
Lipidomic Analysis of Hand Skin Surface Lipids Reveals Smoking-Related Skin Changes




Abstract
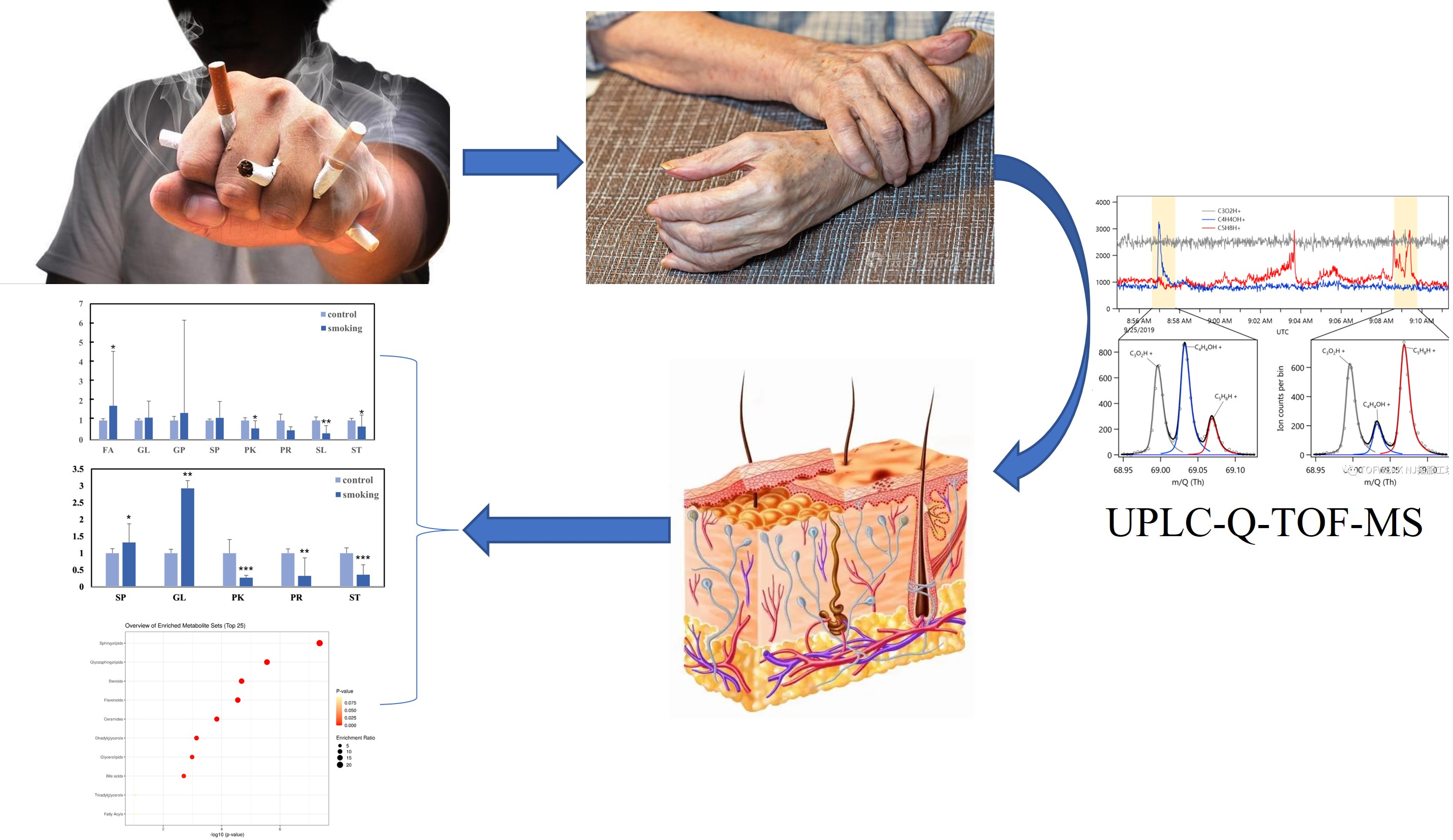
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Study Population
2.3. Sample Collection and Storage
2.4. Sample Treatment
2.5. Analysis Conditions
2.5.1. Liquid Chromatography Conditions
2.5.2. Mass Spectrometry Conditions
2.6. Multivariate Data Analysis and Statistical Analysis
3. Results
3.1. Variations in the Main Class of Lipids
3.2. OPLS-DA Analysis of Hand Lipids from the Smoking Group and the Control Group
3.3. Identification of Important Individual Lipids in the Smoking and Control Groups
3.4. Enrichment Analysis of Skin Lipid Metabolism between Smoking and Control Groups
4. Discussion
4.1. Analysis of Differences in Sphingolipid
4.2. Analysis of Differences in Glycerolipids
4.3. Analysis of Differences in Sterol Lipids and Polyketides
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gravitz, L. Skin. Nature 2018, 563, S83. [Google Scholar] [CrossRef]
- Knox, S.; O’Boyle, N.M. Skin lipids in health and disease: A review. Chem. Phys. Lipids 2021, 236, 105055. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Honeywell-Nguyen, P.L. Skin structure and mode of action of vesicles. Adv. Drug Deliv. Rev. 2002, 54 (Suppl. S1), S41–S55. [Google Scholar] [CrossRef]
- Kruse, V.; Neess, D.; Færgeman, N.J. The Significance of Epidermal Lipid Metabolism in Whole-Body Physiology. Trends Endocrinol. Metab. TEM 2017, 28, 669–683. [Google Scholar] [CrossRef]
- Jia, Y.; Gan, Y.; He, C.; Chen, Z.; Zhou, C. The mechanism of skin lipids influencing skin status. J. Dermatol. Sci. 2018, 89, 112–119. [Google Scholar] [CrossRef]
- Cui, L.; Jia, Y.; Cheng, Z.-W.; Gao, Y.; Zhang, G.-L.; Li, J.-Y.; He, C.-F. Advancements in the maintenance of skin barrier/skin lipid composition and the involvement of metabolic enzymes. J. Cosmet. Dermatol. 2016, 15, 549–558. [Google Scholar] [CrossRef]
- Coderch, L.; López, O.; de la Maza, A.; Parra, J.L. Ceramides and skin function. Am. J. Clin. Dermatol. 2003, 4, 107–129. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.P.; Zhao, M.T.; Jia, Z.X.; Zhang, J.L.; Jiao, L.; Ma, L. Skin Ceramide Profile in Children with Atopic Dermatitis. Dermat. Contact Atopic Occup. Drug 2018, 29, 219–222. [Google Scholar] [CrossRef]
- Meckfessel, M.H.; Brandt, S. The structure, function, and importance of ceramides in skin and their use as therapeutic agents in skin-care products. J. Am. Acad. Dermatol. 2014, 71, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Clayton, R.W.; Göbel, K.; Niessen, C.M.; Paus, R.; van Steensel, M.A.M.; Lim, X. Homeostasis of the sebaceous gland and mechanisms of acne pathogenesis. Br. J. Dermatol. 2019, 181, 677–690. [Google Scholar] [CrossRef]
