, R.J.; Wood, A.K.W. Vitamin D-dependent rickets in a Saint Bernard dog. J. Small Anim. Pract. 1988, 29, 657–666. [Google Scholar] [CrossRef]
- Teshima, T.; Kurita, S.; Sasaki, T.; Matsumoto, H.; Niina, A.; Abe, D. A genetic variant of CYP2R1 identified in a cat with type 1B vitamin D-dependent rickets: A case report. BMC Vet. Res. 2019, 15, 62. [Google Scholar] [CrossRef]
- Malloy, P.J.; Tasic, V.; Taha, D.; Tütüncüler, F.; Siok, G.; Kah, L.; Wang, J.; Feldman, D. Vitamin D receptor mutations in patients with hereditary 1,25-dihydroxyvitamin D-resistant rickets. Mol. Genet. Metab. 2014, 111, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, D.R.; Anderson, R.M.; Barber, P.J.; Hewison, M. Vitamin D-dependent rickets type II in a cat. J. Small Anim. Pract. 2005, 46, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Tanner, E.; Langley-Hobbs, S.J. Vitamin D-dependent rickets type 2 with characteristic radiographic changes in a 4-month-old kitten. J. Feline Med. Surg. 2005, 7, 307–311. [Google Scholar] [CrossRef] [PubMed]
- LeVine, D.N.; Zhou, Y.; Ghiloni, R.J.; Fields, E.L.; Birkenheuer, A.J.; Gookin, J.L.; Roberston, I.D.; Malloy, P.J.; Feldman, D. Hereditary 1,25-Dihydroxyvitamin D-Resistant Rickets in a Pomeranian Dog Caused by a Novel Mutation in the Vitamin D Receptor Gene. J. Vet. Intern. Med. 2009, 23, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, K.E.; Howe, L.; Thompson, K.G.; Stowell, K.M.; Blair, H.T.; Cockrem, J.F. Normal vitamin D receptor function with increased expression of 25-hydroxyvitamin D 3 -24-hydroxylase in Corriedale sheep with inherited rickets. Res. Vet. Sci. 2011, 91, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.Q.; Ward, L.M.; Liu, S.; Lu, Y.; Xie, Y.; Yuan, B.; Yu, X.; Rauch, F.; Davis, S.I.; Zhang, S.; et al. Loss of DMP1 causes rickets and osteomalacia and identifies a role for osteocytes in mineral metabolism. Nat. Genet. 2006, 38, 1310–1315. [Google Scholar] [CrossRef]
- Lorenz-Depiereux, B.; Bastepe, M.; Benet-Pagès, A.; Amyere, M.; Wagenstaller, J.; Müller-Barth, U.; Badenhoop, K.; Kaiser, S.M.; Rittmaster, R.S.; Shlossberg, A.H.; et al. DMP1 mutations in autosomal recessive hypophosphatemia implicate a bone matrix protein in the regulation of phosphate homeostasis. Nat. Genet. 2006, 38, 1248–1250. [Google Scholar] [CrossRef]
- Bula, C.M.; Huhtakangas, J.; Olivera, C.; Bishop, J.E.; Norman, A.W.; Henry, H.L. Presence of a Truncated Form of the Vitamin D Receptor (VDR) in a Strain of VDR-Knockout Mice. Endocrinology 2005, 146, 5581–5586. [Google Scholar] [CrossRef]
- Mellanby, R.J.; Mellor, P.; Villiers, E.J.; Herrtage, M.E.; Halsall, D.; O’Rahilly, S.; Mcneil, P.E.; Mee, A.P.; Berry, J.L. Hypercalcaemia associated with granulomatous lymphadenitis and elevated 1, 25 dihydroxyvitamin D concentration in a dog. J. Small Anim. Pract. 2006, 47, 207–212. [Google Scholar] [CrossRef]
- Linde, K.J.; Kelleher, T.R.; Perry, J.A. Biological implant-associated granulomatous inflammation resulting in secondary hypercalcemia and azotemia in a dog. Clin. Case Rep. 2018, 6, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Boag, A.K.; Murphy, K.F.; Connolly, D.J. Hypercalcaemia associated with Angiostrongylus vasorum in three dogs. J. Small Anim. Pract. 2005, 46, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Hobi, S.; Bettenay, S.; Majzoub, M.; Mueller, R.; Moser, I. Mycobacterium avium subspecies hominissuis infection in a dog from Germany with multifocal alopecia, exfoliative dermatitis, hypercalcaemia and subsequent sebaceous atrophy. Vet. Rec. Case Rep. 2015, 3, e000168. [Google Scholar] [CrossRef]
- Stern, J.A.; Chew, D.J.; Schissler, J.R.; Green, E.M. Cutaneous and Systemic Blastomycosis, Hypercalcemia, and Excess Synthesis of Calcitriol in a Domestic Shorthair Cat. J. Am. Anim. Hosp. Assoc. 2011, 47, e116–e120. [Google Scholar] [CrossRef] [PubMed]
- Adamany, J.L.; Dhumeaux, M.P. Hypercalcaemia secondary to elevated 1, 25 dihydroxycholecalciferol in a dog with immune-mediated polyarthritis. J. Small Anim. Pract. 2016, 57, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Cortadellas, O.; Fernandez del Palacio, M.J.; Talavera, J.; Bayon, A. Calcium and Phosphorus Homeostasis in Dogs with Spontaneous Chronic Kidney Disease at Different Stages of Severity. J. Vet. Intern. Med. 2010, 24, 73–79. [Google Scholar] [CrossRef]
