
The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming


Abstract
:1. Introduction
2. The Role of PGPMs in Growth and Productivity
3. The Role of PGPMs in Plant Detoxification
4. The Role of PGPMs in Abiotic Stress Tolerance
4.1. Tolerance to a Drought Environment
4.2. Tolerance to Extreme Temperatures
4.3. Tolerance to a Hypersaline Environment
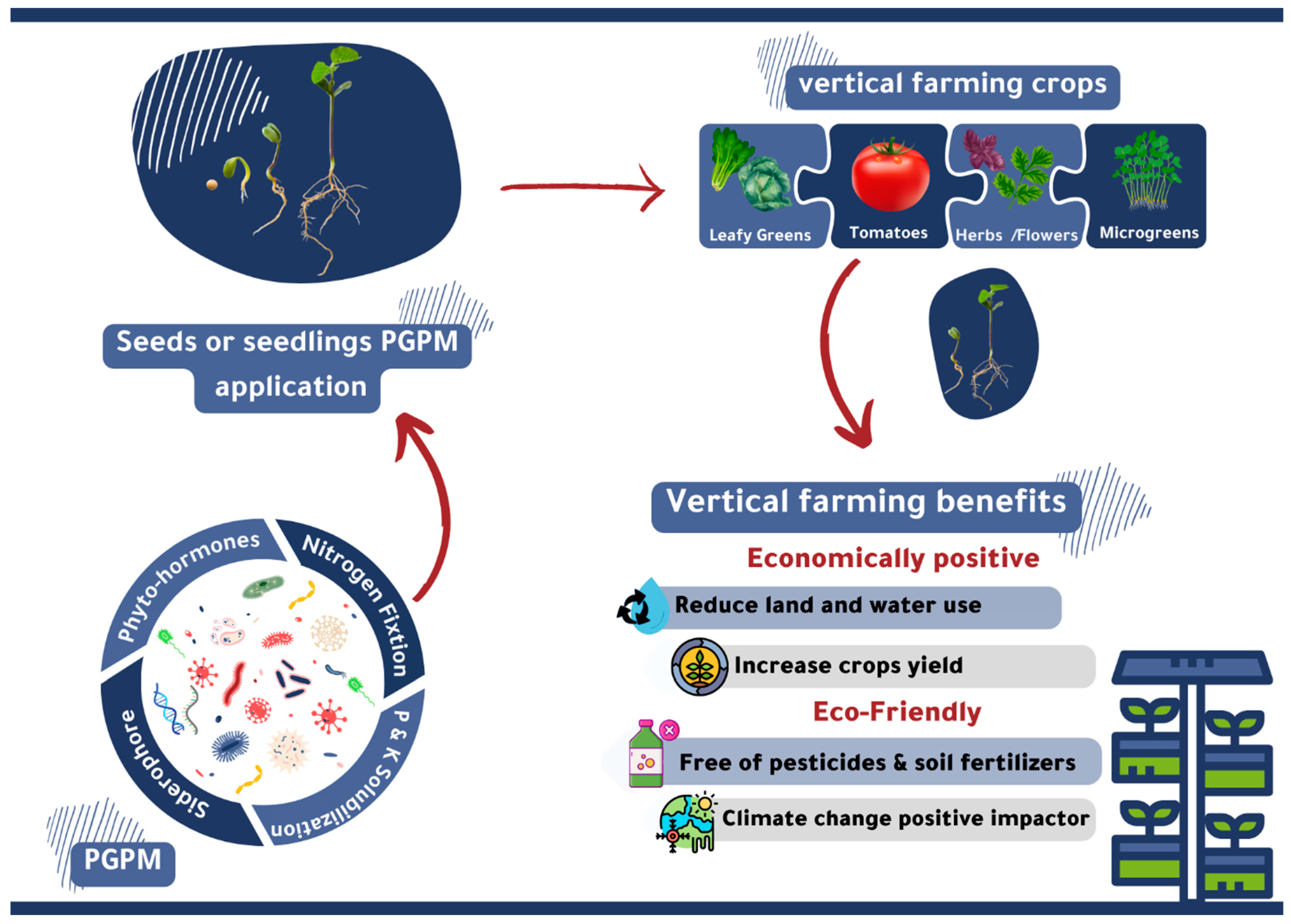
5. Possibilities of Using PGPMs in Hydroponics and Vertical Farming
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, Y. Biotechnological potential of plant-microbe interactions in environmental decontamination. Front. Plant Sci. 2019, 10, 1519. [Google Scholar]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Mycorrhiza and heavy metal resistant bacteria enhance growth, nutrient uptake and alter metabolic profile of sorghum grown in marginal soil. Chemosphere 2016, 157, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Proteomics provides insights into biological pathways altered by plant growth promoting bacteria and arbuscular mycorrhiza in sorghum grown in marginal soil. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Dhawi, F.; Hess, A. Plant growth-prompting bacteria influenced metabolites of Zea mays var. amylacea and Pennisetum americanum p. in a species-specific manner. Adv. Biol. Chem. 2017, 7, 161. [Google Scholar] [CrossRef]
- Dhawi, F.; Hess, A. Poor-soil rhizosphere enriched with different microbial activities influence the availability of base elements. Open J. Ecol. 2017, 7, 495–502. [Google Scholar] [CrossRef]
- Dhawi, F.; Alsanie, S.I. So it is above, so it is below: Microbial Pathways Associated with Date Palm Trees. Ann. Rom. Soc. Cell Biol. 2021, 25, 2725–2740. [Google Scholar]
- Dhawi, F. Investigating Two Date Palm Cultivars Microbial Pathways. J. Hunan Univ. Nat. Sci. 2022, 49, 158–160. [Google Scholar]
- Dhawi, F.; Alsanie, S.I. Investigation of Microbial Community Structure and Diversity in the Rhizosphere of Date Palm (Phoenix dactylifera L.), Sukkari Cultivar. J. Hunan Univ. Nat. Sci. 2022, 4, 9–24. [Google Scholar]
- Mehmood, T.; Gaurav, G.K.; Cheng, L.; Klemeš, J.J.; Usman, M.; Bokhari, A.; Lu, J. A review on plant-microbial interactions, functions, mechanisms and emerging trends in bioretention system to improve multi-contaminated stormwater treatment. J. Environ. Manag. 2021, 294, 113108. [Google Scholar] [CrossRef]
- Conlon, R.; Wang, M.; Germaine, X.L.; Mali, R.; Dowling, D.; Germaine, K.J. Ecopiling: Beneficial Soil Bacteria, Plants, and Optimized Soil Conditions for Enhanced Remediation of Hydrocarbon Polluted Soil. Good Microbes Med. Food Prod. Biotechnol. Bioremediat. Agric. 2022, 337–347. [Google Scholar] [CrossRef]
- Augusta, A.C.; Bertha, E.E.; Eromosele, A.S. Plant-Microbe Interaction: Prospects and Applications in Sustainable Environmental Management. In Plant Hormones: Recent Advances. New Perspectives and Applications; IntechOpen: London, UK, 2022; p. 43. [Google Scholar]
- Schnitzler, W.H. Urban hydroponics–facts and vision. In SEAVEG 2012 High Value Vegetables in Southeast Asia: Production, Supply and Demand; AVRDC-World Vegetable Center: Tainan, Taiwan, 2013; p. 285. [Google Scholar]
- Despommier, D. Farming up the city: The rise of urban vertical farms. Trends Biotechnol. 2013, 31, 388–389. [Google Scholar]
- Kaur, G.; Chawla, P. All about Vertical Farming: A Review. Turk. J. Comput. Math. Educ. 2021, 12, 1–14. [Google Scholar]