- Crivellari, I.; Sticozzi, C.; Belmonte, G.; Muresan, X.M.; Cervellati, F.; Pecorelli, A.; Cavicchio, C.; Maioli, E.; Zouboulis, C.C.; Benedusi, M.; et al. SRB1 as a new redox target of cigarette smoke in human sebocytes. Free Radic. Biol. Med. 2017, 102, 47–56. [Google Scholar] [CrossRef]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef]
- Zhou, M.; Yang, M.; Zheng, Y.; Dong, K.; Song, L.; He, C.; Liu, W.; Wang, Y.; Jia, Y. Skin surface lipidomics revealed the correlation between lipidomic profile and grade in adolescent acne. J. Cosmet. Dermatol. 2020, 19, 3349–3356. [Google Scholar] [CrossRef] [PubMed]
- Bocheńska, K.; Gabig-Cimińska, M. Unbalanced Sphingolipid Metabolism and Its Implications for the Pathogenesis of Psoriasis. Molecules 2020, 25, 1130. [Google Scholar] [CrossRef]
- Nicolaou, A.; Pilkington, S.M.; Rhodes, L.E. Ultraviolet-radiation induced skin inflammation: Dissecting the role of bioactive lipids. Chem. Phys. Lipids 2011, 164, 535–543. [Google Scholar] [CrossRef]
- De Luca, C.; Valacchi, G. Surface lipids as multifunctional mediators of skin responses to environmental stimuli. Mediat. Inflamm. 2010, 2010, 321494. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhou, M.; Li, Y.; Huang, H.; Jia, Y. Lipidomic analysis of facial skin surface lipid reveals the causes of pregnancy-related skin barrier weakness. Sci. Rep. 2021, 11, 3229. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, J.W.; Blaha, M.J.; DeFilippis, A.P.; Lima, J.A.; Bluemke, D.A.; Hundley, W.G.; Min, J.K.; Shaw, L.J.; Lloyd-Jones, D.M.; Barr, R.G.; et al. Cigarette smoking and cardiovascular events: Role of inflammation and subclinical atherosclerosis from the MultiEthnic Study of Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 700–709. [Google Scholar] [CrossRef]
- Wheaton, A.G.; Liu, Y.; Croft, J.B.; VanFrank, B.; Croxton, T.L.; Punturieri, A.; Postow, L.; Greenlund, K.J. Chronic Obstructive Pulmonary Disease Smoking Status—United States 2017. Morb. Mortal. Wkly. Rep. 2019, 68, 533–538. [Google Scholar] [CrossRef]
- Li, L.; Yang, D.C.; Chen, C.H. Metabolic reprogramming: A driver of cigarette smoke-induced inflammatory lung diseases. Free Radic. Biol. Med. 2021, 163, 392–401. [Google Scholar] [CrossRef]
- Ortiz, A.; Grando, S.A. Smoking and the skin. Int. J. Dermatol. 2012, 51, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Langton, A.K.; Tsoureli-Nikita, E.; Merrick, H.; Zhao, X.; Antoniou, C.; Stratigos, A.; Akhtar, R.; Derby, B.; Sherratt, M.J.; Watson, R.E.; et al. The systemic influence of chronic smoking on skin structure and mechanical function. J. Pathol. 2020, 251, 420–428. [Google Scholar] [CrossRef]