- De Brito Galvao, J.F.; Nagode, L.A.; Schenck, P.A.; Chew, D.J. Calcitriol, calcidiol, parathyroid hormone, and fibroblast growth factor-23 interactions in chronic kidney disease. J. Vet. Emerg. Crit. Care 2013, 23, 134–162. [Google Scholar] [CrossRef] [PubMed]
- Barber, P.J.; Elliott, J. Feline chronic renal failure: Calcium homeostasis in 80 cases diagnosed between 1992 and 1995. J. Small Anim. Pract. 1998, 39, 108–116. [Google Scholar] [CrossRef]
- Rosol, T.J.; Nagode, L.A.; Couto, C.G.; Hammer, A.S.; Chew, D.J.; Peterson, J.L.; Ayl, R.D.; Steinmeyer, C.L.; Capen, C.C. Parathyroid hormone (PTH)-related protein, PTH and 1,25-dihydroxyvitamin D in dogs with cancer associated hypercalcemia. Endocrinology 1992, 131, 1157–1164. [Google Scholar] [CrossRef]
- Gerber, B.; Hauser, B.; Reusch, C.E. Serum Levels of 25-Hydroxycholecalciferol and 1, 25- Dihydroxycholecalciferol in Dogs with Hypercalcaemia. Vet. Res. Commun. 2004, 28, 669–680. [Google Scholar] [CrossRef]
- Tryfonidou, M.A.; Oosterlaken-Dijksterhuis, M.A.; Mol, J.A.; Van den Ingh, T.S.G.A.M.; van den Brom, W.E.; Hazewinkel, H.A.W. 24-Hydroxylase: Potential key regulator in hypervitaminosis D3 in growing dogs. Am. J. Physiol. Endocrinol. Metab. 2003, 284, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Tryfonidou, M.A.; Holl, M.S.; Vastenburg, M.; Birkenha, D.H.; Van Den Brom, W.E.; Hazewinkel, H.A.W. Hormonal regulation of calcium homeostasis in two breeds of dogs during growth at different rates. J. Anim. Sci. 2003, 81, 1568–1580. [Google Scholar] [CrossRef] [PubMed]
- Young, L.R.; Backus, R.C. Serum 25-hydroxyvitamin D3 and 24R,25-dihydroxyvitamin D3 concentrations in adult dogs are more substantially increased by oral supplementation of 25-hydroxyvitamin D3 than by vitamin D3. J. Nutr. Sci. 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.J.; Zhang, F.; Richards, J.B.; Kestenbaum, B.; Van Meurs, J.B.; Berry, D.; Kiel, D.P.; Streeten, E.A.; Ohlsson, C.; Koller, D.L.; et al. Common genetic determinants of vitamin D insufficiency: A genome-wide association study. Lancet 2010, 376, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Horst, R.L.; Littledike, E.T. Comparison of plasma concentrations of vitamin D and its metabolites in young and aged domestic animals. Comp. Biochem. Physiol. B 1982, 73, 485–489. [Google Scholar] [CrossRef]
- Tang, J.C.Y.; Jackson, S.; Walsh, N.P.; Greeves, J.; Fraser, W.D. The dynamic relationships between the active and catabolic vitamin D metabolites, their ratios, and associations with PTH. Sci. Rep. 2019, 9, 6974. [Google Scholar] [CrossRef]
- Park, H.; Brannon, P.M.; West, A.A.; Yan, J.; Jiang, X.; Perry, C.A.; Malysheva, O.V.; Mehta, S.; Caudill, M.A. Vitamin D Metabolism Varies among Women in Different Reproductive States Consuming the Same Intakes of Vitamin D and Related Nutrients. J. Nutr. 2016, 25, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, C.; Jones, G.; Lyden, E.; Kaufmann, M.; Armas, L.; Anderson-Berry, A. Vitamin D metabolism in the premature newborn: A randomized trial. Clin. Nutr. 2016, 35, 835–841. [Google Scholar] [CrossRef]
- Bouillon, R. Free or Total 25OHD as Marker for Vitamin D Status? J. Bone Miner. Res. 2016, 31, 1124–1127. [Google Scholar] [CrossRef] [Green Version]
- Faix, J.D. Principles and pitfalls of free hormone measurements. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 631–645. [Google Scholar] [CrossRef]