- Kulak, M.; Graves, A.; Chatterton, J. Reducing greenhouse gas emissions with urban agriculture: A Life Cycle Assessment perspective. Landsc. Urban Plan 2013, 111, 68–78. [Google Scholar] [CrossRef]
- Koriesh, E.M.; Abo-Soud, I.H. Facing Climate Change: Urban Gardening and Sustainable Agriculture. In Climate Change Impacts on Agriculture and Food Security in Egypt; Springer: Cham, Switzerland, 2020; pp. 345–419. [Google Scholar]
- Rajan, P.; Lada, R.R.; MacDonald, M.T. Advancement in indoor vertical farming for microgreen production. Am. J. Plant Sci. 2019, 10, 1397. [Google Scholar]
- Singh, B.K.; Liu, H.; Trivedi, P. Eco-holobiont: A new concept to identify drivers of host-associated microorganisms. Environ. Microbiol. 2020, 22, 564–567. [Google Scholar]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar]
- Kothe, E.; Turnau, K. Mycorrhizosphere communication: Mycorrhizal fungi and endophytic fungus-plant interactions. Front. Microbiol. 2018, 9, 3015. [Google Scholar]
- de la Fuente Cantó, C.; Simonin, M.; King, E.; Moulin, L.; Bennett, M.J.; Castrillo, G.; Laplaze, L. An extended root phenotype: The rhizosphere, its formation and impacts on plant fitness. Plant J. 2020, 103, 951–964. [Google Scholar]
- Dhawi, F. Plant growth promoting Rhizobacteria (PGPR) regulated Phyto and microbial beneficial protein interactions. Open Life Sci. 2020, 15, 68–78. [Google Scholar] [CrossRef]
- Ghitti, E.; Rolli, E.; Crotti, E.; Borin, S. Flavonoids Are Intra-and Inter-Kingdom Modulator Signals. Microorganisms 2022, 10, 2479. [Google Scholar]
- Pratush, A.; Kumar, A.; Hu, Z. Adverse effect of heavy metals (As, Pb, Hg, and Cr) on health and their bioremediation strategies: A review. Int. Microbiol. 2018, 21, 97–106. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed]
- Baset, M.M.; Shamsuddin, Z.H.; Wahab, Z.; Marziah, M. Effect of plant growth promoting rhizobacterial (PGPR) inoculation on growth and nitrogen incorporation of tissue-cultured'musa'plantlets under nitrogen-free hydroponics condition. Aust. J. Crop Sci. 2010, 4, 85–90. [Google Scholar]
- Goswami, M.; Chakraborty, P.; Mukherjee, K.; Mitra, G.; Bhattacharyya, P.; Dey, S.; Tribedi, P. Bioaugmentation and biostimulation: A potential strategy for environmental remediation. J. Microbiol. Exp. 2018, 6, 223–231. [Google Scholar]
- Delaporte-Quintana, P.; Lovaisa, N.C.; Rapisarda, V.A.; Pedraza, R.O. The plant growth promoting bacteria Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production. Plant Growth Regul. 2020, 91, 185–199. [Google Scholar] [CrossRef]
- Phieler, R.; Merten, D.; Roth, M.; Büchel, G.; Kothe, E. Phytoremediation using microbially mediated metal accumulation in Sorghum bicolor. Environ. Sci. Pollut. Res. 2015, 22, 19408–19416. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.C.; Negi, S.; Shukla, P. Engineering PGPMOs through gene editing and systems biology: A solution for phytoremediation? Trends Biotechnol. 2018, 36, 499–510. [Google Scholar] [PubMed]
- Abhilash, P.C.; Powell, J.R.; Singh, H.B.; Singh, B.K. Plant–microbe interactions: Novel applications for exploitation in multipurpose remediation technologies. Trends Biotechnol. 2012, 30, 416–420. [Google Scholar] [PubMed]
