
Characterizing the Flavor Precursors and Liberation Mechanisms of Various Dry-Aging Methods in Cull Beef Loins Using Metabolomics and Microbiome Approaches

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Free Amino Acid and Sugar Concentration
2.2. Free Fatty Acid and Volatile Content Analysis
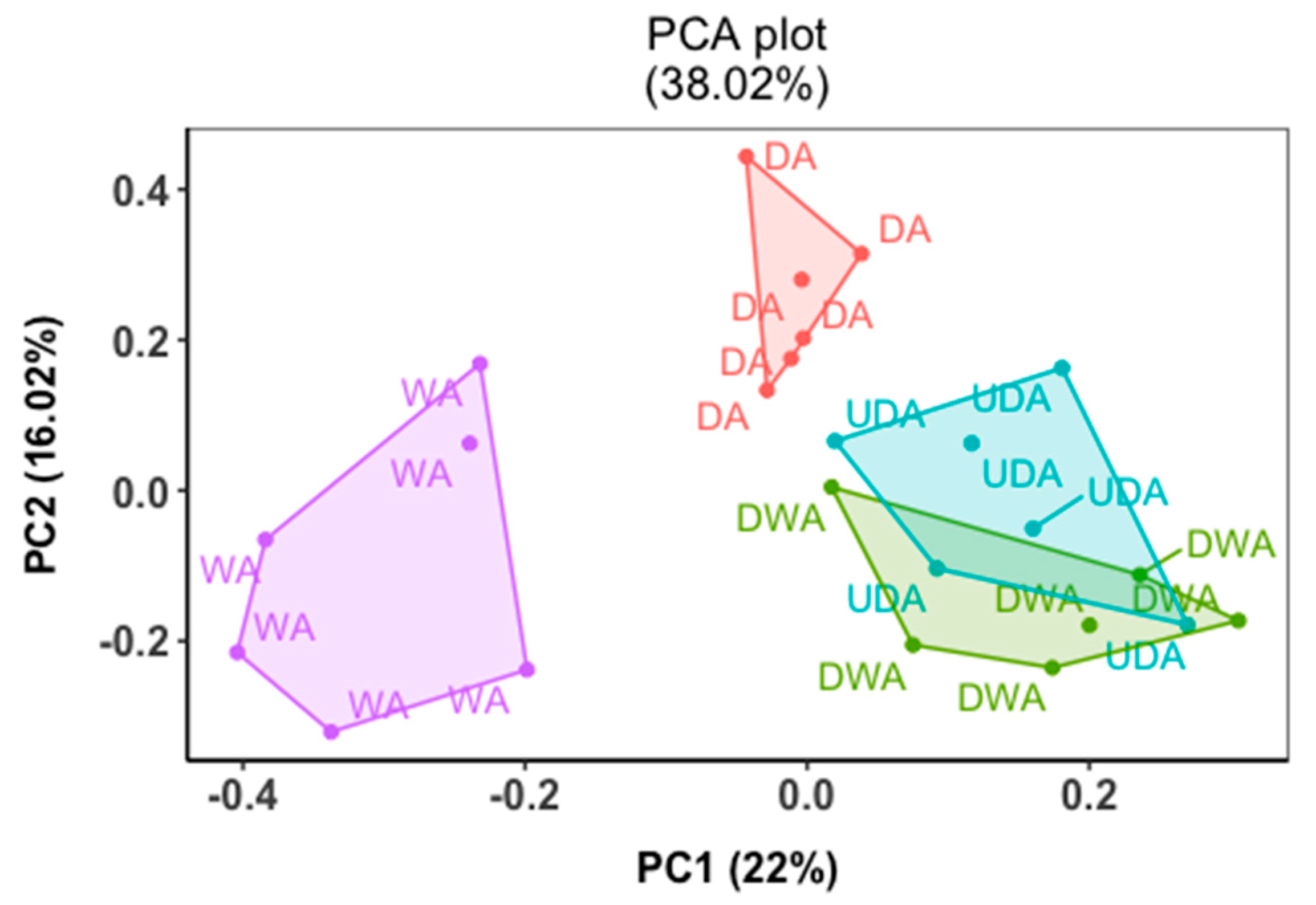
2.3. Metabolomics Analysis
2.4. Microbiome Analysis
2.4.1. Sequence Quality and Contamination
2.4.2. Diversity Measures
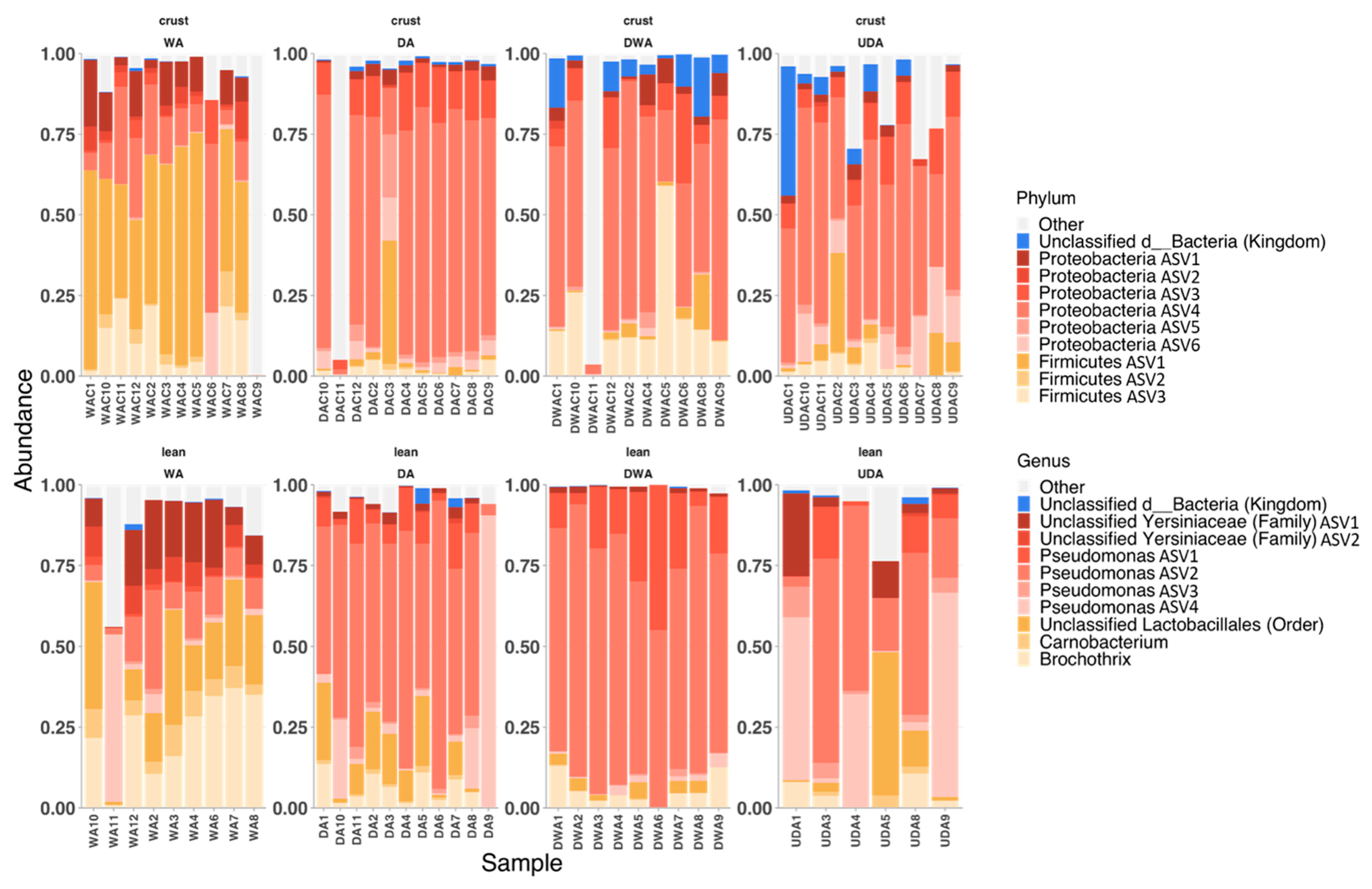
2.4.3. Relative Abundances, Microbial Markers, and Co-Occurrence
3. Discussion
3.1. Flavor Precursors and Flavor Generation
3.2. Flavor Precursors Generation Mechanism
3.2.1. Dehydration
3.2.2. Microbial Involvement
4. Materials and Methods
4.1. Sample Collection, Preparation, and Processing
4.2. Free Amino Acid Analysis
4.3. Sugar Content Analysis
4.4. Free Fatty Acid Analysis
4.5. Volatile Compound Analysis
4.6. Metabolomics Profiling
4.6.1. Sample Extraction
4.6.2. Ultra-Performance Liquid Chromatography–Mass Spectrometry Analysis (UPLC_MS)
4.7. Microbiome Analysis
4.7.1. Sample Preparation and DNA Extraction
4.7.2. 16S Library Preparation and Sequencing
4.7.3. Bioinformatics Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dashdorj, D.; Tripathi, V.K.; Cho, S.; Kim, Y.; Hwang, I. Dry Aging of Beef; Review. J. Anim. Sci. Technol. 2016, 58, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.B.; Kemp, R.; Samuelsson, L.M. Effects of Dry-Aging on Meat Quality Attributes and Metabolite Profiles of Beef Loins. Meat Sci. 2016, 111, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Kim, Y.H.B.; Cooper, B.; Oh, J.-H.; Chun, H.; Choe, J.-H.; Schoonmaker, J.P.; Ajuwon, K.; Min, B. Metabolomics Profiling to Determine the Effect of Postmortem Aging on Color and Lipid Oxidative Stabilities of Different Bovine Muscles. J. Agric. Food Chem. 2017, 65, 6708–6716. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Dillwith, J.W.; Mafi, G.G.; VanOverbeke, D.L.; Ramanathan, R. Metabolite Profile Differences between Beef Longissimus and Psoas Muscles during Display. Meat Muscle Biol. 2017, 1, 18. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Suh, D.H.; Zhang, J.; Chao, Y.; Duttlinger, A.W.; Johnson, J.S.; Lee, C.H.; Kim, Y.H.B. Elucidating the Involvement of Apoptosis in Postmortem Proteolysis in Porcine Muscles from Two Production Cycles Using Metabolomics Approach. Sci. Rep. 2021, 11, 3465. [Google Scholar] [CrossRef]
- Setyabrata, D.; Cooper, B.R.; Sobreira, T.J.P.; Legako, J.F.; Martini, S.; Kim, Y.H.B. Elucidating Mechanisms Involved in Flavor Generation of Dry-Aged Beef Loins Using Metabolomics Approach. Food Res. Int. 2021, 139, 109969. [Google Scholar] [CrossRef]
