
Host-Microbiome Interactions Mediated by Phenolic Metabolites in Chronically Critically Ill Patients

 , ,
, , 
 and
and
Abstract
:1. Introduction
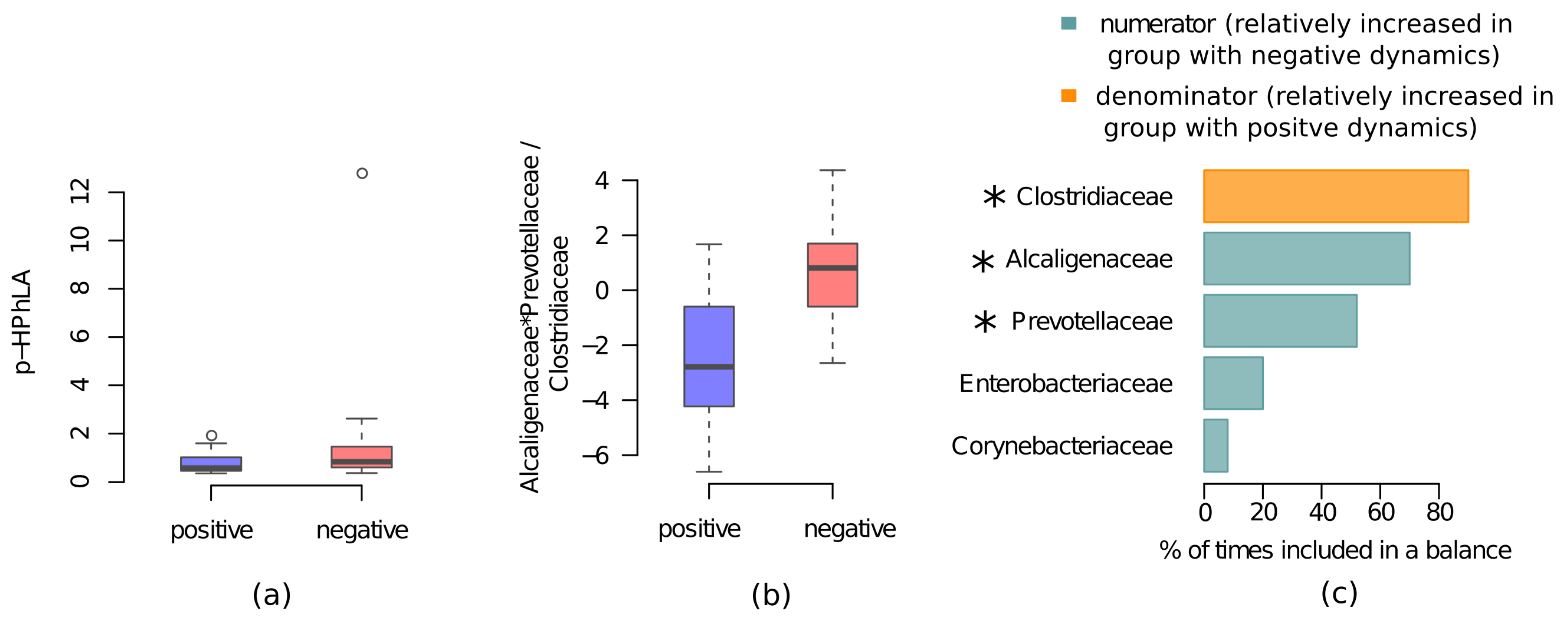
2. Results
2.1. Metabolic and Microbiome Differences between Healthy Subjects and CCI Patients
2.2. Metabolic and Microbiome Differences between the CCI Patients with Positive and Negative Clinical and Neurological Dynamics
2.3. Association between Metabolites, Biomarkers and Microbiome
2.4. Associations of Taxa and Metabolites with the Therapy
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Patients and Samples
4.3. Reagents and Chemicals
4.4. Serum Sample Preparation and GC-MS Analysis
4.5. Microbiome Sample Preparation
4.6. Microbiome Data Processing
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, J.E.; Cox, C.E.; Hope, A.A.; Carson, S.S. Chronic critical illness. Am. J. Respir. Crit. Care Med. 2010, 182, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.E. Persistent systemic inflammation in chronic critical illness. Respir. Care 2012, 57, 859–864. [Google Scholar] [CrossRef]
- Parfenov, A.L.; Petrova, M.V.; Pichugina, I.M.; Luginina, E.V. Comorbidity Development in Patients with Severe Brain Injury Resulting in Chronic Critical Condition. General Reanimatol. 2020, 16, 72–89. [Google Scholar] [CrossRef]
- Giau, V.V.; Wu, S.Y.; Jamerlan, A.; Soo, S.A.; Kim, A.N.; Hulme, J. Gut microbiota and their neuroinflammatory implications in Alzheimer’s disease. Nutrients 2018, 10, 1765. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.; Borchers, F.; Endres, M.; Weiss, B.; Spies, C.; Emmrich, J.V. Cognitive deficits following intensive care. Dtsch. Ärzteblatt Int. 2019, 116, 627–634. [Google Scholar] [CrossRef]
- Ojima, M.; Motooka, D.; Shimizu, K.; Gotoh, K.; Shintani, A.; Yoshiya, K.; Nakamura, S.; Ogura, H.; Iida, T.; Shimazu, T. Metagenomic analysis reveals dynamic changes of whole gut microbiota in the acute phase of intensive care unit patients. Dig. Dis. Sci. 2016, 61, 1628–1634. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Ackermann, G.; Khailova, L.; Baird, C.; Heyland, D.; Kozar, R.; Lemieux, M.; Derenski, K.; King, J.; Vis-Kampen, C.; et al. Extreme dysbiosis of the microbiome in critical illness. mSphere 2016, 1, e00199-16. [Google Scholar] [CrossRef] [Green Version]
- Haak, B.W.; Levi, M.; Wiersinga, W.J. Microbiota-targeted therapies on the intensive care unit. Curr. Opin. Crit. Care 2017, 23, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Klingensmith, N.J.; Coopersmith, C.M. New insights into the gut as the driver of critical illness and organ failure. Curr. Opin. Crit. Care 2017, 23, 143–148. [Google Scholar] [CrossRef]
