
The Role of Citrate Transporter INDY in Metabolism and Stem Cell Homeostasis


Abstract
:1. Introduction
2. Indy Reduction Extends Longevity of Flies and Worms by Mimicking Calorie Restriction
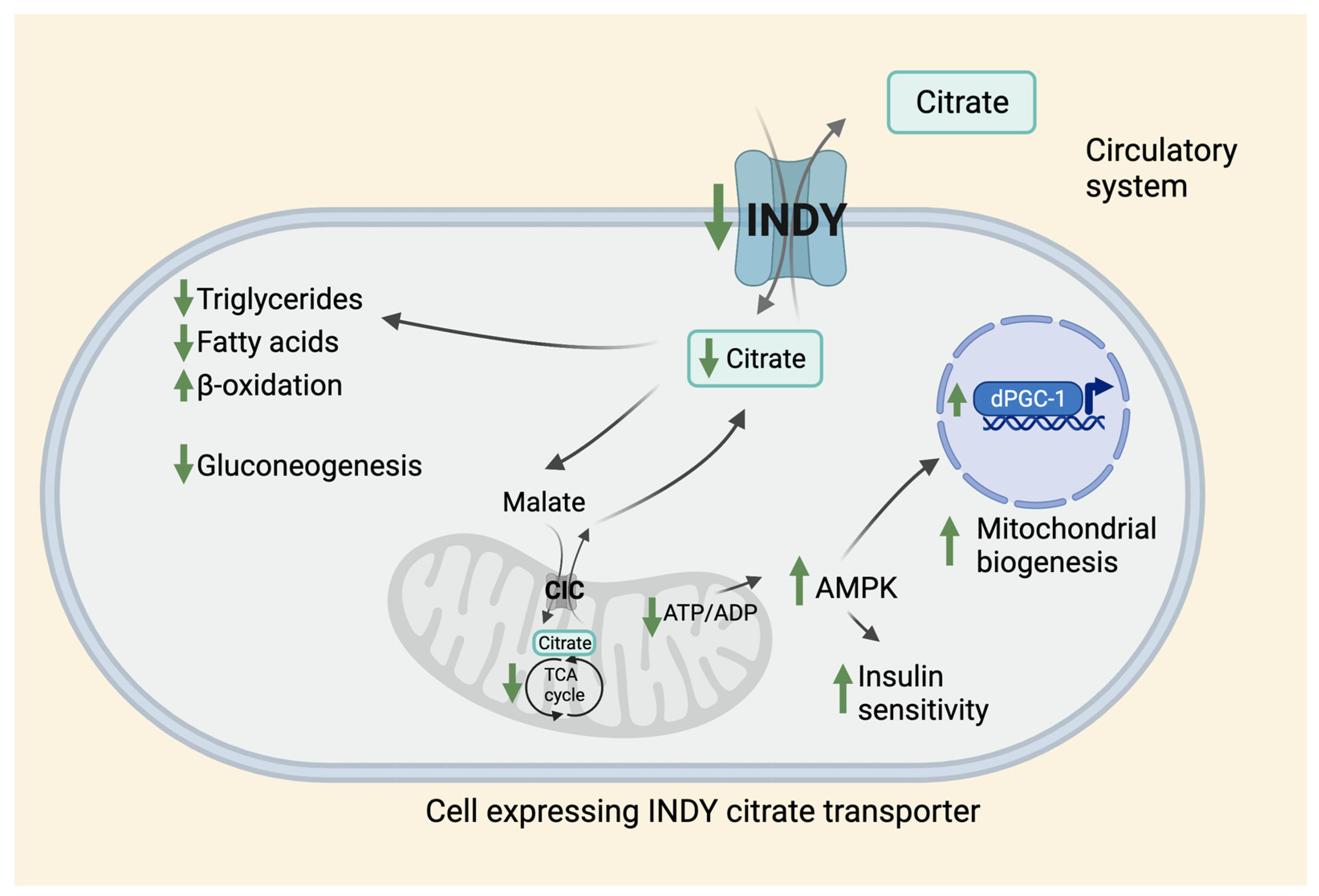
3. The Role of INDY in Metabolism
4. Indy Transcriptional Regulation
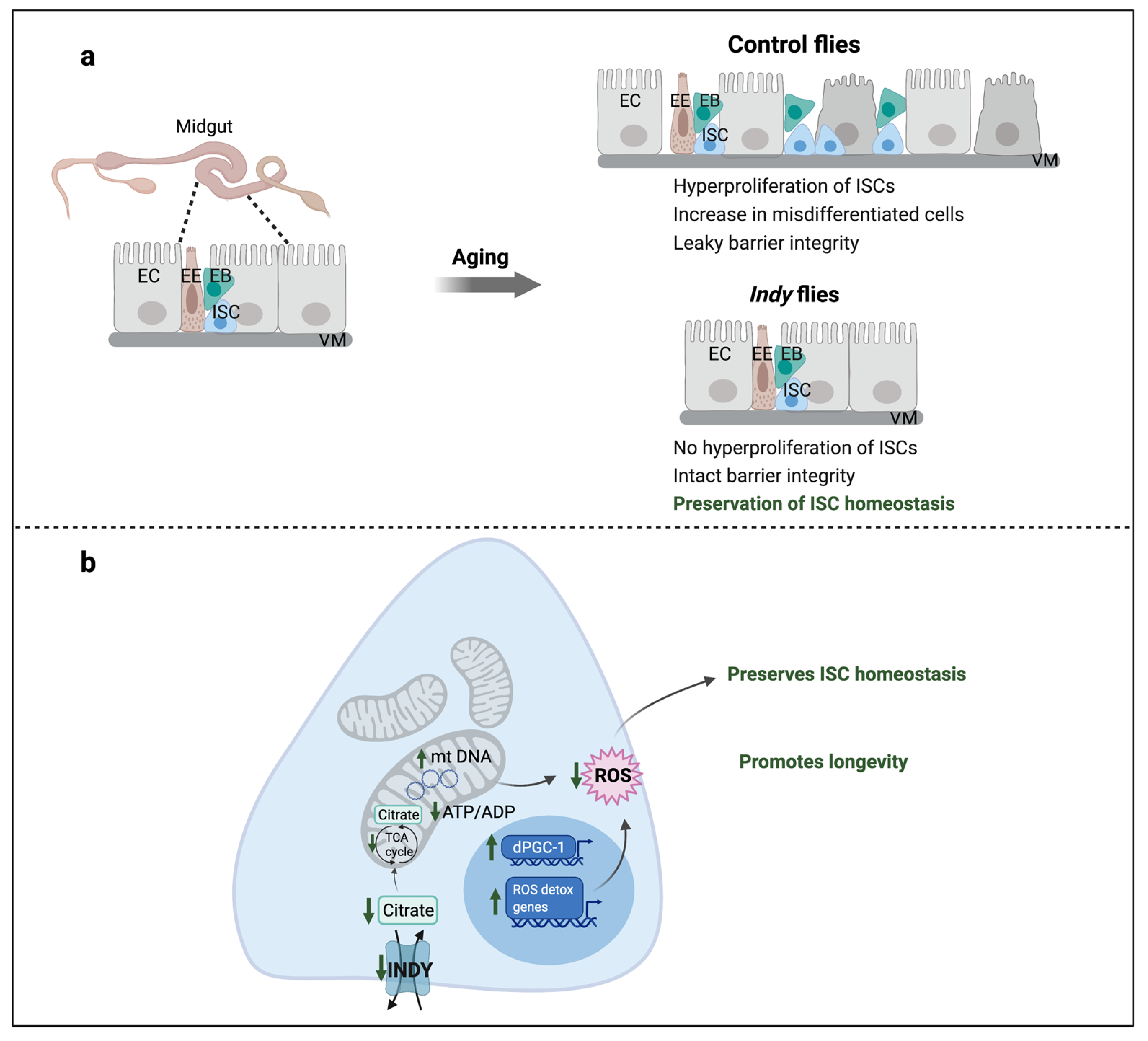
5. Effects of Reduced Indy on Intestinal Stem Cell Homeostasis
6. Effects of Indy Reduction on Spermatogenesis
7. Conclusions and Future Directions
- INDY as a potential therapeutic target
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rogina, B.; Reenan, R.A.; Nilsen, S.P.; Helfand, S.L. Extended Life-Span Conferred by Cotransporter Gene Mutations in Drosophila. Science 2000, 290, 2137–2140. [Google Scholar] [CrossRef]
- Knauf, F.; Mohebbi, N.; Teichert, C.; Herold, D.; Rogina, B.; Helfand, S.; Gollasch, M.; Luft, F.C.; Aronson, P.S. The Life-Extending Gene Indy Encodes an Exchanger for Krebs-Cycle Intermediates. Biochem. J. 2006, 397, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Knauf, F.; Rogina, B.; Jiang, Z.; Aronson, P.S.; Helfand, S.L. Functional Characterization and Immunolocalization of the Transporter Encoded by the Life-Extending Gene Indy. Proc. Natl. Acad. Sci. USA 2002, 99, 14315–14319. [Google Scholar] [CrossRef] [Green Version]
- Gopal, E.; Babu, E.; Ramachandran, S.; Bhutia, Y.D.; Prasad, P.D.; Ganapathy, V. Species-Specific Influence of Lithium on the Activity of SLC13A5 (NaCT): Lithium-Induced Activation Is Specific for the Transporter in Primates. J. Pharmacol. Exp. Ther. 2015, 353, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.Y.; Jiang, J.; Ma, H.M. Molecular Characterization, Tissue Expression Profile, and SNP Analysis of Porcine SLC13A5. Genet. Mol. Res. 2015, 14, 16090–16101. [Google Scholar] [CrossRef]
- Mancusso, R.; Gregorio, G.G.; Liu, Q.; Wang, D.-N. Structure and Mechanism of a Bacterial Sodium-Dependent Dicarboxylate Transporter. Nature 2012, 491, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Fei, Y.-J.; Zhuang, L.; Gopal, E.; Miyauchi, S.; Ganapathy, V. Functional Features and Genomic Organization of Mouse NaCT, a Sodium-Coupled Transporter for Tricarboxylic Acid Cycle Intermediates. Biochem. J. 2004, 378, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Fei, Y.-J.; Huang, W.; Zhuang, L.; Chen, Z.; Ganapathy, V. Functional Identity of Drosophila melanogaster Indy as a Cation-Independent, Electroneutral Transporter for Tricarboxylic Acid-Cycle Intermediates. Biochem. J. 2002, 367, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo-Martinez, V.; Urbatsch, I.L.; Ganapathy, V. Functional Distinction between Human and Mouse Sodium-Coupled Citrate Transporters and Its Biologic Significance: An Attempt for Structural Basis Using a Homology Modeling Approach. Chem. Rev. 2021, 121, 5359–5377. [Google Scholar] [CrossRef]
