
Variability of Lipids in Human Milk

 , , ,
, , , 
Abstract
:
1. Introduction
1.1. Analytical Challenge of Human Milk Lipidomic Analysis
1.2. Diurnal Variation of Breast Milk Components
1.3. Aims and Objectives
2. Results
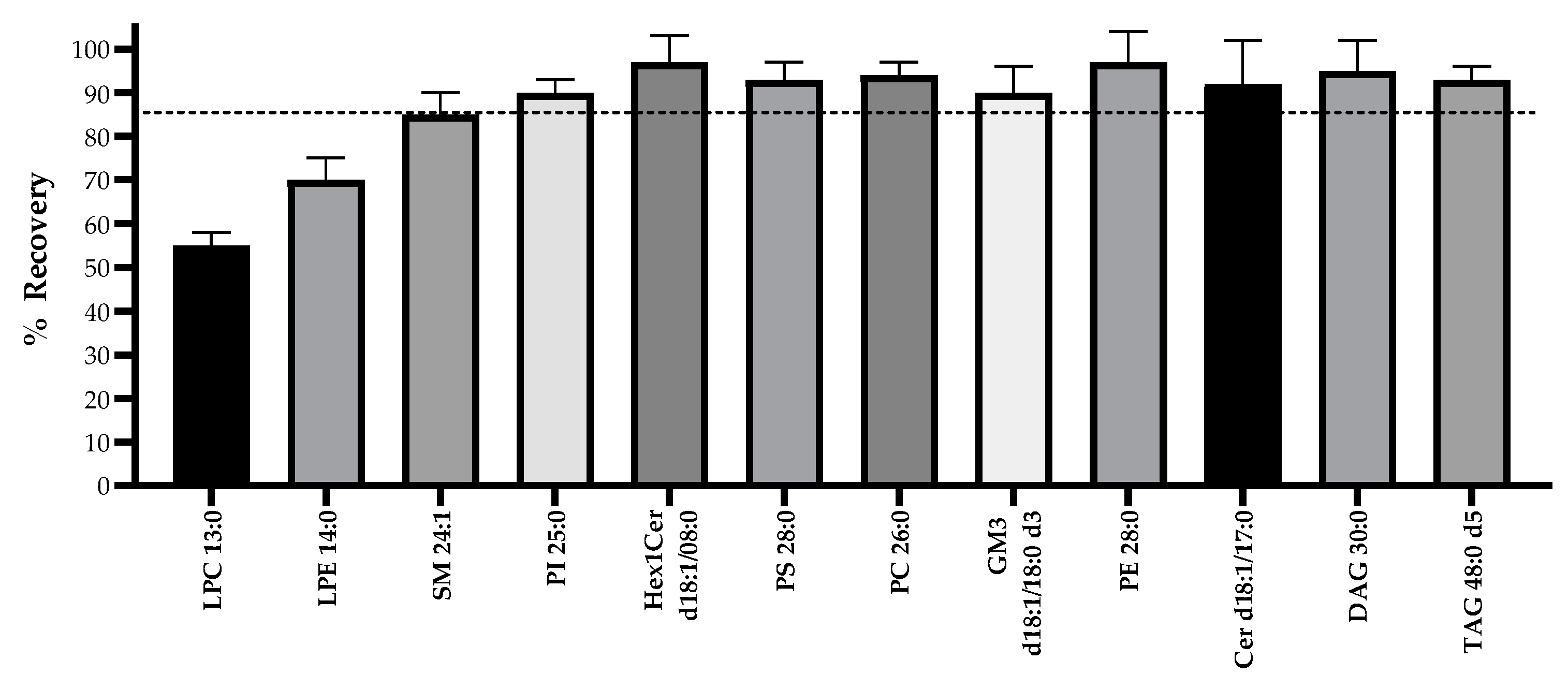
2.1. Optimization of the Analytical Workflow
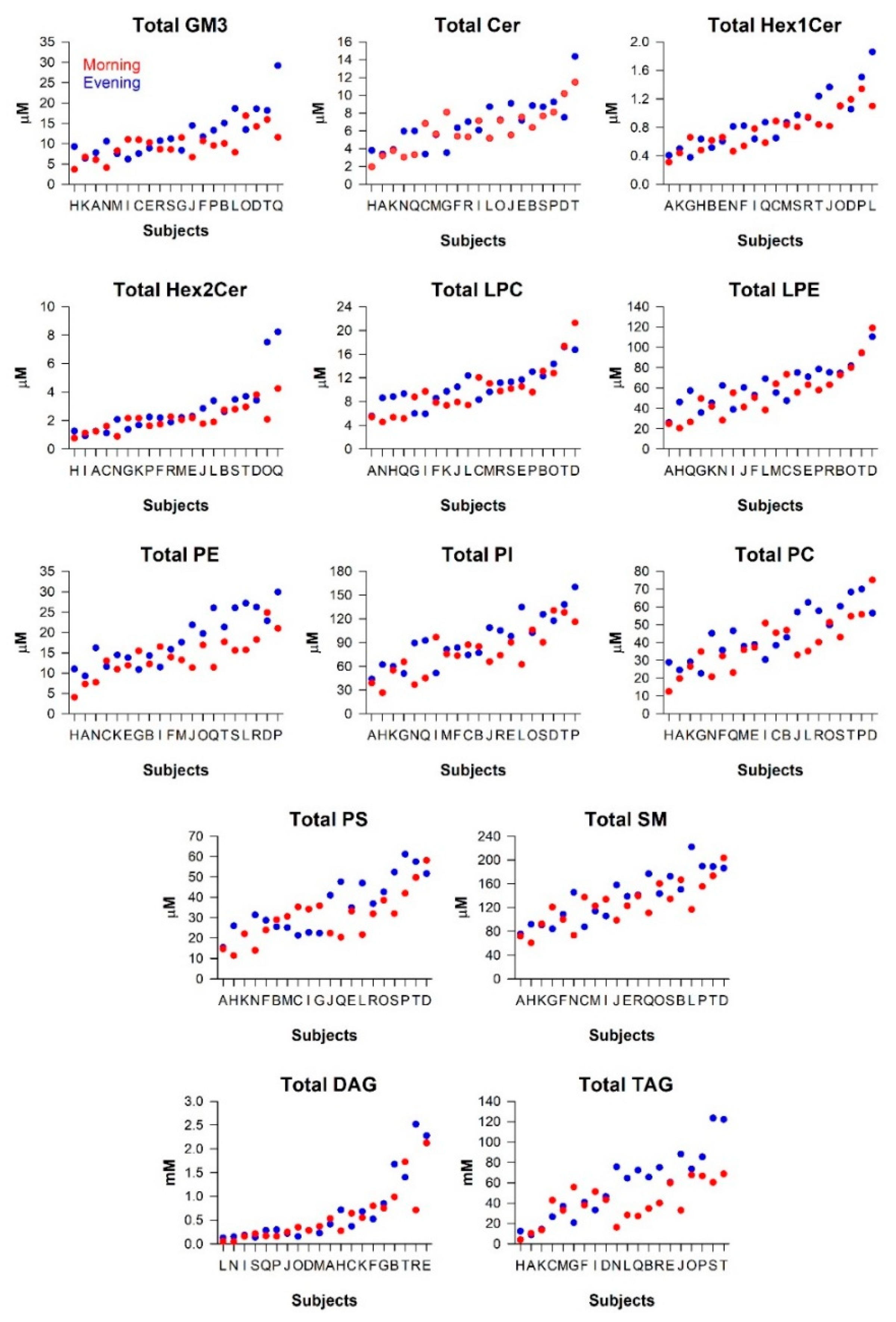
2.2. Variation in the Milk Lipidome between Participants and within Participants
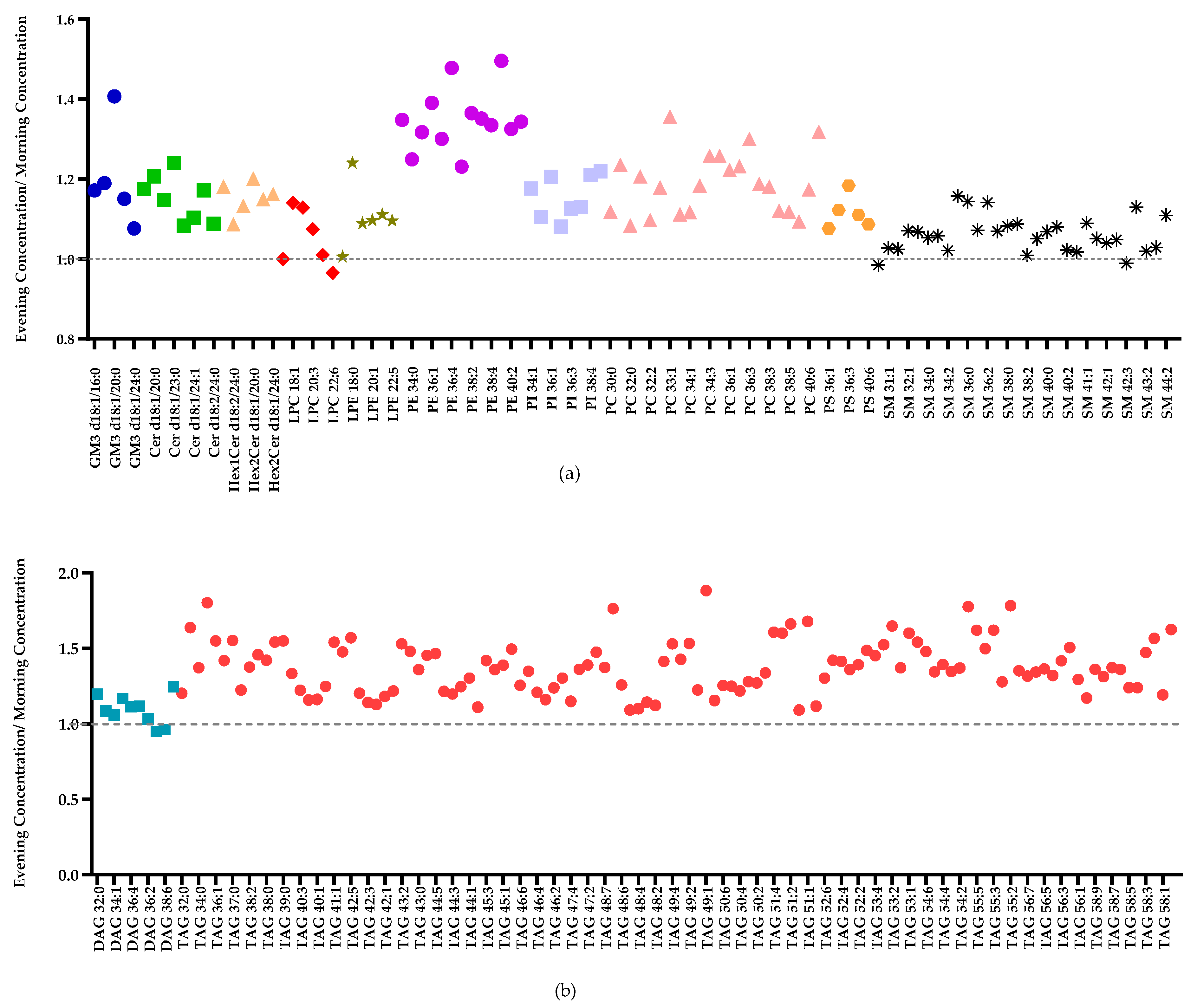
2.3. Inter-Individual Variation in the Human Milk Lipidome in the Morning and Evening
3. Discussion
3.1. Summary of Findings
3.2. Optimization of Analytical Workflow
3.3. Variability of Lipids in Human Milk
3.3.1. Inter-Individual Variation
3.3.2. Intra-Individual Variation
4. Materials and Methods
4.1. Participants and Recruitment
4.2. Chemicals for Sample Analysis
4.3. Lipid Extraction
4.4. Mass Spectrometric Method
4.4.1. Triacylglycerol and Diacylglycerol Measurements
4.4.2. Phospholipids and Sphingolipids Measurements
4.5. Method Validation/Quality Control
4.6. Internal Standards
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andreas, N.J.; Kampmann, B.; Le-Doare, K.M. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef]
- Eriksen, K.G.; Christensen, S.H.; Lind, M.V.; Michaelsen, K.F. Human milk composition and infant growth. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 200–206. [Google Scholar] [CrossRef]
- Stam, J.; Sauer, P.J.; Boehm, G. Can we define an infant’s need from the composition of human milk? Am. J. Clin. Nutr. 2013, 98, 521S–528S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenness, R. The composition of human milk. Semin. Perinatol. 1979, 3, 225–239. [Google Scholar]
- Ballard, O.; Morrow, A.L. Human milk composition. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Freed, L.M.; Neville, M.C.; Hamosh, P.; Hamosh, M. Diurnal and within-feed variations in lipase activity and triglyceride content of human milk. J. Pediatr. Gastroenterol. Nutr. 1986, 5, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine. Milk composition. In Nutrition During Lactation; The National Academies Press: Washington, DC, USA, 1991; Volume 6, p. 326. [Google Scholar]