- Otani, S.; Coopersmith, C.M. Gut integrity in critical illness. J. Intensive Care 2019, 20, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, H.; Hayes, J.; Boivin, M.; Tomba, T. Transpyloric feeding tube placement in critically ill patients using electromyogram and erythromycin infusion. Chest 2004, 125, 587–591. [Google Scholar] [CrossRef]
- Luft, V.M. Sovremennye vozmozhnosti nutricionnoj podderzhki bol’nyh v intensivnoj medicine. Vestn. Anesteziol. Reanimatol. 2010, 7, 42–51. [Google Scholar]
- Beloborodova, N.; Pautova, A.; Sergeev, A.; Fedotcheva, N. Serum Levels of Mitochondrial and Microbial Metabolites Reflect Mitochondrial Dysfunction in Different Stages of Sepsis. Metabolites 2019, 9, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenderov, B.A.; Sinitsa, A.V.; Zakharchenko, M.M.; Lang, C. Metabolic Relationship Between the Host and Its Gut Microbiota. In Metabiotics; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Beloborodova, N.V.; Khodakova, A.S.; Bairamov, I.T.; Olenin, A.Y. Microbial origin of phenylcarboxylic acids in the human body. Biochemistry 2009, 74, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Fedotcheva, N.I.; Kazakov, R.E.; Kondrashova, M.N.; Beloborodova, N.V. Toxic effects of microbial phenolic acids on the functions of mitochondria. Toxicol. Lett. 2008, 180, 182–188. [Google Scholar] [CrossRef]
- Beloborodova, N.V.; Moroz, V.V.; Bedova, A.Y.; Osipov, A.A.; Sarshor, Y.N.; Chernevskaya, E.A. Participation of aromatic microbial metabolites in the development of severe infection and sepsis. Anesteziol. Reanimatol. 2016, 61, 202–208. [Google Scholar]
- Beloborodova, N.V.; Sarshor, Y.N.; Bedova, A.Y.; Chernevskaya, E.A.; Pautova, A.K. Involvement of aromatic metabolites in the pathogenesis of septic shock. Shock 2018, 50, 273–279. [Google Scholar] [CrossRef]
- Moroz, V.V.; Beloborodova, N.V.; Osipov, A.A.; Vlasenko, A.V.; Bedova, A.Y.; Pautova, A.K. Phenylcarboxylic acids in the assessment of the severity of patient condition and the efficiency of intensive treatment in critical care medicine. Gen. Reanimatol. 2016, 12, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Chernevskaya, E.; Beloborodova, N.; Klimenko, N.; Pautova, A.; Shilkin, D.; Gusarov, V.; Tyakht, A. Serum and fecal profiles of aromatic microbial metabolites reflect gut microbiota disruption in critically ill patients: A prospective observational pilot study. Crit. Care 2020, 24, 312. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Empadinhas, N. The Microbiome-Mitochondria Dance in Prodromal Parkinson’s Disease. Front. Physiol. 2018, 9, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautova, A.K.; Bedova, A.Y.; Sarshor, Y.N.; Beloborodova, N.V. Determination of aromatic microbial metabolites in blood serum by gas chromatography–mass spectrometry. J. Analyt. Chem. 2018, 73, 160–166. [Google Scholar] [CrossRef]