- Kuiper, I.; Lagendijk, E.L.; Bloemberg, G.V.; Lugtenberg, B.J. Rhizoremediation: A beneficial plant-microbe interaction. Mol. Plant-Microbe Interact. 2004, 17, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.T.; Waidyarathne, P.; Kloepper, J.; De La Fuente, L. Strain-specific quantification of root colonization by plant growth promoting rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in non-sterile soil and field conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Arnepalli, D.N. Biominerlisation as a remediation technique: A critical review. In Geotechnical Characterisation and Geoenvironmental Engineering; Springer: Singapore, 2019; pp. 155–162. [Google Scholar]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar]
- Lacalle, R.G.; Aparicio, J.D.; Artetxe, U.; Urionabarrenetxea, E.; Polti, M.A.; Soto, M.; Garbisu, C.; Becerril, J.M. Gentle remediation options for soil with mixed chromium (VI) and lindane pollution: Biostimulation, bioaugmentation, phytoremediation and vermiremediation. Heliyon 2020, 6, e04550. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.R.; Smania, A.M.; Fabro, G.; Alvarez, M.E.; Argarana, C.E. Engineering Pseudomonas fluorescens for biodegradation of 2,4-dinitrotoluene. Appl. Environ. Microbiol. 2005, 71, 8864–8872. [Google Scholar]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wan, J.; Wang, J.; Deng, Y.; Liu, Y.; Peng, B. The potential impact on the biodegradation of organic pollutants from composting technology for soil remediation. Waste Manag. 2018, 72, 138–149. [Google Scholar]
- Kumar, A.; Chaturvedi, A.K.; Yadav, K.; Arunkumar, K.P.; Malyan, S.K.; Raja, P.; Kumar, R.; Khan, S.A.; Yadav, K.K.; Rana, K.L.; et al. Fungal phytoremediation of heavy metal-contaminated resources: Current scenario and future prospects. In Recent Advancement in White Biotechnology through Fungi; Springer: Cham, Switzerland, 2019; pp. 437–461. [Google Scholar]
- Bhantana, P.; Rana, M.S.; Sun, X.C.; Moussa, M.G.; Saleem, M.H.; Syaifudin, M.; Shah, A.; Poudel, A.; Pun, A.B.; Bhat, M.A.; et al. Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 2021, 84, 19–37. [Google Scholar]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar]
- Göhre, V.; Paszkowski, U. Contribution of the arbuscular mycorrhizal symbiosis to heavy metal phytoremediation. Planta 2006, 223, 1115–1122. [Google Scholar] [CrossRef] [Green Version]
- Dhawi, F.; Datta, R.; Ramakrishna, W. Metabolomics, biomass and lignocellulosic total sugars analysis in foxtail millet (Setaria italica) inoculated with different combinations of plant growth promoting bacteria and mycorrhiza. Commun. Plant Sci. 2018, 8, 8–14. [Google Scholar]
- Vamerali, T.; Bandiera, M.; Mosca, G. Field crops for phytoremediation of metal-contaminated land. A review. Environ. Chem. Lett. 2010, 8, 1–17. [Google Scholar]
- Chen, B.D.; Li, X.L.; Tao, H.Q.; Christie, P.; Wong, M.H. The role of arbuscular mycorrhiza in zinc uptake by red clover growing in a calcareous soil spiked with various quantities of zinc. Chemosphere 2003, 50, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Joner, E.J.; Leyval, C. Phytoremediation of organic pollutants using mycorrhizal plants: A new aspect of rhizosphere interactions. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 885–894. [Google Scholar]