- Bikle, D.D.; Schwartz, J. Vitamin D Binding Protein, Total and Free Vitamin D Levels in Different Physiological and Pathophysiological Conditions. Front. Endocrinol. (Lausanne) 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chun, R.F.; Peercy, B.E.; Orwoll, E.S.; Nielson, C.M.; Adams, J.S.; Hewison, M. Vitamin D and DBP: The free hormone hypothesis revisited. J. Steroid Biochem. Mol. Biol. 2014, 144, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundgren, S.; Carling, T.; Hjälm, G.; Juhlin, C.; Rastad, J.; Pihlgren, U.; Rask, L.; Åkerström, G.; Hellman, P. Tissue Distribution of Human gp330/Megalin, a Putative Ca2+-sensing Protein. J. Histochem. Cytochem. 1997, 45, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safadi, F.F.; Liebhaber, S.A.; Cooke, N.E.; Safadi, F.F.; Thornton, P.; Magiera, H.; Hollis, B.W.; Gentile, M.; Haddad, J.G.; Liebhaber, S.A.; et al. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J. Clin. Invest. 1999, 103, 239–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, R.F.; Nielson, C.M. Chapter 51—Free Vitamin D: Concepts, Assays, Outcomes, and Prospects, 4th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 1, ISBN 9780128099650. [Google Scholar]
- Nielson, C.M.; Jones, K.S.; Chun, R.F.; Jacobs, J.M.; Wang, Y.; Hewison, M.; Adams, J.S.; Swanson, C.M.; Lee, C.G.; Vanderschueren, D.; et al. Free 25-Hydroxyvitamin D: Impact of Vitamin D Binding Protein Assays on Racial-Genotypic Associations. J. Clin. Endocrinol. Metab. 2016, 101, 2226–2234. [Google Scholar] [CrossRef] [Green Version]
- Jassil, N.K.; Sharma, A.; Bikle, D.; Wang, X. Vitamin D binding protein and 25-hydroxyvitamin d levels: Emerging clinical applications. Endocr. Pract. 2017, 23. [Google Scholar] [CrossRef]
- Constans, J.; Gouaillard, C.; Bouissou, C.; Dugoujon, J.M. Polymorphism of the vitamin D binding protein (DBP) among primates: An evolutionary analysis. Am. J. Phys. Anthropol. 1987, 73, 365–377. [Google Scholar] [CrossRef]
- Juneja, R.K.; Gahne, B.; Sandberg, K. Genetic polymorphism of the vitamin D binding protein and another post-albumin protein in horse serum. Anim. Blood Groups Biochem. Genet. 1978, 9, 29–36. [Google Scholar] [CrossRef]
- Bouquet, Y.; Van De Weghe, A.; Van Zeveren, A.; Varewyck, H. Evolutionary conservation of the linkage between the structural loci for serum albumin and vitamin D binding protein (Gc) in cattle. Anim. Genet. 1986, 17, 175–182. [Google Scholar] [CrossRef]
- Tate, M.L.; Dodds, K.G.; Thomas, K.J.; McEwan, K.M. Genetic polymorphism of plasminogen and vitamin D binding protein in red deer (Cervus elaphus L.). Anim. Genet. 1992, 23, 303–313. [Google Scholar] [CrossRef]
- Kaláb, P.; Stratil, A.; Glasnák, V. Genetic polymorphism of serum vitamin D-binding protein (GC) in sheep and mouflon. Anim. Genet. 1990, 21, 317–321. [Google Scholar] [CrossRef]
- Baelen, H.V.A.N.; Allewaert And, K.; Bouillon, R. New Aspects of the Plasma Carrier Protein for 25-Hydroxycholecalciferol in Vertebrates. Ann. N. Y. Acad. Sci. 1988, 538, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Vieth, R.; Kessler, M.J.; Pritzker, K.P.H. Species differences in the binding kinetics of 25-hydroxyvitamin D3 to vitamin D binding protein. Can. J. Physiol. Pharmacol. 1990, 68, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Chun, R.F.; Shieh, A.; Gottlieb, C.; Yacoubian, V.; Wang, J.; Hewison, M.; Adams, J.S. Vitamin D Binding Protein and the Biological Activity of Vitamin D. Front. Endocrinol. 2019, 10, 718. [Google Scholar] [CrossRef]
- Chun, R.F. New perspectives on the vitamin D binding protein. Cell Biochem. Funct. 2012, 30, 445–456. [Google Scholar] [CrossRef]
- Bouillon, R. Chapter 5—The Vitamin D Binding Protein DBP. In Vitamin D; Feldman, D., Pike, J.W., Adams, J.S.B.T.-V.D., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 57–72. ISBN 978-0-12-381978-9. [Google Scholar]
- Bikle, D.; Bouillon, R.; Thadhani, R.; Schoenmakers, I. Vitamin D metabolites in captivity? Should we measure free or total 25 (OH) D to assess vitamin D status? J. Steroid Biochem. Mol. Biol. 2017, 173, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, B.; Cavalier, E.; Bhattoa, H.P.; Pérez-López, F.R.; López-Baena, M.T.; Pérez-Roncero, G.R.; Chedraui, P.; Annweiler, C.; Della Casa, S.; Zelzer, S.; et al. Vitamin D testing: Advantages and limits of the current assays. Eur. J. Clin. Nutr. 2020, 74, 231–247. [Google Scholar] [CrossRef]