- Mosca, F.; Giannì, M.L. Human milk: Composition and health benefits. Pediatr. Med. Chir. 2017, 39, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cilla, A.; Quintaes, K.D.; Barberá, R.; Alegría-Torán, A. Phospholipids in human milk and infant formulas: Benefits and needs for correct infant nutrition. Crit. Rev. Food Sci. Nutr. 2014, 56, 1880–1892. [Google Scholar] [CrossRef]
- Hernell, O.; Timby, N.; Domellöf, M.; Lönnerdal, B. Clinical benefits of milk fat globule membranes for infants and children. J. Pediatr. 2016, 173, S60–S65. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, X.; Wang, L. The investigation of fatty acid composition of breast milk and its relationship with dietary fatty acid intake in 5 regions of China. Medicine 2019, 98, e15855. [Google Scholar] [CrossRef]
- Sokol, E.; Ulven, T.; Færgeman, N.J.; Ejsing, C.S. Comprehensive and quantitative profiling of lipid species in human milk, cow milk and a phospholipid-enriched milk formula by GC and MS/MS ALL. Eur. J. Lipid Sci. Technol. 2015, 117, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demmelmair, H.; Koletzko, B. Lipids in human milk. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.G.; Lammi-Keefe, C.J.; Ferris, A.M.; Jackson, M.B.; Couch, S.C.; Capacchione, C.M.; Ahn, H.S.; Murtaugh, M. Human milk total lipid and cholesterol are dependent on interval of sampling during 24 Hours. J. Pediatr. Gastroenterol. Nutr. 1995, 20, 91–94. [Google Scholar] [CrossRef]
- Hewelt-Belka, W.; Garwolińska, D.; Belka, M.; Bączek, T.; Namieśnik, J.; Kot-Wasik, A. A new dilution-enrichment sample preparation strategy for expanded metabolome monitoring of human breast milk that overcomes the simultaneous presence of low- and high-abundance lipid species. Food Chem. 2019, 288, 154–161. [Google Scholar] [CrossRef]
- Garcia, C.; Lutz, N.W.; Confort-Gouny, S.; Cozzone, P.J.; Armand, M.; Bernard, M. Phospholipid fingerprints of milk from different mammalians determined by 31P NMR: Towards specific interest in human health. Food Chem. 2012, 135, 1777–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villaseñor, A.; Garcia-Perez, I.; Garcia, A.; Posma, J.M.; Fernández-López, M.; Nicholas, A.J.; Modi, N.; Holmes, E.; Barbas, C. Breast milk metabolome characterization in a single-phase extraction, multiplatform analytical approach. Anal. Chem. 2014, 86, 8245–8252. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Rochfort, S.; Cocks, B.G. Optimization of a single phase method for lipid extraction from milk. J. Chromatogr. A 2016, 1458, 145–149. [Google Scholar] [CrossRef]