- Willmes, D.M.; Kurzbach, A.; Henke, C.; Schumann, T.; Zahn, G.; Heifetz, A.; Jordan, J.; Helfand, S.L.; Birkenfeld, A.L. The Longevity Gene INDY (I’m Not Dead Yet) in Metabolic Control: Potential as Pharmacological Target. Pharmacol. Ther. 2018, 185, 1–11. [Google Scholar] [CrossRef]
- Kopel, J.J.; Bhutia, Y.D.; Sivaprakasam, S.; Ganapathy, V. Consequences of NaCT/SLC13A5/mINDY Deficiency: Good versus Evil, Separated Only by the Blood–Brain Barrier. Biochem. J. 2021, 478, 463–486. [Google Scholar] [CrossRef] [PubMed]
- Mishra, D.; Kannan, K.; Meadows, K.; Macro, J.; Li, M.; Frankel, S.; Rogina, B. INDY—From Flies to Worms, Mice, Rats, Non-Human Primates, and Humans. Front. Genet. 2021, 8, 66. [Google Scholar]
- Rogina, B.; Helfand, S.L. INDY Mutations and Drosophila longevity. Front. Genet. 2013, 4, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, Y.-J.; Liu, J.-C.; Inoue, K.; Zhuang, L.; Miyake, K.; Miyauchi, S.; Ganapathy, V. Relevance of NAC-2, an Na+-Coupled Citrate Transporter, to Life Span, Body Size and Fat Content in Caenorhabditis Elegans. Biochem. J. 2004, 379, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.-J.; Inoue, K.; Ganapathy, V. Structural and Functional Characteristics of Two Sodium-Coupled Dicarboxylate Transporters (CeNaDC1 and CeNaDC2) from Caenorhabditis Elegans and Their Relevance to Life Span. J. Biol. Chem. 2003, 278, 6136–6144. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.P.; Rogina, B. Increased Mitochondrial Biogenesis Preserves Intestinal Stem Cell Homeostasis and Contributes to Longevity in Indy Mutant Flies. Aging (Albany NY) 2014, 6, 335–350. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, F.; Karadeniz, Z.; Fischer-Rosinsky, A.; Willmes, D.M.; Spranger, J.; Birkenfeld, A.L. Knockdown of Indy/CeNac2 Extends Caenorhabditis Elegans Life Span by Inducing AMPK/Aak-2. Aging 2015, 7, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Birkenfeld, A.L.; Lee, H.-Y.; Guebre-Egziabher, F.; Alves, T.C.; Jurczak, M.J.; Jornayvaz, F.R.; Zhang, D.; Hsiao, J.J.; Martin-Montalvo, A.; Fischer-Rosinsky, A.; et al. Deletion of the Mammalian INDY Homolog Mimics Aspects of Dietary Restriction and Protects against Adiposity and Insulin Resistance in Mice. Cell Metab. 2011, 14, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Bhutia, Y.D.; Kopel, J.J.; Lawrence, J.J.; Neugebauer, V.; Ganapathy, V. Plasma Membrane Na+-Coupled Citrate Transporter (SLC13A5) and Neonatal Epileptic Encephalopathy. Molecules 2017, 22, E378. [Google Scholar] [CrossRef]
- Inoue, K.; Zhuang, L.; Maddox, D.M.; Smith, S.B.; Ganapathy, V. Structure, Function, and Expression Pattern of a Novel Sodium-Coupled Citrate Transporter (NaCT) Cloned from Mammalian Brain. J. Biol. Chem. 2002, 277, 39469–39476. [Google Scholar] [CrossRef] [Green Version]
- Costello, L.C.; Franklin, R.B. Plasma Citrate Homeostasis: How It Is Regulated; And Its Physiological and Clinical Implications. An Important, But Neglected, Relationship in Medicine. HSOA J. Hum. Endocrinol. 2016, 1, 005. [Google Scholar] [CrossRef]