- Handelsman, D.J.; Wartofsky, L. Requirement for Mass Spectrometry Sex Steroid Assays in the Journal of Clinical Endocrinology and Metabolism. J. Clin. Endocrinol. Metab. 2013, 98, 3971–3973. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.D.; Berry, J.; Gunter, E.; Jones, G.; Jones, J.; Makin, H.L.J.; Pattni, P.; Sempos, C.T.; Twomey, P.; Williams, E.L.; et al. Hydroxyvitamin D assays: An historical perspective from DEQAS. J. Steroid Biochem. Mol. Biol. 2018, 177, 30–35. [Google Scholar] [CrossRef]
- Kaufmann, M.; Sepiashvili, L.; Singh, R.J. Mass Spectrometry Assays of Vitamin D Metabolites. In Vitamin D: Volume 1: Biochemistry, Physiology and Diagnostics, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 909–923. ISBN 9780128099667. [Google Scholar]
- Garg, U. 25-Hydroxyvitamin D Testing Immunoassays versus Tandem Mass Spectrometry. Clin. Lab. Med. 2018, 38, 439–453. [Google Scholar] [CrossRef]
- van den Ouweland, J.M.W. Analysis of vitamin D metabolites by liquid chromatography-tandem mass spectrometry. Trends Anal. Chem. 2016, 84, 117–130. [Google Scholar] [CrossRef]
- Couchman, L.; Moniz, C.F. Analytical considerations for the biochemical assessment of vitamin D status. Ther. Adv. Musculoskelet. Dis. 2017, 9, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelzer, S.; Goessler, W.; Herrmann, M. Measurement of vitamin D metabolites by mass spectrometry, an analytical challenge. J. Lab. Precis. Med. 2018, 3. [Google Scholar] [CrossRef]
- Volmer, D.A.; Stokes, C.S.; Introduction, I. Analysis of vitamin D metabolic markers by mass spectrometry: Current techniques, limitations of the “ gold standard ” method, and anticipated future directions. Mass Spectrom. Rev. 2015, 34, 2–23. [Google Scholar] [CrossRef]
- Kale, M.S.; Dittmer, K.E.; Roe, W.D.; Gartrell, B.D. Interspecies differences in plasma concentrations of 25-hydroxyvitamin D3 and dermal Vitamin D synthesis of kiwi (Apteryx mantelli), tuatara (Sphenodon punctatus), and New Zealand sea lions (Phocarctos hookeri). J. Comp. Physiol. B 2018, 188, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.J. Quantitation of 25-OH-Vitamin D (25OHD) Using Liquid Tandem Mass Spectrometry (LC-MS-MS) BT—Clinical Applications of Mass Spectrometry: Methods and Protocols. In Clinical Applications of Mass Spectrometry; Garg, U., Hammett-Stabler, C.A., Eds.; Humana Press: Totowa, NJ, USA, 2010; pp. 509–517. ISBN 978-1-60761-459-3. [Google Scholar]
- Singh, R.J.; Taylor, R.L.; Reddy, G.S.; Grebe, S.K.G. C-3 Epimers Can Account for a Significant Proportion of Total Circulating 25-Hydroxyvitamin D in Infants, Complicating Accurate Measurement and Interpretation of Vitamin D Status. J. Clin. Endocrinol. Metab. 2006, 91, 3055–3061. [Google Scholar] [CrossRef] [Green Version]
- Lankes, U.; Elder, P.A.; Lewis, J.G.; George, P. Differential extraction of endogenous and exogenous 25-OH-vitamin D from serum makes the accurate quantification in liquid chromatography-tandem mass spectrometry assays challenging. Ann. Clin. Biochem. 2014, 52, 151–160. [Google Scholar] [CrossRef]
- Maunsell, Z.; Wright, D.J.; Rainbow, S.J. Routine Isotope-Dilution Liquid Chromatography–Tandem Mass Spectrometry Assay for Simultaneous Measurement of the 25-Hydroxy Metabolites of Vitamins D2 and D3. Clin. Chem. 2005, 51, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Schadt, H.S.; Gössl, R.; Seibel, N.; Aebischer, C.-P. Quantification of Vitamin D3 in Feed, Food, and Pharmaceuticals Using High-Performance Liquid Chromatography/Tandem Mass Spectrometry. J. AOAC Int. 2019, 95, 1487–1494. [Google Scholar] [CrossRef]
- Priego Capote, F.; Jiménez, J.R.; Granados, J.M.M.; de Castro, M.D.L. Identification and determination of fat-soluble vitamins and metabolites in human serum by liquid chromatography/triple quadrupole mass spectrometry with multiple reaction monitoring. Rapid Commun. Mass Spectrom. 2007, 21, 1745–1754. [Google Scholar] [CrossRef]
- Zheng, N.; Jiang, H.; Zeng, J. Current advances and strategies towards fully automated sample preparation for regulated LC–MS/MS bioanalysis. Bioanalysis 2014, 6, 2441–2459. [Google Scholar] [CrossRef]