- Chua, E.C.-P.; Shui, G.; Lee, I.T.-G.; Lau, P.; Tan, L.-C.; Yeo, S.-C.; Lam, B.D.; Bulchand, S.; Summers, S.A.; Puvanendran, K.; et al. Extensive diversity in circadian regulation of plasma lipids and evidence for different circadian metabolic phenotypes in humans. Proc. Natl. Acad. Sci. USA 2013, 110, 14468–14473. [Google Scholar] [CrossRef] [Green Version]
- Chua, E.C.-P.; Shui, G.; Gassiot, A.C.; Wenk, M.R.; Gooley, J.J. Changes in plasma lipids during exposure to total sleep deprivation. Sleep 2015, 38, 1683–1691. [Google Scholar] [CrossRef] [Green Version]
- Hall, B. Uniformity of human milk. Am. J. Clin. Nutr. 1979, 32, 304–312. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.A.; Imong, S.M.; Silprasert, A.; Ruckphaopunt, S.; Woolridge, M.W.; Baum, J.D.; Amatayakul, K. Circadian variation in fat concentration of breast-milk in a rural northern Thai population. Br. J. Nutr. 1988, 59, 349–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentice, A.; Prentice, A.M.; Whitehead, R.G. Breast-milk fat concentrations of rural African women. Br. J. Nutr. 1981, 45, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deem, H.E. Observations on the milk of New Zealand women. Arch. Dis. Child. 1931, 6, 53–70. [Google Scholar] [CrossRef] [Green Version]
- Gunther, M.; Stanier, J.E. Diurnal variation in the fat content of breast-milk. Lancet 1949, 2, 235–237. [Google Scholar] [CrossRef]
- Hytten, F.E. Clinical and chemical studies in human lactation—I, II, and III. BMJ 1954, 1, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.H.; Black, R.E.; Robertson, A.D.; Akhtar, N.A.; Ahmed, G.; Becker, S. Clinical and field studies of human lactation: Methodological considerations. Am. J. Clin. Nutr. 1982, 35, 745–756. [Google Scholar] [CrossRef] [Green Version]
- Nims, B.; Macy, I.G.; Brown, M.; Hunscher, H.A. Human milk studies. Am. J. Dis. Child. 1932, 43, 828. [Google Scholar] [CrossRef]