- Kovitwanichkanont, T.; Chong, A.H.; Foley, P. Beyond skin deep: Addressing comorbidities in psoriasis. Med. J. Aust. 2020, 212, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Gyger, G.; Hudson, M.; Lo, E.; Steele, R.; Baron, M. Does cigarette smoking mitigate the severity of skin disease in systemic sclerosis? Rheumatol. Int. 2013, 33, 943–948. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Armstrong, E.J.; Fuller, E.N.; Sockolov, M.E.; Voyles, S.V. Smoking and pathogenesis of psoriasis: A review of oxidative, inflammatory and genetic mechanisms. Br. J. Dermatol. 2011, 165, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, T.; Klose, C.; Gerl, M.J.; Wójcik-Maciejewicz, A.; Herzog, R.; Simons, K.; Reich, A.; Surma, M.A. Large-scale human skin lipidomics by quantitative, high-throughput shotgun mass spectrometry. Sci. Rep. 2017, 7, 43761. [Google Scholar] [CrossRef]
- Puri, P.; Nandar, S.K.; Kathuria, S.; Ramesh, V. Effects of air pollution on the skin: A review. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 415–423. [Google Scholar] [CrossRef]
- Breslow, D.K.; Weissman, J.S. Membranes in balance: Mechanisms of sphingolipid homeostasis. Mol. Cell 2010, 40, 267–279. [Google Scholar] [CrossRef]
- Li, Q.; Fang, H.; Dang, E.; Wang, G. The role of ceramides in skin homeostasis and inflammatory skin diseases. J. Dermatol. Sci. 2020, 97, 2–8. [Google Scholar] [CrossRef]
- Bodas, M.; Pehote, G.; Silverberg, D.; Gulbins, E.; Vij, N. Autophagy augmentation alleviates cigarette smoke-induced CFTR-dysfunction, ceramide-accumulation and COPD-emphysema pathogenesis. Free Radic. Biol. Med. 2019, 131, 81–97. [Google Scholar] [CrossRef]
- Kornhuber, J.; Hoertel, N.; Gulbins, E. The acid sphingomyelinase/ceramide system in COVID-19. Mol. Psychiatry 2022, 27, 307–314. [Google Scholar] [CrossRef]
- Zulueta, A.; Caretti, A.; Campisi, G.M.; Brizzolari, A.; Abad, J.L.; Paroni, R.; Signorelli, P.; Ghidoni, R. Inhibitors of ceramide de novo biosynthesis rescue damages induced by cigarette smoke in airways epithelia. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2017, 390, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Xin, S.; Ye, L.; Man, G.; Lv, C.; Elias, P.M.; Man, M.Q. Heavy Cigarette Smokers in a Chinese Population Display a Compromised Permeability Barrier. BioMed Res. Int. 2016, 2016, 9704598. [Google Scholar] [CrossRef] [PubMed]
- Cumin, C.; Huang, Y.L.; Everest-Dass, A.; Jacob, F. Deciphering the Importance of Glycosphingolipids on Cellular and Molecular Mechanisms Associated with Epithelial-to-Mesenchymal Transition in Cancer. Biomolecules 2021, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Kondo, Y.; Ohkawa, Y.; Tajima, O.; Furukawa, K. Regulatory function of glycosphingolipids in the inflammation and degeneration. Arch. Biochem. Biophys. 2015, 571, 58–65. [Google Scholar] [CrossRef]