- Lee, H.J.; Yoon, J.W.; Kim, M.; Oh, H.; Yoon, Y.; Jo, C. Changes in Microbial Composition on the Crust by Different Air Flow Velocities and Their Effect on Sensory Properties of Dry-Aged Beef. Meat Sci. 2019, 153, 152–158. [Google Scholar] [CrossRef]
- Setyabrata, D.; Xue, S.; Vierck, K.; Legako, J.; Ebner, P.; Zuelly, S.; Kim, Y.H.B.; Setyabrata, D.; Xue, S.; Vierck, K.; et al. Impact of Various Dry-Aging Methods on Meat Quality and Palatability Attributes of Beef Loins (M. Longissimus Lumborum) from Cull Cow. Meat Muscle Biol. 2022, 6, 13025. [Google Scholar] [CrossRef]
- Dashdorj, D.; Amna, T.; Hwang, I. Influence of Specific Taste-Active Components on Meat Flavor as Affected by Intrinsic and Extrinsic Factors: An Overview. Eur. Food Res. Technol. 2015, 241, 157–171. [Google Scholar] [CrossRef]
- Mungure, T.E.; Farouk, M.M.; Birch, E.J.; Carne, A.; Staincliffe, M.; Stewart, I.; Bekhit, A.E.D.A. Effect of PEF Treatment on Meat Quality Attributes, Ultrastructure and Metabolite Profiles of Wet and Dry Aged Venison Longissimus Dorsi Muscle. Innov. Food Sci. Emerg. Technol. 2020, 65, 102457. [Google Scholar] [CrossRef]
- Lee, H.J.; Choe, J.; Kim, M.; Kim, H.C.; Yoon, J.W.; Oh, S.W.; Jo, C. Role of Moisture Evaporation in the Taste Attributes of Dry- and Wet-Aged Beef Determined by Chemical and Electronic Tongue Analyses. Meat Sci. 2019, 151, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Calkins, C.R.; Hodgen, J.M. A Fresh Look at Meat Flavor. Meat Sci. 2007, 77, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, V.; Song, Z.; Hwang, I. The Potential Role of Secondary Metabolites in Modulating the Flavor and Taste of the Meat. Food Res. Int. 2019, 122, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Liu, J.; Song, H.; Liu, Y. Discovery of Amadori-Type Conjugates in a Peptide Maillard Reaction and Their Corresponding Influence on the Formation of Pyrazines. J. Food Sci. 2018, 83, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of Taste-Active Amino Acids, Amino Acid Derivatives and Peptides in Food Fermentations—A Review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Foraker, B.A.; Woerner, D.R.; Gredell, D.; Tatum, J.D.; Legako, J.F.; Belk, K.E.; Stevens, R.D. Flavor, Tenderness, and Related Chemical Changes of Aged Beef Strip Loins. Meat Muscle Biol. 2020, 4, 28. [Google Scholar] [CrossRef]
- Baek, H.H. Process Flavors. In Handbook of Meat, Poultry and Seafood Quality; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 151–162. ISBN 081382446X. [Google Scholar]
- Koutsidis, G.; Elmore, J.S.; Oruna-Concha, M.J.; Campo, M.M.; Wood, J.D.; Mottram, D.S. Water-Soluble Precursors of Beef Flavour. Part II: Effect of Post-Mortem Conditioning. Meat Sci. 2008, 79, 270–277. [Google Scholar] [CrossRef]