- Wang, P.-Y.; Neretti, N.; Whitaker, R.; Hosier, S.; Chang, C.; Lu, D.; Rogina, B.; Helfand, S.L. Long-Lived Indy and Calorie Restriction Interact to Extend Life Span. Proc. Natl. Acad. Sci. USA. 2009, 106, 9262–9267. [Google Scholar] [CrossRef] [Green Version]
- Marden, J.H.; Rogina, B.; Montooth, K.L.; Helfand, S.L. Conditional Tradeoffs between Aging and Organismal Performance of INDY Long-Lived Mutant Flies. Proc. Natl. Acad. Sci. USA. 2003, 100, 3369–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.-T.; Chang, C.; Reenan, R.A.; Helfand, S.L. Indy Gene Variation in Natural Populations Confers Fitness Advantage and Life Span Extension through Transposon Insertion. Aging (Albany NY) 2014, 6, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogina, B.; Benzer, S.; Helfand, S.L. Drosophila Drop-Dead Mutations Accelerate the Time Course of Age-Related Markers. Proc. Natl. Acad. Sci. USA 1997, 94, 6303–6306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luckinbill, L.S.; Clare, M.J. Selection for Life Span in Drosophila melanogaster. Heredity 1985, 55, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, J.M.; Walker, G.A.; Martinez-Diaz, P.; Bjedov, I.; Driege, Y.; Jacobs, H.T.; Gems, D.; Partridge, L. No Influence of Indy on Lifespan in Drosophila after Correction for Genetic and Cytoplasmic Background Effects. PLoS Genet. 2007, 3, e95. [Google Scholar] [CrossRef] [Green Version]
- Evangelou, A.; Ignatiou, A.; Antoniou, C.; Kalanidou, S.; Chatzimatthaiou, S.; Shianiou, G.; Ellina, S.; Athanasiou, R.; Panagi, M.; Apidianakis, Y.; et al. Unpredictable Effects of the Genetic Background of Transgenic Lines in Physiological Quantitative Traits. G3 (Bethesda) 2019, 9, 3877–3890. [Google Scholar] [CrossRef] [Green Version]
- Tamamouna, V.; Panagi, M.; Theophanous, A.; Demosthenous, M.; Michail, M.; Papadopoulou, M.; Teloni, S.; Pitsouli, C.; Apidianakis, Y. Evidence of Two Types of Balance between Stem Cell Mitosis and Enterocyte Nucleus Growth in the Drosophila Midgut. Development 2020, 147, dev189472. [Google Scholar] [CrossRef] [PubMed]
- Bross, T.G.; Rogina, B.; Helfand, S.L. Behavioral, Physical, and Demographic Changes in Drosophila Populations through Dietary Restriction. Aging Cell 2005, 4, 309–317. [Google Scholar] [CrossRef]
- Parashar, V.; Rogina, B. dSir2 Mediates the Increased Spontaneous Physical Activity in Flies on Calorie Restriction. Aging 2009, 1, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Neretti, N.; Wang, P.-Y.; Brodsky, A.S.; Nyguyen, H.H.; White, K.P.; Rogina, B.; Helfand, S.L. Long-Lived Indy Induces Reduced Mitochondrial Reactive Oxygen Species Production and Oxidative Damage. Proc. Natl. Acad. Sci. USA 2009, 106, 2277–2282. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. Elegans Mutant That Lives Twice as Long as Wild Type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of Life-Span by Loss of CHICO, a Drosophila Insulin Receptor Substrate Protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A Mutant Drosophila Insulin Receptor Homolog That Extends Life-Span and Impairs Neuroendocrine Function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, K.; Fridell, Y.