- Rivera-Pinto, J.; Egozcue, J.J.; Pawlowsky-Glahn, V.; Paredes, R.; Noguera-Julian, M.; Calle, M.L. Balances: A New Perspective for Microbiome Analysis. mSystems 2018, 3, e00053-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaborin, A.; Smith, D.; Garfield, K.; Quensen, J.; Shakhsheer, B.; Kade, M.; Tirrell, M.; Tiedje, J.; Gilbert, J.A.; Zaborina, O.; et al. Membership and behavior of ultra-low-diversity pathogen communities present in the gut of humans during prolonged critical illness. mBio 2014, 5, e01361-14. [Google Scholar] [CrossRef] [Green Version]
- Ravi, A.; Halstead, F.D.; Bamford, A.; Casey, A.; Thomson, N.M.; Schaik, W.; Snelson, C.; Goulden, R.; Foster-Nyarko, E.; Savva, G.M.; et al. Loss of microbial diversity and pathogen domination of the gut microbiota in critically ill patients. Microb. Genom. 2019, 5, e000293. [Google Scholar] [CrossRef]
- Li, N.; Wang, X.; Sun, C.; Wu, X.; Lu, M.; Si, Y.; Ye, X.; Wang, T.; Yu, X.; Zhao, X.; et al. Change of intestinal microbiota in cerebral ischemic stroke patients. BMC Microbiol. 2019, 19, 191. [Google Scholar] [CrossRef] [Green Version]
- Leslie, D.L.; Kozma, L.; Martin, A.; Landeros, A.; Katsovich, L.; King, R.A.; Leckman, J.F. Neuropsychiatric disorders associated with streptococcal infection: A case-control study among privately insured children. J. Am. Acad. Child. Adolesc. Psychiatry 2008, 47, 1166–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.; Ding, B.; Feng, C.; Yin, S.; Zhang, T.; Qi, X.; Lv, H.; Guo, X.; Dong, K.; Zhu, Y.; et al. Prevotella and Klebsiella proportions in fecal microbial communities are potential characteristic parameters for patients with major depressive disorder. J. Affect. Disord. 2017, 207, 300–304. [Google Scholar] [CrossRef]
- Bo, J.; Wang, S.; Bi, Y.; Ma, S.; Wang, M.; Du, Z. Eggerthella lenta bloodstream infections: Two cases and review of the literature. Future Microbiol. 2020, 15, 981–985. [Google Scholar] [CrossRef]
- Rekdal, V.M.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and inhibition of an interspecies gut bacterial pathway for Levodopa metabolism. Science 2019, 364, eaau6323. [Google Scholar] [CrossRef] [Green Version]
- Ticinesi, A.; Mancabelli, L.; Tagliaferri, S.; Nouvenne, A.; Milani, C.; Del Rio, D.; Lauretani, F.; Maggio, M.G.; Ventura, M.; Meschi, T. The Gut-Muscle Axis in Older Subjects with Low Muscle Mass and Performance: A Proof of Concept Study Exploring Fecal Microbiota Composition and Function with Shotgun Metagenomics Sequencing. Int. J. Mol. Sci. 2020, 21, 8946. [Google Scholar] [CrossRef]