- Italianer, M.F.; Naninck, E.F.G.; Roelants, J.A.; Van Der Horst, G.; Reiss, I.K.; Van Goudoever, J.B.; Joosten, K.F.M.; Chaves, I.; Vermeulen, M.J. Circadian variation in human milk composition, a systematic review. Nutrients 2020, 12, 2328. [Google Scholar] [CrossRef]
- Karra, M.V.; Kirksey, A. Variation in zinc, calcium, and magnesium concentrations of human milk within a 24-hour period from 1 to 6 months of lactation. J. Pediatr. Gastroenterol. Nutr. 1988, 7, 100–106. [Google Scholar] [CrossRef]
- Picciano, M.F.; A Guthrie, H. Copper, iron, and zinc contents of mature human milk. Am. J. Clin. Nutr. 1976, 29, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Viverge, D.; Grimmonprez, L.; Cassanas, G.; Bardet, L.; Solère, M. Diurnal variations and within the feed in lactose and oligosaccharides of human milk. Ann. Nutr. Metab. 1986, 30, 196–209. [Google Scholar] [CrossRef] [PubMed]
- Illnerová, H.; Buresová, M.; Presl, J. Melatonin rhythm in human milk. J. Clin. Endocrinol. Metab. 1993, 77, 838–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Zavaleta, N.; Chen, S.-Y.; Lönnerdal, B.; Slupsky, C.M. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. NPJ Sci. Food 2018, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Alshehry, Z.H.; Barlow, C.K.; Weir, J.M.; Zhou, Y.; McConville, M.J.; Meikle, P.J. An efficient single phase method for the extraction of plasma lipids. Metabolites 2015, 5, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Ruel, M.T.; Dewey, K.G.; Martinez, C.; Flores, R.; Brown, K.H. Validation of single daytime samples of human milk to estimate the 24-h concentration of lipids in urban Guatemalan mothers. Am. J. Clin. Nutr. 1997, 65, 439–444. [Google Scholar] [CrossRef]
- Lammi-Keefe, C.J.; Ferris, A.M.; Jensen, R.G. Changes in human milk at 0600, 1000, 1400, 1800, and 2200 h. J. Pediatr. Gastroenterol. Nutr. 1990, 11, 83–88. [Google Scholar] [CrossRef]
- Lacruz, M.; Kluttig, A.; Tiller, D.; Medenwald, D.; Giegling, I.; Rujescu, D.; Prehn, C.; Adamski, J.; Greiser, K.H.; Kastenmüller, G. Instability of personal human metabotype is linked to all-cause mortality. Sci. Rep. 2018, 8, 9810. [Google Scholar] [CrossRef] [Green Version]