-W.C. Functional Implications of Drosophila Insulin-like Peptides in Metabolism, Aging, and Dietary Restriction. Front. Physiol. 2013, 4, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nässel, D.R.; Liu, Y.; Luo, J. Insulin/IGF Signaling and Its Regulation in Drosophila. Gen. Comp. Endocrinol. 2015, 221, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Gross, D.N.; van den Heuvel, A.P.J.; Birnbaum, M.J. The Role of FoxO in the Regulation of Metabolism. Oncogene 2008, 27, 2320–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesta, D.H.; Perry, R.J.; Guebre-Egziabher, F.; Zhang, D.; Jurczak, M.; Fischer-Rosinsky, A.; Daniels, M.A.; Willmes, D.M.; Bhanot, S.; Bornstein, S.R.; et al. Prevention of Diet-Induced Hepatic Steatosis and Hepatic Insulin Resistance by Second Generation Antisense Oligonucleotides Targeted to the Longevity Gene mINDY (Slc13a5). Aging (Albany NY) 2015, 7, 1086–1093. [Google Scholar] [CrossRef] [Green Version]
- Brachs, S.; Winkel, A.F.; Tang, H.; Birkenfeld, A.L.; Brunner, B.; Jahn-Hofmann, K.; Margerie, D.; Ruetten, H.; Schmoll, D.; Spranger, J. Inhibition of Citrate Cotransporter Slc13a5/mINDY by RNAi Improves Hepatic Insulin Sensitivity and Prevents Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. Mol. Metab. 2016, 5, 1072–1082. [Google Scholar] [CrossRef]
- Gopal, E.; Miyauchi, S.; Martin, P.M.; Ananth, S.; Srinivas, S.R.; Smith, S.B.; Prasad, P.D.; Ganapathy, V. Expression and Functional Features of NaCT, a Sodium-Coupled Citrate Transporter, in Human and Rat Livers and Cell Lines. Am. J. Physiol. Gastrointest Liver Physiol. 2007, 292, G402–G408. [Google Scholar] [CrossRef]
- Ruderman, N.B.; Saha, A.K.; Vavvas, D.; Witters, L.A. Malonyl-CoA, Fuel Sensing, and Insulin Resistance. Am. J. Physiol. 1999, 276, E1–E18. [Google Scholar] [CrossRef]
- Rogers, R.P.; Rogina, B. The Role of INDY in Metabolism, Health and Longevity. Front. Genet. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Rogina, B. INDY—A New Link to Metabolic Regulation in Animals and Humans. Front. Genet. 2017, 8, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferramosca, A.; Zara, V. Dietary Fat and Hepatic Lipogenesis: Mitochondrial Citrate Carrier as a Sensor of Metabolic Changes. Adv. Nutr. 2014, 5, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Aluvila, S.; Kotaria, R.; Mayor, J.A.; Walters, D.E.; Kaplan, R.S. Mitochondrial and Plasma Membrane Citrate Transporters: Discovery of Selective Inhibitors and Application to Structure/Function Analysis. Mol. Cell Pharmacol. 2010, 2, 101–110. [Google Scholar]
- Zhang, J.; Nuebel, E.; Daley, G.Q.; Koehler, C.M.; Teitell, M.A. Metabolic Regulation in Pluripotent Stem Cells during Reprogramming and Self-Renewal. Cell Stem Cell 2012, 11, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajor, A.M.; Gangula, R.; Yao, X. Cloning and Functional Characterization of a High-Affinity Na(+)/Dicarboxylate Cotransporter from Mouse Brain. Am. J. Physiol. Cell Physiol. 2001, 280, C1215–C1223. [Google Scholar] [CrossRef] [Green Version]