- Dinh, T.T.N.; Legako, J.F.; Miller, M.F.; Brooks, J.C. Effects of USDA Quality Grade and Cooking on Water-Soluble Precursors of Beef Flavor. Meat Sci. 2018, 146, 122–130. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeon, M.Y.; Lee, C.H. Physicochemical and Sensory Characteristics of Commercial, Frozen, Dry, and Wet-Aged Hanwoo Sirloins. Asian-Australas. J. Anim. Sci. 2019, 32, 1621–1629. [Google Scholar] [CrossRef]
- Min, B.; Ahn, D.U. Mechanism of Lipid Peroxidation in Meat and Meat Products -A Review. Food Sci. Biotechnol. 2005, 14, 152–163. [Google Scholar]
- Mottram, D.S. Flavour Formation in Meat and Meat Products: A Review. Food Chem. 1998, 62, 415–424. [Google Scholar] [CrossRef]
- Whitfield, F.B.; Mottram, D.S. Volatiles from Interactions of Maillard Reactions and Lipids. Crit. Rev. Food Sci. Nutr. 1992, 31, 58. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.T.N.; To, K.V.; Schilling, M.W. Fatty Acid Composition of Meat Animals as Flavor Precursors. Meat Muscle Biol. 2021, 5, 34. [Google Scholar] [CrossRef]
- Ribeiro, F.A.; Lau, S.K.; Furbeck, R.A.; Herrera, N.J.; Henriott, M.L.; Bland, N.A.; Fernando, S.C.; Subbiah, J.; Sullivan, G.A.; Calkins, C.R. Ultimate PH Effects on Dry-Aged Beef Quality. Meat Sci. 2021, 172, 309–1740. [Google Scholar] [CrossRef] [PubMed]
- Capouya, R.; Mitchell, T.; Clark, D.L.; Clark, D.I.; Bass, P. A Survey of Microbial Communities on Dry-Aged Beef in Commercial Meat Processing Facilities. Meat Muscle Biol. 2020, 4, 5–6. [Google Scholar] [CrossRef]
- Pothakos, V.; Devlieghere, F.; Villani, F.; Björkroth, J.; Ercolini, D. Lactic Acid Bacteria and Their Controversial Role in Fresh Meat Spoilage. Meat Sci. 2015, 109, 66–74. [Google Scholar] [CrossRef]
- Casaburi, A.; Piombino, P.; Nychas, G.J.; Villani, F.; Ercolini, D. Bacterial Populations and the Volatilome Associated to Meat Spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef]
- Cangelosi, G.A.; Meschke, J.S. Dead or Alive: Molecular Assessment of Microbial Viability. Appl. Environ. Microbiol. 2014, 80, 5884–5891. [Google Scholar] [CrossRef] [Green Version]
- Vierck, K.R.; Legako, J.F.; Kim, J.K.; Johnson, B.J.; Brooks, J.C. Determination of Package and Muscle-Type Influence on Proteolysis, Beef-Flavor-Contributing Free Amino Acids, Final Beef Flavor, and Tenderness. Meat Muscle Biol. 2020, 4, 1–14. [Google Scholar] [CrossRef]