- Arantza, S.J.; Hiram, M.R.; Erika, K.; Chávez-Avilés, M.N.; Valiente-Banuet, J.I.; Fierros-Romero, G. Bio- and phytoremediation: Plants and microbes to the rescue of heavy metal polluted soils. SN Appl. Sci. 2022, 4, 59. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Prieto-Fernández, A.; Álvarez-Lopez, V.; Monterroso, C.; Cabello-Conejo, M.I.; Acea, M.J.; Kidd, P.S. Nickel solubilizing capacity and characterization of rhizobacteria isolated from hyperaccumulating and non-hyperaccumulating subspecies of Alyssum serpyllifolium. Int. J. Phytoremediat. 2011, 13, 229–244. [Google Scholar] [PubMed]
- Gill, R.A.; Ahmar, S.; Ali, B.; Saleem, M.H.; Khan, M.U.; Zhou, W.; Liu, S. The role of membrane transporters in plant growth and development, and abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 12792. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.E. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2005, 28, 67–77. [Google Scholar]
- Doornbos, R.F.; Geraats, B.P.; Kuramae, E.E.; Van Loon, L.C.; Bakker, P.A. Effects of jasmonic acid, ethylene, and salicylic acid signaling on the rhizosphere bacterial community of Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2011, 24, 395–407. [Google Scholar] [PubMed]
- Osmont, K.S.; Sibout, R.; Hardtke, C.S. Hidden branches: Developments in root system architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef]
- Verbon, E.H.; Liberman, L.M. Beneficial microbes affect endogenous mechanisms controlling root development. Trends Plant science 2016, 21, 218–229. [Google Scholar] [CrossRef]
- Braud, A.; Jézéquel, K.; Bazot, S.; Lebeau, T. Enhanced phytoextraction of an agricultural Cr-and Pb-contaminated soil by bioaugmentation with siderophore-producing bacteria. Chemosphere 2009, 74, 280–286. [Google Scholar] [CrossRef]
- Chang, P.; Gerhardt, K.E.; Huang, X.D.; Yu, X.M.; Glick, B.R.; Gerwing, P.D.; Greenberg, B.M. Plant growth-promoting bacteria facilitate the growth of barley and oats in salt-impacted soil: Implications for phytoremediation of saline soils. Int. J. Phytoremediation 2014, 16, 1133–1147. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.G.; Kim, J.D.; Oh, B.T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard. Mater. 2013, 250, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Agnello, A.C.; Bagard, M.; van Hullebusch, E.D.; Esposito, G.; Huguenot, D. Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci. Total Environ. 2016, 563, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Gu, L.; Guo, C.; Xun, F.; Liu, J. Effect of PGPR Serratia marcescens BC-3 and AMF Glomus intraradices on phytoremediation of petroleum contaminated soil. Ecotoxicology 2014, 23, 674–680. [Google Scholar] [CrossRef]
- DalCorso, G.; Fasani, E.; Manara, A.; Visioli, G.; Furini, A. Heavy metal pollutions: State of the art and innovation in phytoremediation. Int. J. Mol. Sci. 2019, 20, 3412. [Google Scholar] [CrossRef]
- Ma, Y.; Oliveira, R.S.; Freitas, H.; Zhang, C. Biochemical and molecular mechanisms of plant-microbe-metal interactions: Relevance for phytoremediation. Front. Plant Sci. 2016, 7, 918. [Google Scholar]
- Nasfi, Z.; Busch, H.; Kehraus, S.; Linares-Otoya, L.; König, G.M.; Schäberle, T.F.; Bachoual, R. Soil bacteria isolated from tunisian arid areas show promising antimicrobial activities against gram-negatives. Front. Microbiol. 2018, 9, 2742. [Google Scholar]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar]
- Saleem, M.; Arshad, M.; Hussain, S.; Bhatti, A.S. Perspective of plant growth promoting rhizobacteria (PGPR) containing ACC deaminase in stress agriculture. J. Ind. Microbiol. Biotechnol. 2007, 34, 635–648. [Google Scholar]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [PubMed]