- Thevenon, J.; Milh, M.; Feillet, F.; St-Onge, J.; Duffourd, Y.; Jugé, C.; Roubertie, A.; Héron, D.; Mignot, C.; Raffo, E.; et al. Mutations in SLC13A5 Cause Autosomal-Recessive Epileptic Encephalopathy with Seizure Onset in the First Days of Life. Am. J. Hum. Genet. 2014, 95, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Hardies, K.; de Kovel, C.G.F.; Weckhuysen, S.; Asselbergh, B.; Geuens, T.; Deconinck, T.; Azmi, A.; May, P.; Brilstra, E.; Becker, F.; et al. Recessive Mutations in SLC13A5 Result in a Loss of Citrate Transport and Cause Neonatal Epilepsy, Developmental Delay and Teeth Hypoplasia. Brain 2015, 138, 3238–3250. [Google Scholar] [CrossRef] [Green Version]
- Schossig, A.; Bloch-Zupan, A.; Lussi, A.; Wolf, N.I.; Raskin, S.; Cohen, M.; Giuliano, F.; Jurgens, J.; Krabichler, B.; Koolen, D.A.; et al. SLC13A5 Is the Second Gene Associated with Kohlschütter-Tönz Syndrome. J. Med. Genet. 2017, 54, 54–62. [Google Scholar] [CrossRef]
- Henke, C.; Töllner, K.; van Dijk, R.M.; Miljanovic, N.; Cordes, T.; Twele, F.; Bröer, S.; Ziesak, V.; Rohde, M.; Hauck, S.M.; et al. Disruption of the Sodium-Dependent Citrate Transporter SLC13A5 in Mice Causes Alterations in Brain Citrate Levels and Neuronal Network Excitability in the Hippocampus. Neurobiol. Dis. 2020, 143, 105018. [Google Scholar] [CrossRef]
- Fan, S.-Z.; Sung, C.-W.; Tsai, Y.-H.; Yeh, S.-R.; Lin, W.-S.; Wang, P.-Y. Nervous System Deletion of Mammalian INDY in Mice Mimics Dietary Restriction-Induced Memory Enhancement. J. Gerontol. Ser. A 2021, 76, 50–56. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Choi, E.Y.; Lapidus, R.; Zhang, L.; Huang, S.-M.; Shapiro, P.; Wang, H. Silencing of Solute Carrier Family 13 Member 5 Disrupts Energy Homeostasis and Inhibits Proliferation of Human Hepatocarcinoma Cells. J. Biol. Chem. 2017, 292, 13890–13901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Cordes, T.; Thalacker-Mercer, A.E.; Pajor, A.M.; Murphy, A.N.; Metallo, C.M. NaCT/SLC13A5 Facilitates Citrate Import and Metabolism under Nutrient-Limited Conditions. Cell Rep. 2021, 36, 109701. [Google Scholar] [CrossRef] [PubMed]
- Eniafe, J.; Jiang, S. The Functional Roles of TCA Cycle Metabolites in Cancer. Oncogene 2021, 40, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- Neuschäfer-Rube, F.; Lieske, S.; Kuna, M.; Henkel, J.; Perry, R.J.; Erion, D.M.; Pesta, D.; Willmes, D.M.; Brachs, S.; von Loeffelholz, C.; et al. The Mammalian INDY Homolog Is Induced by CREB in a Rat Model of Type 2 Diabetes. Diabetes 2014, 63, 1048–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, H.; Garzel, B.; Yang, H.; Sueyoshi, T.; Li, Q.; Shu, Y.; Zhang, J.; Hu, B.; Heyward, S.; et al. SLC13A5 Is a Novel Transcriptional Target of the Pregnane X Receptor and Sensitizes Drug-Induced Steatosis in Human Liver. Mol. Pharmacol. 2015, 87, 674–682. [Google Scholar] [CrossRef] [Green Version]