- Chail, A.; Legako, J.F.; Pitcher, L.R.; Griggs, T.C.; Ward, R.E.; Martini, S.; MacAdam, J.W. Legume Finishing Provides Beef with Positive Human Dietary Fatty Acid Ratios and Consumer Preference Comparable with Grain-Finished Beef1. J. Anim. Sci. 2016, 94, 2184–2197. [Google Scholar] [CrossRef] [Green Version]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A Direct Method for Fatty Acid Methyl Ester Synthesis: Application to Wet Meat Tissues, Oils, and Feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, K.; Legako, J.F. Volatile Flavor Compounds Vary by Beef Product Type and Degree of Doneness. J. Anim. Sci. 2018, 96, 4238–4250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the Miseq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan” Title Community Ecology Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 17 April 2022).



{kind=link}
{kind=link}
| Free Amino Acids (mmol/Kg of Wet Meat) | WA | DA | DWA | UDA | SEM | p-Value |
|---|---|---|---|---|---|---|
| Alanine | 3.899 b | 6.477 a | 5.730 a | 5.601 a | 0.345 | <0.0001 |
| Asparagine | 0.218 c | 0.323 ab | 0.348 a | 0.269 bc | 0.027 | 0.0021 |
| Aspartate | 0.103 | 0.153 | 0.165 | 0.145 | 0.022 | 0.0622 |
| Beta-Alanine | 0.241 b | 0.349 a | 0.315 ab | 0.386 a | 0.040 | 0.0498 |
| Cysteine | 1.056 b | 1.578 a | 1.685 a | 1.398 a | 0.129 | 0.0008 |
| Cystine | 0.015 | 0.016 | 0.016 | 0.014 | 0.003 | 0.9738 |
| Glutamate | 0.920 c | 1.586 b | 2.278 a | 1.287 b | 0.139 | <0.0001 |
| Glutamine | 0.002 c | 0.012 a | 0.011 a | 0.006 b | 0.001 | <0.0001 |
| Glycine | 1.329 b | 2.153 a | 1.961 a | 1.859 a | 0.140 | 0.0004 |
| Histidine | 1.050 b | 2.773 a | 3.675 a | 3.164 a | 0.352 | <0.0001 |
| Hydroxyproline | 0.041 | 0.047 | 0.048 | 0.057 | 0.005 | 0.1702 |
| Isoleucine | 0.846 b | 1.134 a | 1.130 a | 1.059 ab | 0.095 | 0.0415 |
| Leucine | 1.065 b | 1.396 a | 1.256 ab | 1.340 a | 0.103 | 0.0462 |
| Lysine | 0.477 c | 1.353 a | 1.308 a | 1.002 b | 0.115 | <0.0001 |
| Methionine | 0.379 b | 0.606 a | 0.523 a | 0.520 a | 0.052 | 0.0048 |
| Ornithine | 0.048 b | 0.118 a | 0.110 a | 0.083 ab | 0.013 | 0.0017 |
| Phenyl Alanine | 0.508 b | 0.749 a | 0.721 a | 0.720 a | 0.062 | 0.0057 |
| Proline | 0.356 b | 0.483 a | 0.476 a | 0.415 ab | 0.033 | 0.0206 |
| Serine | 1.336 c | 2.070 b | 2.726 a | 2.029 b | 0.213 | 0.0001 |
| Threonine | 0.767 c | 1.159 ab | 1.394 a | 1.113 b | 0.103 | 0.0007 |
| Tyrosine | 0.389 b | 0.840 a | 0.825 a | 0.769 a | 0.074 | <0.0001 |
| Tyrptophan | 0.047 b | 0.102 a | 0.087 a | 0.083 a | 0.009 | 0.0004 |
| Valine | 1.409 b | 1.875 a | 1.865 a | 1.729 ab | 0.148 | 0.0454 |
| Total Free Amino Acid | 16.308 b | 27.351 a | 28.652 a | 24.522 a | 1.806 | <0.0001 |
| Total Free Amino Acid Dry Basis (mmol/Kg dry meat) | 50.663 c | 69.480 b | 85.513 a | 53.074 c | 5.226 | <0.0001 |
| Reducing Sugars (mmol/Kg of Wet Meat) | WA | DA | DWA | UDA | SEM | p-Value |
|---|---|---|---|---|---|---|