- Strathmann, F.G.; Laha, T.J.; Hoofnagle, A.N. Quantification of 1α,25-Dihydroxy Vitamin D by Immunoextraction and Liquid Chromatography–Tandem Mass Spectrometry. Clin. Chem. 2011, 57, 1279–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivison, F.M.; Hinchliffe, E.; Howarth, N.; Pickersgill, M.; Tetlow, L. Development of a mass spectrometry method for 1,25-dihydroxy vitamin D3 using immunoextraction sample preparation. Ann. Clin. Biochem. 2019, 56, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Dirks, N.F.; Martens, F.; Vanderschueren, D.; Billen, J.; Pauwels, S.; Ackermans, M.T.; Endert, E.; den Heijer, M.; Blankenstein, M.A.; Heijboer, A.C. Determination of human reference values for serum total 1,25-dihydroxyvitamin D using an extensively validated 2D ID-UPLC–MS/MS method. J. Steroid Biochem. Mol. Biol. 2016, 164, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Geib, T.; Schorr, P.; Meier, F.; Volmer, D.A. On the isobaric space of 25-hydroxyvitamin D in human serum: Potential for interferences in liquid chromatography/tandem mass spectrometry, systematic errors and accuracy issues. Rapid Commun. Mass Spectrom. 2015, 29, 1–9. [Google Scholar] [CrossRef]
- Couchman, L.; Benton, C.M.; Moniz, C.F. Variability in the analysis of 25-hydroxyvitamin D by liquid chromatography–tandem mass spectrometry: The devil is in the detail. Clin. Chim. Acta 2012, 413, 1239–1243. [Google Scholar] [CrossRef]
- Adamec, J.; Jannasch, A.; Huang, J.; Hohman, E.; Fleet, J.C.; Peacock, M.; Ferruzzi, M.G.; Martin, B.; Weaver, C.M. Development and optimization of an LC-MS/MS-based method for simultaneous quantification of vitamin D2, vitamin D3, 25-hydroxyvitamin D2 and 25-hydroxyvitamin D3. J. Sep. Sci. 2011, 34, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Binkley, N.; Dawson-Hughes, B.; Durazo-Arvizu, R.; Thamm, M.; Tian, L.; Merkel, J.M.; Jones, J.C.; Carter, G.D.; Sempos, C.T. Vitamin D measurement standardization: The way out of the chaos. J. Steroid Biochem. Mol. Biol. 2017, 173, 117–121. [Google Scholar] [CrossRef]
- Binkley, N.; Krueger, D.C.; Morgan, S.; Wiebe, D. Current status of clinical 25-hydroxyvitamin D measurement: An assessment of between-laboratory agreement. Clin. Chim. Acta 2010, 411, 1976–1982. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.D. Accuracy of 25-Hydroxyvitamin D Assays: Confronting the Issues. Curr. Drug Targets 2011, 12, 19–28. [Google Scholar] [CrossRef]
- Tai, S.S.-C.; Nelson, M.A.; Bedner, M.; Lang, B.E.; Phinney, K.W.; Sander, L.C.; Yen, J.H.; Betz, J.M.; Sempos, C.T.; Wise, S.A. Development of Standard Reference Material (SRM) 2973 Vitamin D Metabolites in Frozen Human Serum (High Level). J. AOAC Int. 2017, 100, 1294–1303. [Google Scholar] [CrossRef]
- Tai, S.S.-C.; Bedner, M.; Phinney, K.W. Development of a Candidate Reference Measurement Procedure for the Determination of 25-Hydroxyvitamin D3 and 25-Hydroxyvitamin D2 in Human Serum Using Isotope-Dilution Liquid Chromatography−Tandem Mass Spectrometry. Anal. Chem. 2010, 82, 1942–1948. [Google Scholar] [CrossRef] [Green Version]
- Stepman, H.C.M.; Vanderroost, A.; Van Uytfanghe, K.; Thienpont, L.M. Candidate Reference Measurement Procedures for Serum 25-Hydroxyvitamin D3 and 25-Hydroxyvitamin D2 by Using Isotope-Dilution Liquid Chromatography–Tandem Mass Spectrometry. Clin. Chem. 2011, 57, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Tai, S.S.-C.; Nelson, M.A. Candidate Reference Measurement Procedure for the Determination of (24R),25-Dihydroxyvitamin D3 in Human Serum Using Isotope-Dilution Liquid Chromatography–Tandem Mass Spectrometry. Anal. Chem. 2015, 87, 7964–7970. [Google Scholar] [CrossRef]
- Burdette, C.Q.; Camara, J.E.; Nalin, F.; Pritchett, J.; Sander, L.C.; Carter, G.D.; Jones, J.; Betz, J.M.; Sempos, C.T.; Wise, S.A. Establishing an Accuracy Basis for the Vitamin D External Quality Assessment Scheme (DEQAS). J. AOAC Int. 2017, 100, 1277–1287. [Google Scholar] [CrossRef]
- Committee for Medicinal Products for Human Use (CHMP). EMA Guideline on Bioanalytical Method Validation; CHMP: London, UK, 2011. [Google Scholar]
- USFDA. Guidance for Industry Bioanalytical Method Validation; U.S. Department of Health and Human Services, Food and Drug Administration: Washington, DC, USA, 2001.