- Neuschäfer-Rube, F.; Schraplau, A.; Schewe, B.; Lieske, S.; Krützfeldt, J.-M.; Ringel, S.; Henkel, J.; Birkenfeld, A.L.; Püschel, G.P. Arylhydrocarbon Receptor-Dependent mINDY (Slc13a5) Induction as Possible Contributor to Benzo[a]Pyrene-Induced Lipid Accumulation in Hepatocytes. Toxicology 2015, 337, 1–9. [Google Scholar] [CrossRef]
- von Loeffelholz, C.; Lieske, S.; Neuschäfer-Rube, F.; Willmes, D.M.; Raschzok, N.; Sauer, I.M.; König, J.; Fromm, M.F.; Horn, P.; Chatzigeorgiou, A.; et al. The Human Longevity Gene Homolog INDY and Interleukin-6 Interact in Hepatic Lipid Metabolism: Liver Biology/Pathobiology. Hepatology 2017, 66, 616–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Fernandez, I.A.; Tauc, H.M.; Jasper, H. Hallmarks of Aging Drosophila Intestinal Stem Cells. Mech. Ageing Dev. 2020, 190, 111285. [Google Scholar] [CrossRef]
- Jasper, H. Intestinal Stem Cell Aging: Origins and Interventions. Annu. Rev. Physiol. 2020, 82, 203–226. [Google Scholar] [CrossRef] [Green Version]
- Funk, M.C.; Zhou, J.; Boutros, M. Ageing, Metabolism and the Intestine. EMBO Rep. 2020, 21. [Google Scholar] [CrossRef]
- Edgecomb, R.S.; Harth, C.E.; Schneiderman, A.M. Regulation of Feeding Behavior in Adult Drosophila melanogaster Varies with Feeding Regime and Nutritional State. J. Exp. Biol 1994, 197, 215–235. [Google Scholar] [CrossRef]
- Micchelli, C.A.; Perrimon, N. Evidence That Stem Cells Reside in the Adult Drosophila Midgut Epithelium. Nature 2006, 439, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Ohlstein, B.; Spradling, A. The Adult Drosophila Posterior Midgut Is Maintained by Pluripotent Stem Cells. Nature 2006, 439, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Edgar, B.A. Intestinal Stem Cell Function in Drosophila and Mice. Curr. Opin. Genet. Dev. 2012, 22, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Hur, J.H.; Bahadorani, S.; Graniel, J.; Koehler, C.L.; Ulgherait, M.; Rera, M.; Jones, D.L.; Walker, D.W. Increased Longevity Mediated by Yeast NDI1 Expression in Drosophila Intestinal Stem and Progenitor Cells. Aging (Albany NY) 2013, 5, 662–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rera, M.; Bahadorani, S.; Cho, J.; Koehler, C.L.; Ulgherait, M.; Hur, J.H.; Ansari, W.S.; Lo, T.; Jones, D.L.; Walker, D.W. Modulation of Longevity and Tissue Homeostasis by the Drosophila PGC-1 Homolog. Cell Metab. 2011, 14, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.P.; Rogina, B. A Gutsy Way to Extend Longevity. Front. Gene. 2012, 3, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagi, K.; Wilson, K.A.; Katewa, S.D.; Ortega, M.; Simons, J.; Hilsabeck, T.A.; Kapuria, S.; Sharma, A.; Jasper, H.; Kapahi, P. Dietary Restriction Improves Intestinal Cellular Fitness to Enhance Gut Barrier Function and Lifespan in D. Melanogaster. PLoS Genet. 2018, 14, e1007777. [Google Scholar] [CrossRef]