| Ribose | 0.5173 b | 0.7881 a | 0.6672 ab | 0.6808 ab | 0.0690 | 0.0418 |
| Fructose | 0.8581 | 1.3355 | 1.1871 | 1.1818 | 0.1743 | 0.2260 |
| Mannose | 1.8872 | 2.7303 | 2.7838 | 2.5197 | 0.2789 | 0.0734 |
| Glucose | 11.1989 b | 19.4053 a | 16.1550 a | 16.4701 a | 1.4932 | 0.0017 |
| Myoinositol | 0.3815 c | 0.6911 a | 0.6224 ab | 0.4802 bc | 0.0703 | 0.0046 |
| Ribose 5-phosphate | 0.0467 | 0.0875 | 0.1674 | 0.0680 | 0.0343 | 0.0771 |
| Fructose 6-phosphate | 1.6241 | 2.0178 | 2.7800 | 1.7763 | 0.5367 | 0.4410 |
| Mannose 6-phosphate | 0.9645 | 1.4169 | 1.4313 | 1.4415 | 0.3009 | 0.5632 |
| Glucose 6-phosphate | 7.2096 | 13.1020 | 10.3266 | 8.7132 | 1.8726 | 0.1276 |
| Maltose | 0.0492 | 0.0603 | 0.0438 | 0.0468 | 0.0152 | 0.8184 |
| Total Sugars | 24.7371 b | 41.6349 a | 36.1647 a | 33.3785 ab | 3.8001 | 0.0222 |
| Total Reducing Sugars | 24.3556 b | 40.9438 a | 35.5423 a | 32.8983 ab | 3.7877 | 0.0251 |
| Total Sugars Dry Basis (mmol/Kg of dry meat) | 78.9235 | 105.82 | 108.33 | 73.6774 | 11.2584 | 0.0535 |
| Volatile Compounds Name (ng/g Sample) | WA | DA | DWA | UDA | SEM | p-Value |
|---|---|---|---|---|---|---|
| n-aldehydes | ||||||
| Acetaldehyde | 9.12 c | 19.22 c | 53.58 a | 36.99 b | 5.18 | <0.0001 |
| Butanal | 4.67 c | 22.02 b | 44.26 a | 44.04 a | 5.54 | <0.0001 |
| Heptanal | 8.99 | 8.70 | 17.06 | 7.72 | 3.13 | 0.1397 |
| Hexanal | 119.12 | 79.55 | 175.56 | 88.48 | 48.16 | 0.4940 |
| Nonanal | 6.74 ab | 4.4 b | 10.96 a | 2.72 b | 1.93 | 0.0245 |
| Octanal | 1.69 b | 2.32 b | 4.48 a | 2.16 b | 0.44 | 0.0002 |
| Pentanal | 40.86 | 3.41 | 8.28 | 4.21 | 19.87 | 0.4931 |
| Strecker aldehydes | ||||||
| 2-methylbutanal | 8.00 c | 49.59 b | 110.07 a | 106.20 a | 16.23 | <0.0001 |
| 3-methylbutanal | 10.29 c | 67.42 b | 144.84 a | 143.90 a | 21.82 | <0.0001 |
| Benzaldehyde | 27.55 | 19.14 | 23.35 | 21.99 | 5.26 | 0.7209 |
| Phenylacetaldehyde | 2.94 | 1.74 | 2.31 | 1.45 | 0.56 | 0.2062 |
| Alcohols | ||||||
| 1-Hexanol | 1.11 c | 3.54 b | 5.60 a | 3.78 ab | 0.76 | 0.0007 |
| 1-Octanol | 2.90 b | 2.38 b | 6.12 a | 1.06 b | 0.99 | 0.0061 |
| 1-Octen-3-ol | 2.68 b | 5.63 ab | 7.74 a | 5.70 ab | 1.28 | 0.0262 |
| 1-Pentanol | 12.83 | 14.89 | 23.77 | 26.65 | 6.20 | 0.3014 |
| 1-penten-3-ol | 0.10 b | 0.64 ab | 1.12 a | 0.98 a | 0.21 | 0.0043 |
| Ethanol | 59.84 | 89.84 | 277.57 | 136.94 | 59.87 | 0.0635 |
| Ketone | ||||||
| 2,3-butanedione | 8.17 b | 54.37 a | 70.71 a | 65.68 a | 15.03 | 0.0198 |
| 2,3-pentanedione | 0.04 b | 0.14 a | 0.19 a | 0.14 a | 0.02 | 0.0002 |
| 2-heptanone | 1.69 | 2.25 | 2.89 | 2.44 | 0.48 | 0.3274 |
| 2-pentanone | 0.41 c | 1.28 bc | 2.79 a | 1.79 ab | 0.35 | 0.0002 |
| 2-Propanone | 42.12 b | 63.01 b | 131.49 a | 114.52 a | 13.86 | <0.0001 |