- Bowen, R.A.R.; Remaley, A.T. Interferences from blood collection tube components on clinical chemistry assays. Biochem. Med. 2014, 24, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.W.; Jian, W.; Sullivan, S.; Sankaran, B.; Edom, R.W.; Weng, N.; Sharkey, D. Development and validation of an LC–MS/MS based method for quantification of 25 hydroxyvitamin D2 and 25 hydroxyvitamin D3 in human serum and plasma. J. Chromatogr. B 2014, 961, 62–70. [Google Scholar] [CrossRef]
- Ding, S.; Schoenmakers, I.; Jones, K.; Koulman, A.; Prentice, A.; Volmer, D.A. Quantitative determination of vitamin D metabolites in plasma using UHPLC-MS/MS. Anal. Bioanal. Chem. 2010, 398, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Tørnqvist-Johnsen, C.; Schnabel, T.; Gow, A.G.; Del-Pozo, J.; Milne, E.; Macklin, V.; Fisher, C.; Handel, I.; Mellanby, R.J. Investigation of the relationship between ionised and total calcium in dogs with ionised hypercalcaemia. J. Small Anim. Pract. 2020, 61, 247–252. [Google Scholar] [CrossRef]
- Lawrence, S.H.; Melnick, P.J.; Weimer, H.E. A Species Comparison of Serum Proteins and Enzymes by Starch Gel Electrophoresis. Proc. Soc. Exp. Biol. Med. 1960, 105, 572–575. [Google Scholar] [CrossRef]
- Kosa, T.; Maruyama, T.; Otagiri, M. Species Differences of Serum Albumins: II. Chemical and Thermal Stability. Pharm. Res. 1998, 15, 449–454. [Google Scholar] [CrossRef]
- Nagy, O.; Tóthová, C.; Nagyová, V.; Kováč, G. Comparison of serum protein electrophoretic pattern in cows and small ruminants. Acta Vet. Brun. 2015, 84, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Piccoli, S.P.; Sauer, J.M. Points to Consider Document: Scientific and Regulatory Considerations for the Analytical Validation of Assays Used in the Qualification of Biomarkers in Biological Matrices; Critical Path Institute: Tucson, AZ, USA, 2019; pp. 1–79. [Google Scholar]
- Subramaniam, S. Bioanalysis. AAPS Adv. Pharm. Sci. Ser. 2014, 13, 419–458. [Google Scholar] [CrossRef]
- Briggs, R.J.; Nicholson, R.; Vazvaei, F.; Busch, J.; Mabuchi, M.; Mahesh, K.S.; Brudny-Kloeppel, M.; Weng, N.; Galvinas, P.A.R.; Duchene, P.; et al. Method Transfer, Partial Validation, and Cross Validation: Recommendations for Best Practices and Harmonization from the Global Bioanalysis Consortium Harmonization Team. AAPS J. 2014, 16, 1143–1148. [Google Scholar] [CrossRef] [Green Version]
- Annesley, T.M. Ion Suppression in Mass Spectrometry. Clin. Chem. 2003, 49, 1041–1044. [Google Scholar] [CrossRef] [Green Version]
- Trufelli, H.; Palma, P.; Famiglini, G.; Cappiello, A. An overview of matrix effects in liquid chromatography–mass spectrometry. Mass Spectrom. Rev. 2011, 30, 491–509. [Google Scholar] [CrossRef]
- Van Eeckhaut, A.; Lanckmans, K.; Sarre, S.; Smolders, I.; Michotte, Y. Validation of bioanalytical LC–MS/MS assays: Evaluation of matrix effects. J. Chromatogr. B 2009, 877, 2198–2207. [Google Scholar] [CrossRef]
- Matuszewski, B.K.; Constanzer, M.L.; Chavez-Eng, C.M. Strategies for the Assessment of Matrix Effect in Quantitative Bioanalytical Methods Based on HPLC−MS/MS. Anal. Chem. 2003, 75, 3019–3030. [Google Scholar] [CrossRef]
- Panuwet, P.; Hunter, R.E.; D’Souza, P.E.; Chen, X.; Radford, S.A.; Cohen, J.R.; Marder, M.E.; Kartavenka, K.; Ryan, P.B.; Barr, D.B. Biological Matrix Effects in Quantitative Tandem Mass Spectrometry-Based Analytical Methods: Advancing Biomonitoring. Crit. Rev. Anal. Chem. 2016, 46, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Yang, S.; Wang, P.G. Matrix effects and application of matrix effect factor. Bioanalysis 2017, 9, 1839–1844. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, C.T.; Bansal, S.; Booth, B.; DeStefano, A.J.; Rose, M.J.; Sailstad, J.; Shah, V.P.; Skelly, J.P.; Swann, P.G.; Weiner, R. Workshop/conference report—Quantitative bioanalytical methods validation and implementation: Best practices for chromatographic and ligand binding assays. AAPS J. 2007, 9, E30–E42. [Google Scholar] [CrossRef] [Green Version]
- Hewavitharana, A.K. Matrix matching in liquid chromatography–mass spectrometry with stable isotope labelled internal standards—Is it necessary? J. Chromatogr. A 2011, 1218, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Kersey, D.C.; Dehnhard, M. The use of noninvasive and minimally invasive methods in endocrinology for threatened mammalian species conservation. Gen. Comp. Endocrinol. 2014, 203, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Ouschan, C.; Kuchar, A.; Möstl, E. Measurement of cortisol in dog hair: A noninvasive tool for the diagnosis of hypercortisolism. Vet. Dermatol. 2013, 24, 428-e94. [Google Scholar] [CrossRef] [PubMed]