- Zhang, T.; de Waard, A.A.; Wuhrer, M.; Spaapen, R.M. The Role of Glycosphingolipids in Immune Cell Functions. Front. Immunol. 2019, 10, 90. [Google Scholar] [CrossRef]
- Dany, M.; Elston, D. Gene expression of sphingolipid metabolism pathways is altered in hidradenitis suppurativa. J. Am. Acad. Dermatol. 2017, 77, 268–273. [Google Scholar] [CrossRef]
- Sochorová, M.; Staňková, K.; Pullmannová, P.; Kováčik, A.; Zbytovská, J.; Vávrová, K. Permeability Barrier and Microstructure of Skin Lipid Membrane Models of Impaired Glucosylceramide Processing. Sci. Rep. 2017, 7, 6470. [Google Scholar] [CrossRef]
- Holleran, W.M.; Ginns, E.I.; Menon, G.K.; Grundmann, J.U.; Fartasch, M.; McKinney, C.E.; Elias, P.M.; Sidransky, E. Consequences of beta-glucocerebrosidase deficiency in epidermis. Ultrastructure and permeability barrier alterations in Gaucher disease. J. Clin. Investig. 1994, 93, 1756–1764. [Google Scholar] [CrossRef]
- Tascini, A.S.; Noro, M.G.; Chen, R.; Seddon, J.M.; Bresme, F. Understanding the interactions between sebum triglycerides and water: A molecular dynamics simulation study. Phys. Chem. Chem. Phys. PCCP 2018, 20, 1848–1860. [Google Scholar] [CrossRef]
- Raghow, R.; Yellaturu, C.; Deng, X.; Park, E.A.; Elam, M.B. SREBPs: The crossroads of physiological and pathological lipid homeostasis. Trends Endocrinol. Metab. TEM 2008, 19, 65–73. [Google Scholar] [CrossRef]
- Yuan, H.; Shyy, J.Y.; Martins-Green, M. Second-hand smoke stimulates lipid accumulation in the liver by modulating AMPK and SREBP-1. J. Hepatol. 2009, 51, 535–547. [Google Scholar] [CrossRef]
- Bizoń, A.; Milnerowicz, H. The effect of passive and active exposure to tobacco smoke on lipid profile parameters and the activity of certain membrane enzymes in the blood of women in the first trimester of pregnancy. Environ. Toxicol. Pharmacol. 2017, 53, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Elias, P.M.; Williams, M.L.; Holleran, W.M.; Jiang, Y.J.; Schmuth, M. Pathogenesis of permeability barrier abnormalities in the ichthyoses: Inherited disorders of lipid metabolism. J. Lipid Res. 2008, 49, 697–714. [Google Scholar] [CrossRef]
- Amin, R.; Lechner, A.; Vogt, A.; Blume-Peytavi, U.; Kottner, J. Molecular characterization of xerosis cutis: A systematic review. PLoS ONE 2021, 16, e0261253. [Google Scholar] [CrossRef]
- Bhattacharya, N.; Sato, W.J.; Kelly, A.; Ganguli-Indra, G.; Indra, A.K. Epidermal Lipids: Key Mediators of Atopic Dermatitis Pathogenesis. Trends Mol. Med. 2019, 25, 551–562. [Google Scholar] [CrossRef]