| 3-hydroxy-2-butanone | 10.88 b | 60.72 ab | 169.82 a | 133.57 a | 39.62 | 0.0223 |
| Hydrocarbon | ||||||
| Alpha-pinene | 0.00 c | 0.13 cb | 0.35 a | 0.25 ab | 0.05 | <0.0001 |
| Benzene | 0.99 b | 2.07 a | 2.58 a | 2.65 a | 0.31 | 0.0011 |
| D-limonene | 10.67 b | 30.01 a | 37.29 a | 30.31 a | 3.81 | <0.0001 |
| Ethyl benzene | 0.37 b | 0.86 a | 1.06 a | 0.79 a | 0.13 | 0.0025 |
| p-Xylene | 0.97 b | 2.07 ab | 2.96 a | 2.22 a | 0.41 | 0.0083 |
| Styrene | 1.24 b | 2.38 a | 2.90 a | 2.46 a | 0.31 | 0.0028 |
| Toluene | 6.59 b | 17.03 a | 21.87 a | 20.13 a | 2.15 | <0.0001 |
| Octane | 2.13 c | 6.13 b | 10.71 a | 7.96 ab | 1.02 | <0.0001 |
| Pentane | 3.85 b | 5.46 b | 13.48 a | 10.56 a | 1.63 | 0.0003 |
| Pyrazine | ||||||
| 2,5-dimethylpyrazine | 5.69 | 4.89 | 6.64 | 5.25 | 2.04 | 0.9229 |
| 2-ethyl-3,5/6-dimethylpyrazine | 1.48 | 1.41 | 2.08 | 1.56 | 0.45 | 0.6246 |
| Methyl-pyrazine | 0.97 b | 2.77 a | 3.91 a | 3.26 a | 0.78 | 0.0107 |
| Trimethylpyrazine | 1.08 | 1.30 | 2.35 | 1.58 | 0.46 | 0.1949 |
| Furans | ||||||
| 2-Pentyl furan | 0.92 | 0.45 | 0.69 | 0.38 | 0.28 | 0.5207 |
| Lactone | ||||||
| Butyrolactone | 2.09 b | 15.94 a | 24.46 a | 18.11 a | 3.54 | 0.0004 |
| Sulfur-containing | ||||||
| 2-methyl thiophene | 0.59 | 0.47 | 0.59 | 0.51 | 0.05 | 0.2327 |
| Carbon disulfide | 10.33 b | 12.38 b | 21.11 a | 13.36 b | 2.57 | 0.0232 |
| Dimethyl sulfide | 3.74 c | 7.37 cb | 14.82 a | 10.82 ab | 1.43 | <0.0001 |
| Dimethyl-disulfide | 0.03 | 0.03 | 0.06 | 0.05 | 0.01 | 0.1275 |
| Methanethiol | 2.88 bc | 1.54 c | 5.22 a | 4.12 ab | 0.77 | 0.0107 |
| Methional | 3.17 | 2.36 | 3.58 | 2.50 | 1.20 | 0.8636 |
| Carboxylic acid | ||||||
| Acetic acid | 9.30 c | 11.25 cb | 23.11 a | 18.27 ab | 2.52 | 0.0008 |
| Butanoic acid | 20.30 c | 110.38 b | 202.04 a | 164.57 ab | 26.03 | 0.0001 |
| Butanoic acid, methyl ester | 3.55 | 0.98 | 1.55 | 2.01 | 1.67 | 0.6963 |
| Heptanoic acid, methyl ester | 0.17 | 0.24 | 0.37 | 0.27 | 0.07 | 0.2516 |
| Hexanoic acid, methyl ester | 3.51 | 4.57 | 6.70 | 6.70 | 1.53 | 0.3502 |
| Hexanoic acid, methyl ester | 3.11 | 4.57 | 6.70 | 6.70 | 1.53 | 0.2768 |
| Nonanoic acid, methyl ester | 0.79 | 0.46 | 0.53 | 0.34 | 0.13 | 0.0861 |
| Octanoic acid, methyl ester | 1.32 | 1.48 | 1.84 | 1.09 | 0.20 | 0.0690 |
| Mass | RT | Highest Abundant | HMDB ID | Putative Name | WA | DA | DWA | UDA |
|---|---|---|---|---|---|---|---|---|
| Protein-derived | ||||||||
| 115.0633 | 1.21 | WA | HMDB0000162 | Proline | 18.32 a | 16.23 b | 16.92 b | 16.74 b |
| 244.1774 | 6.35 | WA | HMDB28910 | Ile-Ile | 18.37 a | 16.21 b | 16.25 b | 16.49 b |
| 357.2623 | 9.86 | WA | HMDB0094648 | Leu-Leu-Leu | 18.02 a | 16.30 b | 16.26 b | 16.65 b |