- Rai, A.; Borpatragohain, B.; Sahoo, S. Role of plant-microbe interactions on abiotic stress tolerance in plants: A review. Int. J. Agric. Plant Sci. 2019, 1, 25–31. [Google Scholar]
- Ait Barka, E.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [PubMed]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Farrugia, G.; Lurken, M.S.; Bernard, C.E.; Faussone–Pellegrini, M.S.; Smyrk, T.C.; Parkman, H.P.; Abell, T.L.; Snape, W.J.; Hasler, W.L.; et al. Cellular changes in diabetic and idiopathic gastroparesis. Gastroenterology 2011, 140, 1575–1585. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar Ali, S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–4. [Google Scholar] [CrossRef]
- Abobatta, W.F. Drought adaptive mechanisms of plants—A review. Adv. Agric. Environ. Sci. 2019, 2, 62–65. [Google Scholar]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance generated by plant/fungal symbiosis. Science 2002, 298, 1581. [Google Scholar]
- Bui, L.; Luo, H.; Gunther, W.R.; Román-Leshkov, Y. Domino reaction catalyzed by zeolites with Brønsted and Lewis acid sites for the production of γ-valerolactone from furfural. Angew. Chem. 2013, 125, 8180–8183. [Google Scholar]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Wu, J. Advances in K-Means Clustering: A Data Mining Thinking; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar]
- Grover, P.; Sahai, A. Shannon meets Tesla: Wireless information and power transfer. In Proceedings of the 2010 IEEE International Symposium on Information Theory, Austin, TX, USA, 13–18 June 2010; pp. 2363–2367. [Google Scholar]
- Sen, S.; Chandrasekhar, C.N. Effect of PGPR on growth promotion of rice (Oryza sativa L.) under salt stress. Asian J. Plant Sci. Res. 2014, 4, 62–67. [Google Scholar]
- Stegelmeier, A.A.; Rose, D.M.; Joris, B.R.; Glick, B.R. The Use of PGPB to Promote Plant Hydroponic Growth. Plants 2022, 11, 2783. [Google Scholar] [CrossRef] [PubMed]
- Barker, K.R.; Koenning, S.R. Developing sustainable systems for nematode management. Annu. Rev. Phytopathol. 1998, 36, 165–205. [Google Scholar]
- Zapata-Vahos, I.C.; Rojas-Rodas, F.; David, D.; Gutierrez-Monsalve, J.A.; Castro-Restrepo, D. Comparison of antioxidant contents of green and red leaf lettuce cultivated in hydroponic systems in greenhouses and conventional soil cultivation. Rev. Fac. Nac. Agron. Medellín 2020, 73, 9077–9088. [Google Scholar] [CrossRef]
- Nair, A.G.; Chacko, A.; Mohan, G.; Francis, T.K. Smart vertical farming using hydroponics. J. Electr. Electron. Eng. 2015, 1, 14–17. [Google Scholar]
- Panno, S.; Davino, S.; Caruso, A.G.; Bertacca, S.; Crnogorac, A.; Mandi´c, A.; Noris, E.; Mati´c, S. A review of the most common and economically important diseases that undermine the cultivation of tomato crop in the mediterranean basin. Agronomy 2021, 11, 2188. [Google Scholar] [CrossRef]
- O’Callaghan, M. Microbial Inoculation of Seed for Improved Crop Performance: Issues and Opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef]