- Mitoulas, L.R.; Kent, J.C.; Cox, D.; Owens, R.; Sherriff, J.; Hartmann, P. Variation in fat, lactose and protein in human milk over 24 h and throughout the first year of lactation. Br. J. Nutr. 2002, 88, 29–37. [Google Scholar] [CrossRef]
- Blaas, N.; Schüürmann, C.; Bartke, N.; Stahl, B.; Humpf, H.-U. Structural profiling and quantification of sphingomyelin in human breast milk by HPLC-MS/MS. J. Agric. Food Chem. 2011, 59, 6018–6024. [Google Scholar] [CrossRef] [PubMed]
- Oshida, K.; Shimizu, T.; Takase, M.; Tamura, Y.; Shimizu, T.; Yamashiro, Y. Effects of dietary sphingomyelin on central nervous system myelination in developing rats. Pediatr. Res. 2003, 53, 589–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.; Owens, R.A.; Hartmann, P.E. Blood and milk prolactin and the rate of milk synthesis in women. Exp. Physiol. 1996, 81, 1007–1020. [Google Scholar] [CrossRef]
- Cregan, M.D.; Mitoulas, L.R.; Hartmann, P.E. Milk prolactin, feed volume and duration between feeds in women breastfeeding their full-term infants over a 24 h period. Exp. Physiol. 2002, 87, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Villalpando, S.; Del Prado, M. Interrelation among dietary energy and fat intakes, maternal body fatness, and milk total lipid in humans. J. Mammary Gland. Biol. Neoplasia 1999, 4, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Harzer, G.; Haug, M.; Dieterich, I.; Gentner, P.R. Changing patterns of human milk lipids in the course of the lactation and during the day. Am. J. Clin. Nutr. 1983, 37, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Picciano, M.F.; Guthrie, H.A.; Sheehe, D.M. The cholesterol content of human milk. Clin. Pediatr. 1978, 17, 359–362. [Google Scholar] [CrossRef]
- Mien, I.H.; Chua, E.C.-P.; Lau, P.; Tan, L.-C.; Lee, I.T.-G.; Yeo, S.C.; Tan, S.S.; Gooley, J.J. Effects of exposure to intermittent versus continuous red light on human circadian rhythms, melatonin suppression, and pupillary constriction. PLoS ONE 2014, 9, e96532. [Google Scholar] [CrossRef] [Green Version]
- Gooley, J.J.; Chua, E.C.-P. Diurnal regulation of lipid metabolism and applications of circadian lipidomics. J. Genet. Genom. 2014, 41, 231–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guthrie, H.A.; Picciano, M.F.; Sheehe, D. Fatty acid patterns of human milk. J. Pediatr. 1977, 90, 39–41. [Google Scholar] [CrossRef]