| 244.1067 | 7.38 | WA/DA | HMDB0028864 | Hyp-Hyp | 17.91 a | 17.83 a | 17.06 b | 17.25 ab |
| 312.1437 | 5.47 | WA | HMDB0131468 | Phe-Phe | 18.07 a | 17.94 ab | 16.98 c | 17.23 bc |
| 239.0794 | 7.21 | DA/WA | HMDB0131468 | Aspartic Acid | 18.11 a | 18.12 a | 17.21 b | 17.53 b |
| 284.1122 | 3.95 | DA/WA | HMDB0028821 | Gln-His | 19.11 a | 19.16 a | 18.19 b | 18.27 b |
| 204.1112 | 2.23 | DA | HMDB0029136 | Val-Ser | 18.11 b | 18.50 a | 17.55 c | 17.59 bc |
| 236.0465 | 4.78 | DA | HMDB0028750 | Asp-Cys | 17.66 ab | 17.96 a | 17.07 b | 17.24 b |
| 133.0196 | 4.78 | DA | HMDB0062164 | Thioproline | 17.25 ab | 17.49 a | 16.62 b | 16.81 b |
| 218.1259 | 4.01 | DA/DWA/UDA | HMDB0029042 | Ser-Ile | 20.92 b | 21.69 a | 21.40 a | 21.51 a |
| 284.11 | 1.71 | DA | HMDB0028884 | His-Glu | 22.40 ab | 22.73 a | 21.74 b | 21.72 b |
| 174.1032 | 1.43 | DA | HMDB0028854 | Theanine | 17.97 b | 18.33 a | 17.84 b | 17.66 b |
| 188.1165 | 3.43 | DA/DWA | HMDB0000446 | Acetyl-Lysine | 22.50 b | 23.08 a | 23.08 a | 22.56 ab |
| 127.0632 | 8.22 | DA/WA | HMDB0029434 | Methyleneproline | 17.53 a | 17.76 a | 16.83 b | 17.25 ab |
| 115.0634 | 0.86 | DWA/WA | HMDB0000162 | Proline | 19.35 a | 19.10 b | 19.36 a | 18.85 b |
| 257.1022 | 0.82 | DWA/WA | HMDB0039229 | Gln-Gln | 17.44 a | 16.99 b | 17.67 a | 17.13 ab |
| 155.0693 | 3.26 | UDA/DWA/DA | HMDB0000177 | Histidine | 17.42 b | 17.99 a | 18.10 a | 18.32 a |
| 343.1257 | 1.85 | UDA/DWA/DA | HMDB0037845 | Deoxyfructosyl Tyrosine | 20.48 b | 20.36 a | 20.86 a | 20.96 a |
| Carbohydrates-derived | ||||||||
| 464.2283 | 12.43 | WA | HMDB0031367 | Linalooloxide apiosylglucoside | 21.06 a | 20.74 ab | 19.64 c | 20.21 bc |
| 284.1211 | 0.72 | DA | HMDB0029819 | Phenylethyl glucopyranoside | 17.82 b | 18.28 a | 18.15 b | 18.19 ab |
| 379.1063 | 3.34 | UDA/DA | HMDB0001066 | Lactoylglutathione | 19.40 b | 19.86 a | 19.68 ab | 20.22 a |
| Lipids-derived | ||||||||
| 565.4201 | 16.36 | WA | HMDB0011497 | Lysophosphatidylethanolamine | 20.43 a | 19.18 b | 19.25 b | 19.47 b |
| 452.3357 | 14.23 | WA/DWA/UDA | HMDB0037065 | Oxoursadienoate | 20.29 a | 18.96 b | 19.09 a | 19.42 a |
| 452.3361 | 14.04 | WA | HMDB0035888 | Tyromycic acid | 21.83 a | 20.82 b | 20.82 b | 21.13 ab |
| 284.1073 | 1.56 | WA/DA | HMDB0030694 | Demethoxymatteucinol | 21.66 a | 21.56 a | 20.89 b | 20.88 b |
| 232.1129 | 2.2 | WA | HMDB0036189 | Tetrahydrofurfuryl cinnamate | 19.17 a | 18.81 ab | 18.43 b | 18.69 b |
| 286.1532 | 16.3 | DA | HMDB0060085 | Estradiol quinone | 19.62 ab | 19.63 a | 18.64 c | 19.00 bc |
| 266.1728 | 12.08 | UDA/DWA/DA | HMDB0030356 | Didehydrocondyfolan | 16.30 b | 17.32 a | 17.79 a | 18.04 a |