- John, C.J.; Kumar, S.; Ge, M. Probiotic Prospects of PGPR for Green and Sustainable Agriculture. Arch. Phytopathol. Plant Prot. 2020, 53, 899–914. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M. Co-Application of ACC-Deaminase Producing PGPR and Timber-Waste Biochar 1034 Improves Pigments Formation, Growth and Yield of Wheat under Drought Stress. Sci. Rep. 2019, 9, 1–13. [Google Scholar]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C. Co-Inoculation Effect of Rhizobia and Plant Growth Promoting Rhizobacteria on Common Bean Growth in a Low Phosphorus Soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Arshad, M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar]
- Htwe, A.Z.; Moh, S.M.; Soe, K.M.; Moe, K.; Yamakawa, T. Effects of biofertilizer produced from Bradyrhizobium and Streptomyces griseoflavus on plant growth, nodulation, nitrogen fixation, nutrient uptake, and seed yield of mung bean, cowpea, and soybean. Agronomy 2019, 9, 77. [Google Scholar] [CrossRef]
- Wiggins, Z.; Akaeze, O.; Nandwani, D.; Witcher, A. Substrate properties and fertilizer rates on yield responses of lettuce in a vertical growth system. Sustainability 2020, 12, 6465. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of salt stress by plant growth-promoting bacteria in hydroponic leaf lettuce. Agronomy 2020, 10, 1523. [Google Scholar]
- Settanni, L.; Miceli, A.; Francesca, N.; Cruciata, M.; Moschetti, G. Microbiological Investigation of Raphanus sativus L. Grown Hydroponically in Nutrient Solutions Contaminated with Spoilage and Pathogenic Bacteria. Int. J. Food Microbiol. 2013, 160, 344–352. [Google Scholar]
- Stouvenakers, G.; Dapprich, P.; Massart, S.; Jijakli, M.H. Plant Pathogens and Control Strategies in Aquaponics. In Aquaponics Food Production Systems; Springer: Cham, Switzerland, 2019; p. 353. [Google Scholar]
- Bjarnsholt, T.; Van Gennip, M.; Jakobsen, T.H.; Christensen, L.D.; Jensen, P.Ø.; Givskov, M. In Vitro Screens for Quorum Sensing Inhibitors and in vivo Confirmation of Their Effect. Nat. Protoc. 2010, 5, 282–293. [Google Scholar] [CrossRef]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhou, T.; Li, Q.; Xu, X.; Fan, X.; Zhang, L.; Chen, S. Cupriavidus sp. HN-2, a Novel Quorum Quenching Bacterial Isolate, Is a Potent Biocontrol Agent against Xanthomonas campestris pv. campestris. Microorganisms 2020, 8, 45. [Google Scholar]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S. Hydroponic Solutions for Soilless Production Systems: Issues and Opportunities in a Smart Agriculture Perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar]
- Gravel, V.; Martinez, C.; Antoun, H.; Tweddell, R.J. Control of greenhouse tomato root rot [Pythium ultimum] in hydroponic systems, using plant-growth-promoting microorganisms. Can. J. Plant Pathol. 2006, 28, 475–483. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; De Micco, V.; Giordano, M.; Aronne, G.; De Pascale, S. Changes in leaf anatomical traits enhanced photosynthetic activity of soybean grown in hydroponics with plant growth-promoting microorganisms. Front. Plant Sci. 2017, 8, 674. [Google Scholar] [PubMed] [Green Version]
- Varia, J.; Kamaleson, C.; Lerer, L. Biostimulation with phycocyanin-rich Spirulina extract in hydroponic vertical farming. Sci. Hortic. 2022, 299, 111042. [Google Scholar]


{kind=link}
| Plant | Mode of PGPM Application | Type of PGPM | Influence | References |
|---|---|---|---|---|
| Banana Berangan’ (Musa spp. dessert type) | Seedling inoculation with microbial suspension | Bacillus sphaericus and azospirillum | Increase in root formation, leaf area, chlorophyll content, and consequently, total biomass | [28] |