- Muñiz Pedrogo, D.A.; Chen, J.; Hillmann, B.; Jeraldo, P.; Al-Ghalith, G.; Taneja, V.; Davis, J.M.; Knights, D.; Nelson, H.; Faubion, W.A.; et al. An Increased Abundance of Clostridiaceae Characterizes Arthritis in Inflammatory Bowel Disease and Rheumatoid Arthritis: A Cross-sectional Study. Inflamm. Bowel Dis. 2019, 25, 902–913. [Google Scholar] [CrossRef]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerlings, S.Y.; Kostopoulos, I.; de Vos, W.M.; Belzer, C. Akkermansia muciniphila in the human gastrointestinal tract: When, where, and how? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the human gut microbiome in multiple sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar] [CrossRef]
- Toma, I.; Kang, J.J.; Sipos, A.; Vargas, S.; Bansal, E.; Hanner, F.; Meer, E.; Peti-Peterdi, J. Succinate receptor GPR91 provides a direct link between high glucose levels and renin release in murine and rabbit kidney. J. Clin. Investig. 2008, 118, 2526–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.D.; Prykhodko, O.; Fåk Hållenius, F.; Nyman, M. Monovalerin and trivalerin increase brain acetic acid, decrease liver succinic acid, and alter gut microbiota in rats fed high-fat diets. Eur. J. Nutr. 2019, 58, 1545–1560. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Chernevskaya, E.A.; Meglei, A.Y.; Buyakova, I.V.; Kovaleva, N.Y.; Gorshkov, K.M.; Zakharchenko, V.E.; Beloborodova, N.V. Taxonomic dysbiosis of gut microbiota and serum biomarkers reflect severity of central nervous system injury. Bull. RSMU 2020, 5, 54–61. [Google Scholar] [CrossRef]
- Liu, B.; Zhu, X.; Cui, Y.; Wang, W.; Liu, H.; Li, Z.; Guo, Z.; Ma, S.; Li, D.; Wang, C.; et al. Consumption of Dietary Fiber from Different Sources during Pregnancy Alters Sow Gut Microbiota and Improves Performance and Reduces Inflammation in Sows and Piglets. mSystems 2021, 6, e00591-20. [Google Scholar] [CrossRef]
- Iablokov, S.N.; Klimenko, N.S.; Efimova, D.A.; Shashkova, T.; Novichkov, P.S.; Rodionov, D.A.; Tyakht, A.V. Metabolic phenotypes as potential biomarkers for linking gut microbiome with inflammatory bowel diseases. Front. Mol. Biosci. 2020, 7, 603740. [Google Scholar] [CrossRef]
- Saver, J.L.; Filip, B.; Hamilton, S.; Yanes, A.; Craig, S.; Cho, M.; Conwit, R.; Starkman, S. Improving the reliability of stroke disability grading in clinical trials and clinical practice: The Rankin Focused Assessment (RFA). Stroke 2010, 41, 992–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimenko, N.S.; Tyakht, A.V.; Popenko, A.S.; Vasiliev, A.S.; Altukhov, I.A.; Ischenko, D.S.; Shashkova, T.I.; Efimova, D.A.; Nikogosov, D.A.; Osipenko, D.A.; et al. Microbiome Responses to an Uncontrolled Short-Term Diet Intervention in the Frame of the Citizen Science Project. Nutrients 2018, 10, 576. [Google Scholar] [CrossRef] [Green Version]