- Peters, J.M. Flipping a Citrate Switch on Liver Cancer Cells. J. Biol. Chem. 2017, 292, 13902–13903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudry, B.; de Goeij, E.; Mineo, A.; Gaspar, P.; Hadjieconomou, D.; Studd, C.; Mokochinski, J.B.; Kramer, H.B.; Plaçais, P.-Y.; Preat, T.; et al. Sex Differences in Intestinal Carbohydrate Metabolism Promote Food Intake and Sperm Maturation. Cell 2019, 178, 901–918.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchon, N.; Osman, D.; David, F.P.A.; Fang, H.Y.; Boquete, J.-P.; Deplancke, B.; Lemaitre, B. Morphological and Molecular Characterization of Adult Midgut Compartmentalization in Drosophila. Cell Rep. 2013, 3, 1725–1738. [Google Scholar] [CrossRef] [Green Version]
- Marianes, A.; Spradling, A.C. Physiological and Stem Cell Compartmentalization within the Drosophila Midgut. eLife 2013, 2, e00886. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.F.; Sousa, M.; Silva, B.M.; Monteiro, M.P.; Alves, M.G. Obesity, Energy Balance and Spermatogenesis. Reproduction 2017, 153, R173–R185. [Google Scholar] [CrossRef] [Green Version]
- Luo, D.; Zhang, M.; Su, X.; Liu, L.; Zhou, X.; Zhang, X.; Zheng, D.; Yu, C.; Guan, Q. High Fat Diet Impairs Spermatogenesis by Regulating Glucose and Lipid Metabolism in Sertoli Cells. Life Sci. 2020, 257, 118028. [Google Scholar] [CrossRef]
- Neubaum, D.M.; Wolfner, M.F. 3 Wise, Winsome, or Weird? Mechanisms of Sperm Storage in Female Animals. Curr. Top. Dev. Biol. 1998, 41, 67–97. [Google Scholar]
- Rogina, B.; Wolverton, T.; Bross, T.G.; Chen, K.; Müller, H.-G.; Carey, J.R. Distinct Biological Epochs in the Reproductive Life of Female Drosophila melanogaster. Mech. Ageing Dev. 2007, 128, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Rogina, B. The Effect of Sex Peptide and Calorie Intake on Fecundity in Female Drosophila melanogaster. Sci. World J. 2009, 9, 1178–1189. [Google Scholar] [CrossRef] [Green Version]
- Frankel, S.; Rogina, B. INDY Mutants: Live Long and Prosper. Front. Genet. 2012, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, J.C.; Khericha, M.; Dobson, A.J.; Bolukbasi, E.; Rattanavirotkul, N.; Partridge, L. Sex Difference in Pathology of the Ageing Gut Mediates the Greater Response of Female Lifespan to Dietary Restriction. Elife 2016, 5, e10956. [Google Scholar] [CrossRef] [PubMed]
- Millington, J.W.; Rideout, E.J. Sex Differences in Drosophila Development and Physiology. Curr. Opin. Physiol. 2018, 6, 46–56. [Google Scholar] [CrossRef]
- Choi, N.-H.; Kim, J.-G.; Yang, D.-J.; Kim, Y.-S.; Yoo, M.-A. Age-Related Changes in Drosophila Midgut Are Associated with PVF2, a PDGF/VEGF-like Growth Factor. Aging Cell 2008, 7, 318–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biteau, B.; Karpac, J.; Hwangbo, D.; Jasper, H. Regulation of Drosophila Lifespan by JNK Signaling. Exp. Gerontology 2011, 46, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Patel, P.H.; Kohlmaier, A.; Grenley, M.O.; McEwen, D.G.; Edgar, B.A. Cytokine/Jak/Stat Signaling Mediates Regeneration and Homeostasis in the Drosophila Midgut. Cell 2009, 137, 1343–1355. [Google Scholar] [CrossRef] [Green Version]