| 407.0982 | 4.11 | UDA | HMDB0030257 | Erysothiopine | 19.98 ab | 19.46 b | 20.00 ab | 20.33 a |
| 132.0946 | 1.59 | UDA/DWA | HMDB0029641 | Cymenene | 22.03 b | 22.06 b | 22.28 a | 22.29 a |
| Organic acids | ||||||||
| 365.0897 | 4.89 | DA | HMDB0062198 | Glutathionyl acetate | 21.88 b | 22.24 a | 21.23 b | 21.42 b |
| 298.1283 | 3.01 | DA/WA | HMDB06101 | Enterolactone | 17.90 a | 17.95 a | 16.93 b | 17.06 ab |
| 276.1212 | 3.05 | DWA/DA | HMDB0034263 | Triethyl citrate | 22.12 b | 22.46 a | 22.51 a | 22.27 ab |
| 118.0277 | 3.59 | UDA/DA/WA | HMDB0031204 | Hydroxyoxobutanoic acid | 18.57 a | 18.63 a | 18.44 b | 18.65 a |
| 164.0469 | 2.34 | UDA | HMDB0001713 | Coumaric acid | 21.85 b | 21.77 b | 21.32 c | 22.13 a |
| Other | ||||||||
| 113.0843 | 6.55 | WA | HMDB0031199 | Trimethyloxazoline | 21.64 a | 20.32 b | 21.13 ab | 20.32 b |
| 301.1637 | 4.59 | WA/DA | HMDB0032654 | Futoamide | 19.15 a | 19.10 a | 18.43 b | 18.59 ab |
| 194.1156 | 5.62 | DA/DWA/UDA | HMDB0094708 | Tetraethylene glycol | 22.17 b | 22.74 a | 22.56 a | 22.69 a |
| 132.0949 | 1.6 | DWA | HMDB0032303 | Heptanethiol | 22.96 b | 23.12 a | 23.19 b | 23.11 b |
| 94.0395 | 1.53 | DWA | HMDB0000228 | Phenol | 18.66 b | 18.76 b | 18.96 a | 18.71 b |
| 327.1884 | 4.55 | UDA | HMDB0038645 | Piperamide | 20.47 ab | 19.89 b | 20.74 ab | 21.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Setyabrata, D.; Vierck, K.; Sheets, T.R.; Legako, J.F.; Cooper, B.R.; Johnson, T.A.; Kim, Y.H.B. Characterizing the Flavor Precursors and Liberation Mechanisms of Various Dry-Aging Methods in Cull Beef Loins Using Metabolomics and Microbiome Approaches. Metabolites 2022, 12, 472. https://doi.org/10.3390/metabo12060472
Setyabrata D, Vierck K, Sheets TR, Legako JF, Cooper BR, Johnson TA, Kim YHB. Characterizing the Flavor Precursors and Liberation Mechanisms of Various Dry-Aging Methods in Cull Beef Loins Using Metabolomics and Microbiome Approaches. Metabolites. 2022; 12(6):472. https://doi.org/10.3390/metabo12060472
Chicago/Turabian StyleSetyabrata, Derico, Kelly Vierck, Tessa R. Sheets, Jerrad F. Legako, Bruce R. Cooper, Timothy A. Johnson, and Yuan H. Brad Kim. 2022. "Characterizing the Flavor Precursors and Liberation Mechanisms of Various Dry-Aging Methods in Cull Beef Loins Using Metabolomics and Microbiome Approaches" Metabolites 12, no. 6: 472. https://doi.org/10.3390/metabo12060472
APA StyleSetyabrata, D., Vierck, K., Sheets, T. R., Legako, J. F., Cooper, B. R., Johnson, T. A., & Kim, Y. H. B. (2022). Characterizing the Flavor Precursors and Liberation Mechanisms of Various Dry-Aging Methods in Cull Beef Loins Using Metabolomics and Microbiome Approaches. Metabolites, 12(6), 472. https://doi.org/10.3390/metabo12060472