- Carlow, D.C.; Schofield, R.C.; Denburg, M. Quantitation of 25-OH-Vitamin-D2 and 25-OH-Vitamin-D3 in Urine Using LC-MS/MS. In Clinical Applications of Mass Spectrometry in Biomolecular Analysis. Methods in Molecular Biology, Vol 1378; Garg, U., Ed.; Humana Press: New York, NY, USA, 2016. [Google Scholar]
- Higashi, T.; Homma, S.; Iwata, H.; Shimada, K. Characterization of urinary metabolites of vitamin D3 in man under physiological conditions using liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2002, 29, 947–955. [Google Scholar] [CrossRef]
- Ogawa, S.; Ooki, S.; Shinoda, K.; Higashi, T. Analysis of urinary vitamin D3 metabolites by liquid chromatography/tandem mass spectrometry with ESI-enhancing and stable isotope-coded derivatization. Anal. Bioanal. Chem. 2014, 406, 6647–6654. [Google Scholar] [CrossRef]
- Clarke, M.W.; Black, L.J.; Hart, P.H.; Jones, A.P.; Palmer, D.J. The challenges of developing and optimising an assay to measure 25-hydroxyvitamin D in saliva. J. Steroid Biochem. Mol. Biol. 2019, 194, 105437. [Google Scholar] [CrossRef]
- Higashi, T.; Shibayama, Y.; Fuji, M.; Shimada, K. Liquid chromatography–tandem mass spectrometric method for the determination of salivary 25-hydroxyvitamin D3: A noninvasive tool for the assessment of vitamin D status. Anal. Bioanal. Chem. 2008, 391, 229–238. [Google Scholar] [CrossRef]
- Higashi, T.; Hijikuro, M.; Yamagata, K.; Ogawa, S. Overestimation of salivary 25-hydroxyvitamin D3 level when using stimulated saliva with gum-chewing. Steroids 2013, 78, 884–887. [Google Scholar] [CrossRef]
- Zgaga, L.; Laird, E. 25-Hydroxyvitamin D Measurement in Human Hair: Results from a Proof-of-Concept study. Nutrients 2019, 11, 423. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Citation | Species | Metabolites Measured | LOQ | m/z Transitions | Sample Type/Vol | Sample Prep Method | Derivatization | LC Column | Ionization Source | LC-MS System |
|---|---|---|---|---|---|---|---|---|---|---|
| Hurst (2020) [80] | Dog | 25(OH)D2 25(OH)D3 | 0.5 nmol/L 4.0 nmol/L | 758.4 -> 740.2 747.3 -> 729.3 | Serum/200 μL | SLE | Yes—DMEQ-TAD | Raptor FP (2.7 μm, 100 × 2.1 mm) | ESI | Shimadzu Nexera X2 UHPLC—Sciex QTrap 6500+ |
| Hurst (2020) [81] | Dog | 25(OH)D2 C3-epi-25(OH)D2 25(OH)D3 C3-epi-25(OH)D3 | 7.8 nmol/L 7.8 nmol/L 4.0 nmol/L 4.0 nmol/L | 413.3 -> 395.3 413.3 -> 395.3 401.3 -> 383.3 401.3 -> 383.3 | Serum/200 μL | SLE | No | Raptor FP (2.7 μm, 100 × 2.1 mm) | ESI | Shimadzu Nexera X2 UHPLC—Sciex QTrap 6500+ |
| Groth (2019) b [82] | Dog | 25(OH)D3 24,25(OH)2D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Agilent 1290 HPLC—Agilent 6460 triple quadrupole |
| Mick (2019) b [83] | Dog | 25(OH)D3 24,25(OH)2D3 1,25(OH)2D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Agilent 1290 HPLC—Agilent 6460 triple quadrupole |
| Fritz (2017) [84] | Rat Dog Mouse Monkey | 25(OH)D3 25(OH)D2 | 5.0 nmol/L | 383.3 -> 229.2 395.3 -> 269.3 | Serum/50 μL | SLE | No | Phenomenex Luna C8 (3 μm, 5 0 × 2.0 mm) | APCI | Agilent HPLC—Sciex 4000 QTrap |
| Sprinkle (2017) d [85] | Cat Dog Rat | 25(OH)D3 C3-epi-25(OH)D3 | Not specified | 401.4 -> 383.3 401.4 -> 383.3 | Serum/200 μL | PPT + online extraction | No | Chirex-PGLY and DNB (250 × 4.6 mm) | APCI | TX4 Turbo Flow—Sciex API 4000 |
| Weidner (2017) b [28] | Dog | 24,25(OH)2D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Agilent 1290 HPLC—Agilent 6460 triple quadrupole |
| Willcox (2016) c [86] | Dog | 25(OH)D2 25(OH)D3 | 12.5 nmol/L | 413 -> 395 401 -> 383 | Serum/100 μL | SPE | No | Not specified | Not specified | Not specified |
| Spoo (2015) a [87] | Dog | 25(OH)D3 24,25(OH)2D3 | Not specified | Not specified | Serum/plasma 100 μL for 25(OH)D3 200 μL for 24,25(OH)2D3 | PPT + LLE | No | Not specified | Not specified | Agilent 1290 HPLC—Agilent 6460 triple quadrupole |
| Azarpeykan (2016) e [88,126] | Horse | 25(OH)D2 25(OH)D3 | <4 nmol/L | 413.3 -> 355.4 401.3 -> 383.4 | Serum/60 μL | PPT + LLE | No | ACE C8 (3 μm, 50 × 2.1 mm) | ESI | Shimadzu HPLC—Sciex 4000 |
| Allott (2020) e [89] | Sheep | 25(OH)D3 25(OH)D2 | Not specified | Not specified | Plasma/vol not specified | Not specified | Not specified | Not specified | Not specified | Not specified |
| Dittmer (2020) e [90] | Sheep | 25(OH)D3 25(OH)D2 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Not specified |