- Lopez, C.; Cauty, C.; Guyomarc’h, F. Unraveling the complexity of milk fat globules to tailor bioinspired emulsions providing health benefits: The key role played by the biological membrane. Eur. J. Lipid Sci. Technol. 2018, 121, 1800201. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.; Ménard, O. Human milk fat globules: Polar lipid composition and in situ structural investigations revealing the heterogeneous distribution of proteins and the lateral segregation of sphingomyelin in the biological membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Stafford, J.; Villalpando, S.; Aguila, B.U. Circadian variation and changes after a meal in volume and lipid production of human milk from rural mexican women. Ann. Nutr. Metab. 1994, 38, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, H.P.A.; Baynard, M.D.; Maislin, G.; Dinges, D.F. Systematic interindividual differences in neurobehavioral impairment from sleep loss: Evidence of trait-like differential vulnerability. Sleep 2004, 27, 423–433. [Google Scholar]
- Chua, E.C.; Yeo, S.C.; Lee, I.T.; Tan, L.; Lau, P.; Tan, S.S.; Mien, I.H.; Gooley, J.J. Individual differences in physiologic measures are stable across repeated exposures to total sleep deprivation. Physiol. Rep. 2014, 2, e12129. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variance Analysis | Time of Day | Order of Sample | |||||
|---|---|---|---|---|---|---|---|
| Lipid class | Z | p | ICC | F | p | F | p |
| GM3 | 1.43 | 0.076 | 0.35 | 5.21 | 0.035 | 1.90 | 0.19 |
| Cer | 2.27 | 0.012 | 0.61 | 1.66 | 0.214 | 1.46 | 0.24 |
| Hex1Cer | 2.45 | 0.007 | 0.68 | 3.85 | 0.065 | 0.78 | 0.39 |
| Hex2Cer | 1.83 | 0.034 | 0.46 | 3.78 | 0.068 | 0.00 | 0.99 |
| LPC | 2.49 | 0.006 | 0.70 | 1.17 | 0.293 | 0.33 | 0.57 |
| LPE | 2.54 | 0.006 | 0.72 | 3.26 | 0.088 | 0.66 | 0.43 |
| PE | 2.10 | 0.018 | 0.55 | 13.72 | 0.002 | 1.48 | 0.24 |
| PI | 2.13 | 0.017 | 0.56 | 5.81 | 0.027 | 1.25 | 0.28 |
| PC | 2.04 | 0.020 | 0.53 | 4.15 | 0.057 | 1.62 | 0.22 |
| PS | 1.92 | 0.027 | 0.49 | 3.84 | 0.066 | 0.85 | 0.37 |
| SM | 1.90 | 0.029 | 0.49 | 2.35 | 0.143 | 0.56 | 0.47 |