- Efimova, D.; Tyakht, A.; Popenko, A.; Vasilyev, A.; Altukhov, I.; Dovidchenko, N.; Odintsova, V.; Klimenko, N.; Loshkarev, R.; Pashkova, M.; et al. Knomics-Biota—A system for exploratory analysis of human gut microbiota data. BioData Min. 2018, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, H.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Andrianova, N.V.; Popkov, V.A.; Klimenko, N.S.; Tyakht, A.V.; Baydakova, G.V.; Frolova, O.Y.; Zorova, L.D.; Pevzner, I.B.; Zorov, D.B.; Plotnikov, E.Y. Microbiome-Metabolome Signature of Acute Kidney Injury. Metabolites 2020, 10, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, B.W.; Bonder, M.J.; Huse, S.M.; Zaura, E. TaxMan: A Server to Trim rRNA Reference Databases and Inspect Taxonomic Coverage. Nucleic Acids Res. 2012, 40, W82–W87. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Aitchison, J. The Statistical Analysis of Compositional Data. J. R. Stat. Soc. 1982, 44, 139–160. [Google Scholar] [CrossRef]
- Martín-Fernández, J.-A.; Hron, K.; Templ, M.; Filzmoser, P.; Palarea-Albaladejo, J. Bayesian-Multiplicative Treatment of Count Zeros in Compositional Data Sets. Stat. Model. 2015, 15, 134–158. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | CCI Patients | Healthy Subjects * | FDR |
|---|---|---|---|
| Microbial Phenolic Metabolites µM | |||
| PhLA | 0.2 (0.1–0.4) | <LOD ** | 1.9 × 10−7 |
| p-HPhAA | 0.7 (0.3–1.8) | <LOD ** | >0.05 |
| p-HPhLA | 0.8 (0.6–1.1) | 0.7 (0.6–1.0) | >0.05 |
| BA | 0.5 (0.3–0.8) | 0.4 (0.3–0.8) | >0.05 |
| p-HBA | 0.2 (0–0.9) | <LOD ** | 5.3 × 10−10 |
| Mitochondrial Metabolites, µM | |||
| Succinic Acid (SA) | 6.3 (4.4–10.1) | 22.0 (15.5–26.2) | 6.3 × 10−6 |
| Fumaric Acid (FA) | 0.6 (0.3–1.2) | 1.8 (1.3–2.4) | 6.3 × 10−6 |
| Biomarkers *** | |||
| Procalcitonin | 0.06 (0.03–0.09) | 0.25 ng/mL | - |
| Interleukin-6 | 21 (12–33) | < 7 pg/mL | - |
| S100 protein | 0.06 (0.04–0.1) | < 0.1 μg/L | - |
| Taxon/Metabolite | Neurological Scales | Rank | P Value | FDR | Linear Model Coefficient |
|---|---|---|---|---|---|
| (unclassified/Clostridiaceae/Lachnospiraceae)_u_u | Rivermead Mobility Index | species | 3.3 × 10−6 | 8.3 × 10−4 | 0.2962 |
| Ruminococcus (bromii/unclassified) | Rivermead Mobility Index | species | 4.0 × 10−4 | 3.6 × 10−2 | 0.2521 |
| (unclassified/Clostridiaceae/Lachnospiraceae)_u | Rivermead Mobility Index | genus | 4.4 × 10−6 | 8.8 × 10−4 | 0.2955 |
| (unclassified/Clostridiaceae/Lachnospiraceae) | Rivermead Mobility Index | family | 1.2 × 10−5 | 2.0 × 10−3 | 0.3200 |
| (unclassified/Clostridiaceae/Lachnospiraceae)_u_u | Rankin | species | 6.2 × 10−7 | 2.5 × 10−4 | −1.0954 |
| (Coprococcus/unclassified)_u | Rankin | species | 1.0 × 10−4 | 1.4 × 10−2 | −0.7768 |
| Ruminococcus (bromii/unclassified) | Rankin | species | 1.5 × 10−4 | 1.6 × 10−2 | −0.9269 |
| (unclassified/Clostridiaceae/Lachnospiraceae)_u | Rankin | genus | 5.8 × 10−7 | 2.5 × 10−4 | −1.1043 |
| (Coprococcus/unclassified) | Rankin | genus | 1.2 × 10−4 | 1.5 × 10−2 | −0.7815 |