| Zhou (2019) [91] | Sheep | 25(OH)D2 25(OH)D3 | 7.2 nmol/L 5.6 nmol/L | 758.5 -> 740.0 746.5 -> 728.4 | Serum/100 μL | PPT + SPE | Yes—DMEQ-TAD | ACE UltraCore 2.5 SuperC18 (2.5 μm, 75 × 2.1 mm) | ESI | Ultimate 3000 HPLC—Bruker amaZon ETD |
| Handel (2016) f [30] | Sheep | 25(OH)D2 25(OH)D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | LC system not specified—Sciex QTrap 5500 |
| Dittmer (2011) e [202] | Sheep | 25(OH)D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Not specified |
| Celi (2018) h [92] | Cattle | Vitamin D2 Vitamin D3 25(OH)D3 | Not specified | Not specified | Serum tissues (fat, muscle, kidney, liver) Feedstuff | Not specified | Not specified | Not specified | Not specified | Not specified |
| Guo (2018) g [93] | Cattle | Vitamin D3 25(OH)D3 | Not specified | Not specified | Plasma/100 μL Milk/3 g for vitamin D3 and 8 g for 25(OH)D3 | PPT + LLE for plasma Saponification + LLE for milk vitamin D3 Saponification + LLE + SPE for milk 25(OH)D3 | No | Not specified | APCI | Agilent 1290 HPLC—Sciex 4000 |
| Alexander (2017) b [94] | Pig | Vitamin D3 | Not specified | Not specified | Serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Agilent 1290 HPLC—Agilent 6460 triple quadrupole |
| Flohr (2016) i [157,158] | Pig | Vitamin D3 25(OH)D3 Vitamin D3 (feed) | 2.5 nmol/L 12.5 nmol/L 1.6 μg/kg | Not specified 383 -> 159 Not specified | Serum/vol not specified Foodstuff/amount not specified | LLE for serum Saponification + LLE for foodstuff | No | Serum—Zorbax Eclipse XDB-C18 (5 μm, 150 × 4.6mm) Foodstuff—2D LC with Ascentis Express octylsilyl (C8) (2.7 μm, 150 × 3 mm) and Ascentis Express C18 (2.7 μm, 150 × 3mm) | Serum—ESI Foodstuff—ESI (Turbo Ion Spray) | Serum—Agilent 1200 HPLC—Agilent 6410 triple quadrupole Foodstuff—Agilent 1200 HPLC—Sciex API 4000 |
| Kuhn (2015 + 2014 + 2019) [95,175,176] | Poultry | 25(OH)D3 Vitamin D3 25(OH)D3 | 3.7 nmol/L 0.17 μg/100g 0.1 μg/100g | 576 -> 298 560 -> 298 | Plasma/vol not specified Egg yolk + skin | LLE Saponification + LLE + HPLC | Yes—PTAD | Hypersil ODS (5 μm, 100 × 2 mm) | ESI | Agilent 1100—API 2000 (Applied Biosystems) |
| Browning (2014) j [172] | Poultry | Vitamin D3 25(OH)D3 | 0.1 μg/kg | Not specified | Egg yolk/7.5 g | Saponification + SPE | Not specified | Not specified | Not specified | Not specified |
| Schutkowski (2013) [96] | Poultry | 25(OH)D3 Vitamin D3 25(OH)D3 7-DHC | 3.7 nmol/L 0.17 μg/100g 0.1 μg/100g Not specified | Not specified | Plasma/vol not specified Egg yolk + meat/vol or weight not specified | LLE Homogenization and LLE | Yes—PTAD | Hypersil ODS (5 μm, 100 × 2 mm) | Not specified | Agilent 1100—API 2000 (Applied Biosystems) |
| Ziegler (2018) [98] | Baboon | Vitamin D3 25(OH)D2 25(OH)D3 24,25(OH)2D3 | Not specified 2.19 nmol/L 2.52 nmol/L 1.15 nmol/L | Not specified 413.3 -> 355.3 401.3 -> 159.0 417.3 -> 121.0 | Serum/100 μL | PPT + SPE | Yes—PTAD | Phenomenex Kinetex C18 (2.6 μm, 100 × 2.1 mm) | ESI | Shimadzu Prominence HPLC—Sciex QTrap 5500 |
| Ziegler (2015) [97] | Marmosets Rhesus macaques | 25(OH)D2 25(OH)D3 1,25(OH)2D3 | 1.3 nmol/L 37.5 pmol/L | Not specified 748.6 -> 689.5 | Serum/50 μL for rhesus macaques and 10 μL for marmosets Serum/200 μL | PPT Dual column SPE | No Yes—Amplifex | Phenomenex Luna C8 (3 μm, 50 × 2 mm) Phenomenex Kinetex C18 (2.4 μm, 150 × 3 mm) | APCI ESI (Turbo Spray Ion) | Shimadzu Prominence HPLC—Sciex QTrap 5500 Shimadzu Prominence HPLC—Sciex QTrap 5500 |
| Kale (2018) e [254] | Brown kiwi, Tuatara, New Zealand Sea Lion | 25(OH)D2 25(OH)D3 | Not specified | Not specified | Plasma + serum/vol not specified | Not specified | Not specified | Not specified | Not specified | Not specified |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurst, E.A.; Homer, N.Z.; Mellanby, R.J. Vitamin D Metabolism and Profiling in Veterinary Species. Metabolites 2020, 10, 371. https://doi.org/10.3390/metabo10090371
Hurst EA, Homer NZ, Mellanby RJ. Vitamin D Metabolism and Profiling in Veterinary Species. Metabolites. 2020; 10(9):371. https://doi.org/10.3390/metabo10090371
Chicago/Turabian StyleHurst, Emma A., Natalie Z. Homer, and Richard J. Mellanby. 2020. "Vitamin D Metabolism and Profiling in Veterinary Species" Metabolites 10, no. 9: 371. https://doi.org/10.3390/metabo10090371
APA StyleHurst, E. A., Homer, N. Z., & Mellanby, R. J. (2020). Vitamin D Metabolism and Profiling in Veterinary Species. Metabolites, 10(9), 371. https://doi.org/10.3390/metabo10090371