| DAG | 2.59 | 0.005 | 0.74 | 1.30 | 0.270 | 1.45 | 0.24 |
| TAG | 1.89 | 0.029 | 0.48 | 7.95 | 0.011 | 1.18 | 0.29 |
| 20 Most variable lipids | |||||
|---|---|---|---|---|---|
| Lipid | Biological Variation (%) Morning | Technical Variation % (QC) | Lipid | Biological Variation (%) Evening | Technical Variation % (QC) |
| DAG 38:6 | 118 | 5 | Hex2Cer d18:1/16:0 | 172 | 11 |
| TAG 47:0 | 114 | 7 | DAG 38:6 | 149 | 5 |
| DAG 32:0 | 111 | 8 | DAG 32:0 | 126 | 8 |
| DAG 34:2 | 105 | 6 | DAG 36:1 | 121 | 8 |
| TAG 43:0 | 103 | 7 | DAG 34:0 | 119 | 8 |
| DAG 36:1 | 103 | 8 | DAG 34:1 | 115 | 6 |
| TAG 45:0 | 103 | 7 | DAG 34:2 | 112 | 6 |
| DAG 34:1 | 102 | 6 | TAG 47:0 | 110 | 7 |
| DAG 36:4 | 101 | 6 | TAG 32:0 | 110 | 9 |
| DAG 34:0 | 101 | 8 | TAG 42:5 | 109 | 21 |
| TAG 36:2 | 99 | 8 | TAG 43:0 | 106 | 7 |
| TAG 38:3 | 97 | 9 | TAG 45:0 | 104 | 7 |
| TAG 42:5 | 95 | 21 | TAG 50:8 | 101 | 16 |
| TAG 41:0 | 94 | 9 | TAG 34:1 | 101 | 10 |
| TAG 50:8 | 93 | 16 | TAG 41:0 | 100 | 9 |
| TAG 34:1 | 92 | 10 | TAG 37:0 | 100 | 21 |
| DAG 36:3 | 92 | 5 | TAG 34:0 | 100 | 9 |
| TAG 60:3 | 92 | 16 | TAG 38:3 | 100 | 9 |
| TAG 49:1 | 91 | 7 | TAG 44:6 | 100 | 15 |
| DAG 36:2 | 90 | 6 | TAG 36:2 | 97 | 8 |
| 20 Least Variable Lipids | |||||
|---|---|---|---|---|---|
| Lipid | Biological Variation (%) Morning | Technical Variation % (QC) | Lipid | Biological Variation (%) Evening | Technical Variation % (QC) |
| SM 40:2 | 27 | 15 | SM 39:1 | 29 | 16 |
| SM 40:1 | 27 | 11 | SM 44:1 | 29 | 10 |
| SM 38:1 | 28 | 16 | SM 36:0 | 30 | 11 |
| SM 38:2 | 28 | 11 | SM 38:1 | 30 | 16 |
| SM 39:1 | 28 | 16 | PI 36:1 | 32 | 9 |
| SM 40:0 | 29 | 11 | PE 38:4 | 32 | 8 |
| SM 38:0 | 30 | 13 | SM 41:1 | 32 | 9 |
| PE 38:4 | 30 | 8 | SM 40:2 | 32 | 15 |
| PC 38:4 | 31 | 12 | PC 34:2 | 32 | 11 |
| GM3 d18:1/22:0 | 31 | 15 | LPC 18:1 | 32 | 15 |
| SM 41:1 | 32 | 9 | SM 36:1 | 32 | 14 |
| SM 44:1 | 32 | 10 | LPC 22:5 | 32 | 15 |
| SM 37:1 | 32 | 12 | GM3 d18:1/22:0 | 33 | 15 |
| SM 36:1 | 32 | 14 | SM 34:1 | 33 | 10 |
| SM 40:3 | 32 | 15 | SM 40:1 | 33 | 11 |
| SM 36:0 | 32 | 11 | PC 32:0 | 34 | 7 |
| Hex1Cer d18:2/22:0 | 32 | 8 | PC 38:4 | 34 | 12 |
| LPE 18:0 | 33 | 7 | PC 34:0 | 34 | 10 |
| SM 42:1 | 34 | 10 | PC 34:1 | 34 | 8 |