- Li, W.H.; Zhang, Q.; Flach, C.R.; Mendelsohn, R.; Southall, M.D.; Parsa, R. In vitro modeling of unsaturated free fatty acid-mediated tissue impairments seen in acne lesions. Arch. Dermatol. Res. 2017, 309, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Eichmann, T.O.; Lass, A. DAG tales: The multiple faces of diacylglycerol--stereochemistry, metabolism, and signaling. Cell. Mol. Life Sci. CMLS 2015, 72, 3931–3952. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Long, Y.; Xu, D.M.; Zhu, B.L.; Deng, X.J.; Yan, Z.; Sun, F.; Chen, G.J. Age- and Nicotine-Associated Gene Expression Changes in the Hippocampus of APP/PS1 Mice. J. Mol. Neurosci. MN 2019, 69, 608–622. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Valencia, J.C.; Namiki, T.; Suzuki, T.; Hearing, V.J. Diacylglycerol kinase regulates tyrosinase expression and function in human melanocytes. J. Investig. Dermatol. 2012, 132, 2791–2799. [Google Scholar] [CrossRef] [Green Version]
- Chibalin, A.V.; Leng, Y.; Vieira, E.; Krook, A.; Björnholm, M.; Long, Y.C.; Kotova, O.; Zhong, Z.; Sakane, F.; Steiler, T. Downregulation of diacylglycerol kinase delta contributes to hyperglycemia-induced insulin resistance. Cell 2008, 132, 375–386. [Google Scholar] [CrossRef]
- Oka, M.; Kageshita, T.; Ono, T.; Goto, A.; Kuroki, T.; Ichihashi, M. Protein kinase C alpha associates with phospholipase D1 and enhances basal phospholipase D activity in a protein phosphorylation-independent manner in human melanoma cells. J. Investig. Dermatol. 2003, 121, 69–76. [Google Scholar] [CrossRef]
- Brown, D.A. Skin pigmentation enhancers. J. Photochem. Photobiol. B Biol. 2001, 63, 148–161. [Google Scholar] [CrossRef] [PubMed]
- West, H.; Reid, G.E. Hybrid 213 nm photodissociation of cationized Sterol lipid ions yield [M](+.) Radical products for improved structural characterization using multistage tandem mass spectrometry. Anal. Chim. Acta 2021, 1141, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Kloudova-Spalenkova, A.; Holy, P.; Soucek, P. Oxysterols in cancer management: From therapy to biomarkers. Br. J. Pharmacol. 2021, 178, 3235–3247. [Google Scholar] [CrossRef] [PubMed]
- Caliri, A.W.; Tommasi, S.; Besaratinia, A. Relationships among smoking, oxidative stress, inflammation, macromolecular damage, and cancer. Mutation research. Rev. Mutat. Res. 2021, 787, 108365. [Google Scholar] [CrossRef]
- de Medina, P.; Silvente-Poirot, S.; Poirot, M. Oxysterols are potential physiological regulators of ageing. Ageing Res. Rev. 2022, 77, 101615. [Google Scholar] [CrossRef]
- Zarrouk, A.; Vejux, A.; Mackrill, J.; O’Callaghan, Y.; Hammami, M.; O’Brien, N.; Lizard, G. Involvement of oxysterols in age-related diseases and ageing processes. Ageing Res. Rev. 2014, 18, 148–162. [Google Scholar] [CrossRef]
- Xu, X.; Qu, R.; Wu, W.; Jiang, C.; Shao, D.; Shi, J. Applications of microbial co-cultures in polyketides production. J. Appl. Microbiol. 2021, 130, 1023–1034. [Google Scholar] [CrossRef]