- Helfand, S.L.; Rogina, B. From Genes to Aging in Drosophila. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 2003; Volume 49, pp. 67–109. ISBN 978-0-12-017649-6. [Google Scholar]
- Willmes, D.M.; Daniels, M.; Kurzbach, A.; Lieske, S.; Bechmann, N.; Schumann, T.; Henke, C.; El-Agroudy, N.N.; Da Costa Goncalves, A.C.; Peitzsch, M.; et al. The Longevity Gene mINDY (I’m Not Dead, Yet) Affects Blood Pressure through Sympathoadrenal Mechanisms. JCI Insight 2021, 6, e136083. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Abbreviation | Description |
|---|---|
| ACLY | ATP citrate lyase |
| AhR | Aryl hydrocarbon receptor |
| AMPK | 5′AMP-activated protein kinase |
| CeNac2 | Caenorhabditis elegans sodium cotransporter 2 |
| CIC | Citrate/isocitrate carrier also known as SLC25A1 |
| CR | Calorie restriction |
| CREB | cAMP responsive element-binding protein |
| DILPs | Drosophila Insulin-like peptides |
| EB | Enteroblast |
| EC | Enterocyte |
| EE | Entero-endocrine cell |
| ETC | Electron transport chain |
| FoxO | Transcription factor from the Forkhead box-O family |
| GstD5 | Glutathione S transferase D5 |
| GstE1 | Glutathione S transferase E1 |
| HepG2 | Hepatoma G2 cell line |
| Hoppel | Transposable insertion variant |
| IIS | Insulin/insulin-like growth factor signaling pathway |
| IL-6 | Interleukin 6 |
| Indy | Drosophila I’m not dead yet gene |
| ISC | Intestinal stem cell |
| mINDY | Mammalian homolog of Indy (also referred as mSLC13A5/NaCT) |
| mSLC13A5 | Mammalian Sodium-dicarboxylate cotransporter solute carrier family 13, member 5 (also referred as mINDY/NaCT) |
| NaCT | Sodium (Na+) coupled citrate transporter (also referred as mSLC13A5; mINDY) |
| NAFLD | Non-alcoholic fatty liver disease |
| dPGC-1/spargel | Drosophila homologue of peroxisome proliferator-activated receptor gamma co-activator-1 (PGC-1) |
| PXR | Pregnane X receptor |
| ROS | Reactive oxygen species |
| SLC13A5 | Sodium-dicarboxylate cotransporter solute carrier family 13, member 5 (also referred as INDY/NaCT) |
| STAT3 | Signal transducer and activator of transcription 3 |
| TCA | Tricarboxylic acid cycle |
| VM | Visceral muscle |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kannan, K.; Rogina, B. The Role of Citrate Transporter INDY in Metabolism and Stem Cell Homeostasis. Metabolites 2021, 11, 705. https://doi.org/10.3390/metabo11100705
Kannan K, Rogina B. The Role of Citrate Transporter INDY in Metabolism and Stem Cell Homeostasis. Metabolites. 2021; 11(10):705. https://doi.org/10.3390/metabo11100705
Chicago/Turabian StyleKannan, Kavitha, and Blanka Rogina. 2021. "The Role of Citrate Transporter INDY in Metabolism and Stem Cell Homeostasis" Metabolites 11, no. 10: 705. https://doi.org/10.3390/metabo11100705
APA StyleKannan, K., & Rogina, B. (2021). The Role of Citrate Transporter INDY in Metabolism and Stem Cell Homeostasis. Metabolites, 11(10), 705. https://doi.org/10.3390/metabo11100705