| SM 34:1 | 34 | 10 | SM 38:0 | 34 | 13 |
| Characteristic | Mean (SD) | Range |
|---|---|---|
| Maternal age (y) | 29.80 (2.63) | 25–35 |
| Maternal height (m) | 1.60 (0.05) | 1.53–1.73 |
| Maternal weight (kg) | 56.58 (6.43) | 46–72 |
| Maternal BMI | 22.01 (2.00) | 19.40–27.43 |
| Gestational age of baby at delivery (weeks) | 39.15 (0.75) | 38–40 |
| Age of child (days) | 86.40 (8.23) | 62–101 |
| Birth weight of baby (kg) | 3.19 (0.33) | 2.44–3.86 |
| Pre-pregnancy weight of mother (kg) | 52.23 (6.00) | 43–65 |
| Number (%) | ||
| Ethnicity | 19 Chinese (95), 1 Malay (5) | |
| Delivery mode (Vaginal) | 15 (75) | |
| Sex of child (Female) | 10 (50) | |
| Evening | Morning | p Value | |||
|---|---|---|---|---|---|
| Mean (SD) | Range | Mean (SD) | Range | ||
| Total volume of breast milk expressed (mL) | 70.58 (31.53) | 25–130 | 84.45 (52.79) | 20–225 | 0.10 |
| Time taken for milk expression (h:mm) | 0:17 (0:06) | 0:07–0:33 | 0:18 (0:06) | 0:08–0:34 | 0.31 |
| Gap between previous feed and sample (h:mm) | 2:07 (1:10) | 0:13–4:15 | 2:17 (1:56) | 0:18–9:07 | 0.75 |
| Number of feeds between first and second sample | 4.50 (1.50) | 2–8 | 4.15 (1.31) | 2–7 | 0.35 |
| Number of participants who had dinner before evening sample, N (%) | 11 (55) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvalatchmanan, J.; Rukmini, A.V.; Ji, S.; Triebl, A.; Gao, L.; Bendt, A.K.; Wenk, M.R.; Gooley, J.J.; Torta, F. Variability of Lipids in Human Milk. Metabolites 2021, 11, 104. https://doi.org/10.3390/metabo11020104
Selvalatchmanan J, Rukmini AV, Ji S, Triebl A, Gao L, Bendt AK, Wenk MR, Gooley JJ, Torta F. Variability of Lipids in Human Milk. Metabolites. 2021; 11(2):104. https://doi.org/10.3390/metabo11020104
Chicago/Turabian StyleSelvalatchmanan, Jayashree, A.V. Rukmini, Shanshan Ji, Alexander Triebl, Liang Gao, Anne K. Bendt, Markus R. Wenk, Joshua J. Gooley, and Federico Torta. 2021. "Variability of Lipids in Human Milk" Metabolites 11, no. 2: 104. https://doi.org/10.3390/metabo11020104
APA StyleSelvalatchmanan, J., Rukmini, A. V., Ji, S., Triebl, A., Gao, L., Bendt, A. K., Wenk, M. R., Gooley, J. J., & Torta, F. (2021). Variability of Lipids in Human Milk. Metabolites, 11(2), 104. https://doi.org/10.3390/metabo11020104