| Strawberries | Siderophores added to hydroponic medium | Gluconacetobacter diazotrophicus and azospirillum brasilense | Increased the nutrition of iron | [30] |
| Triticum aestivum | Seedling inoculation with microbial suspension | Calothrix sp., anabaena cylindrica, chryseobacterium balustinum, pseudomonas simiae, and pseudomonas fluorescen | Increased the growth, plant height, dry shoot mass, total nutrients, and the ability to produce indole acetic acid | [31] |
| Lettuce | Biostimulant extract added to hydroponic medium | Bacillus spp. | Minimized salt stress | [96] |
| Tomato | Seedling inoculation with microbial suspension | Penicillium brevicompactum, penicillium solitum strain 1, pseudomonas fluorescens subgroup g strain 2, pseudomonas marginalis, pseudomonas putida subgroup b strain 1, pseudomonas syringae strain 1, and trichoderma atroviride | Plant growth and development in the absence of pathogens (antagonistic activity against Pythium ultimum) | [103] |
| Glycine max (L.) Merr. | Seedling inoculation with microbial suspension | Bacteria, yeasts, mycorrhiza, and Trichoderma | Higher density of smaller stomata, thicker palisade parenchyma, larger intercellular spaces in the mesophyll; increased photosynthetic traits, growth and seed production | [104] |
| Lettuce (Salanova® Lactuca sativa and Salanova® Red Crisp). | Bio-stimulant extract added to hydroponic medium | Phycocyanin-rich spirulina extract | Reduced time from seed to harvest by 6 days, increased yield by 12.5%, and improved antioxidant flavonoid levels | [105] |
| Hydroponic Types | Definition | Suggested Mode of PGPM Application |
|---|---|---|
| Wicking System | Small-scale production plants (small plants, e.g., herbs and leafy greens) | Pre-treatment seedling inoculation with microbial suspension |
| Ebb and Flow (Flood and Drain) | Seedling cultivation in commercial settings | Pre-treatment seedling inoculation with microbial suspension |
| Drip Irrigation | Commercial production for larger fruiting crops such as tomatoes, cucumbers, peppers, and strawberries | Synchronize treatment: Bio-stimulant extract added to hydroponic medium |
| Nutrient Film Technique | In commercial use for smaller leafy crops such as lettuce | Pre-treatment seedling inoculation with microbial suspension |
| Deep Water Culture | In commercial settings deep water culture systems are used for small leafy plants such as lettuce or herbs | Pre-treatment seedling inoculation with microbial suspension |
| Aeroponics | Commercial growers to produce small leafy plants and potato mini-tubers | Pre-treatment seedling inoculation with microbial suspension |
| Aquaponics | Growing fish and plants in the same system | Pre-treatment seedling inoculation with microbial suspension |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhawi, F. The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites 2023, 13, 247. https://doi.org/10.3390/metabo13020247
Dhawi F. The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites. 2023; 13(2):247. https://doi.org/10.3390/metabo13020247
Chicago/Turabian StyleDhawi, Faten. 2023. "The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming" Metabolites 13, no. 2: 247. https://doi.org/10.3390/metabo13020247
APA StyleDhawi, F. (2023). The Role of Plant Growth-Promoting Microorganisms (PGPMs) and Their Feasibility in Hydroponics and Vertical Farming. Metabolites, 13(2), 247. https://doi.org/10.3390/metabo13020247