| Roseburia | Rankin | genus | 2.1 × 10−4 | 2.1 × 10−2 | −0.7667 |
| (unclassified/Clostridiaceae/Lachnospiraceae) | Rankin | family | 7.6 × 10−6 | 2.5 × 10−4 | −1.2307 |
| PhPA | Rankin | - | 1.0 × 10−3 | 3.4 × 10−2 | −1.40 |
| PhPA | GCS | - | 7.6 × 10−4 | 3.4 × 10−2 | 0.65 |
| Metabolite | Bacteria | Adjusted R2 of the Balance-Metabolite Model | Direction | Reproducibility of the Taxon |
|---|---|---|---|---|
| PhPA | Coprococcus_u | 0.35593 | + | 78 |
| S100 | Lactococcus_u | 0.20522 | + | 70 |
| Succinic acid | Bifidobacterium_u | 0.31986 | + | 60 |
| Taxon | Factor | Rank | P Value | FDR | Estimate |
|---|---|---|---|---|---|
| (unclassified/Clostridium)_u | antibiotics | species | 1.2 × 10−4 | 0.0264 | −1.77 |
| (unclassified/Clostridium) | antibiotics | genus | 1.0 × 10−4 | 0.0264 | −1.79 |
| Ruminococcus (bromii/unclassified) | nutrition type | species | 3.0 × 10−4 | 0.0443 | 1.90 |
| [Eubacterium]_dolichum | nutrition type | species | 1.3 × 10−4 | 0.0264 | −1.89 |
| Parameters | Group with Positive Dynamics (n = 15) | Group with Negative Dynamics (n = 29) |
|---|---|---|
| Age | 48 (29–59) | 53 (39–64) |
| Number of patients taking antibiotics | 6/15 | 15/29 |
| Nutrition type | 7/15 | 16/29 |
| NIHSS | 15 (11–19) | 18 (13–24) |
| Rivermead Mobility Index | 1 (0–8) | 2 (0–3) |
| Rankin Scale | 5 (4–5) | 5 (4–5) |
| GCS | 15 (14–15) | 15 (13–15) |
| FOUR | 14 (14–16) | 14 (10–16) |
| PCT (0.25 ng/mL) | 0.03 (0.02–0.06) | 0.06 (0.03–0.1) |
| Sum of PhCA | 3.2 (2.3–6.4) | 4.1 (3.1–5.3) |
| S100 protein | 0.05 (0.03–0.08) | 0.06 (0.04–0.09) |
| IL-6 | 14.3 (12.8–27.6) | 18.2 (12.9–31.9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernevskaya, E.; Klimenko, N.; Pautova, A.; Buyakova, I.; Tyakht, A.; Beloborodova, N. Host-Microbiome Interactions Mediated by Phenolic Metabolites in Chronically Critically Ill Patients. Metabolites 2021, 11, 122. https://doi.org/10.3390/metabo11020122
Chernevskaya E, Klimenko N, Pautova A, Buyakova I, Tyakht A, Beloborodova N. Host-Microbiome Interactions Mediated by Phenolic Metabolites in Chronically Critically Ill Patients. Metabolites. 2021; 11(2):122. https://doi.org/10.3390/metabo11020122
Chicago/Turabian StyleChernevskaya, Ekaterina, Natalia Klimenko, Alisa Pautova, Irina Buyakova, Alexander Tyakht, and Natalia Beloborodova. 2021. "Host-Microbiome Interactions Mediated by Phenolic Metabolites in Chronically Critically Ill Patients" Metabolites 11, no. 2: 122. https://doi.org/10.3390/metabo11020122
APA StyleChernevskaya, E., Klimenko, N., Pautova, A., Buyakova, I., Tyakht, A., & Beloborodova, N. (2021). Host-Microbiome Interactions Mediated by Phenolic Metabolites in Chronically Critically Ill Patients. Metabolites, 11(2), 122. https://doi.org/10.3390/metabo11020122