- Yang, M.; Gao, Y.; Zhou, M.; Wang, H.; Jia, Y.; He, C.; Gu, F.; Liu, X. Lipidomics analysis of facial skin surface lipids in neonates with acne. Chin. J. Dermatol. 2020, 53, 914–916. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Accepted Description | m/z | Anova (p) | q Value | Max Fold Change | Highest Mean |
|---|---|---|---|---|---|---|
| Sphingolipids | Cer(d18:0/28:0(28OH)) | 724.7194 | 0.001333 | 0.006793 | 2.530351 | smoking |
| Sphingolipids | Cer(d18:2/30:0) | 732.725 | 0.003913 | 0.017549 | 2.980895 | smoking |
| Sphingolipids | Cer(d18:2/28:0(2OH)) | 720.6875 | 0.016101 | 0.052893 | 2.397683 | smoking |
| Sphingolipids | Cer(d18:0/30:0(30OH)) | 752.751 | 5.17 × 10−05 | 0.000421 | 3.519314 | smoking |
| Sphingolipids | GlcCer(d15:2(4E,6E)/18:0) | 684.5442 | 1.18 × 10−14 | 7.67 × 10−13 | Infinity | control |
| Sphingolipids | GlcCer(d14:2(4E,6E)/16:0) | 642.4955 | 2.74 × 10−07 | 3.94 × 10−06 | 2.452004 | control |
| Sphingolipids | GlcCer(d15:2(4E,6E)/22:0) | 740.6035 | 9.04 × 10−10 | 2.25 × 10−08 | 3.328931 | smoking |
| Sphingolipids | GlcCer(d18:2/22:0) | 782.6508 | 6.06 × 10−06 | 6.58 × 10−05 | 3.283642 | smoking |
| Sphingolipids | Galactosylceramide (d18:1/18:1(9Z)) | 726.5911 | 0 | 0 | Infinity | control |
| Sphingolipids | Galactosylceramide (d18:1/26:1(17Z)) | 838.7142 | 8.22 × 10−08 | 1.37 × 10−06 | 3.35593 | smoking |
| Sphingolipids | Sphing-6E-enine 4R-sulfate | 413.2672 | 5.89 × 10−05 | 0.000475 | 2.987902 | control |
| Glycerolipids | TGs(50:0) | 852.8037 | 6.10 × 10−05 | 0.000487 | 3.870762 | smoking |
| Glycerolipids | TGs(52:0) | 880.8356 | 0.000234 | 0.001585 | 6.50787 | smoking |
| Glycerolipids | TGs(58:5) | 954.8499 | 0.043424 | 0.11684 | 5.250341 | control |
| Glycerolipids | TGs(58:2) | 960.8959 | 0.04401 | 0.118043 | 2.951669 | control |
| Glycerolipids | DG(38:3) | 647.5596 | 1.40 × 10−13 | 7.44 × 10−12 | 437.4144 | smoking |
| Glycerolipids | DG(36:3) | 619.5285 | 7.41 × 10−12 | 2.99 × 10−10 | 72.23256 | smoking |
| Glycerolipids | DG(34:3) | 591.4976 | 0.001338 | 0.006793 | 13.26929 | smoking |
| Sterol Lipids | Cyclopassifloic acid E | 570.4028 | 8.61E-09 | 1.74 × 10−07 | 4.337149 | control |
| Sterol Lipids | Acetylpinnasterol | 489.3218 | 1.49 × 10−05 | 0.000146 | 3.667711 | control |
| Sterol Lipids | Echinasteroside C | 630.4237 | 2.36 × 10−05 | 0.000219 | 3.118912 | control |
| Sterol Lipids | 12alpha-Hydroxy-3-oxo-5beta-cholan-24-oic Acid | 391.2853 | 3.31 × 10−05 | 0.000296 | 4.530213 | control |
| Polyketides | 4′-Hydroxy-5,7-dimethoxy-8-methylflavan | 301.1435 | 0 | 0 | Infinity | control |
| Polyketides | Squamocin-V | 640.5516 | 3.46 × 10−06 | 4.08 × 10−05 | 2.343713 | control |
| Polyketides | Formononetin 7-O-(6″-acetylglcoside) | 473.1465 | 9.89 × 10−06 | 0.000102 | 2.937326 | control |
| Prenol Lipids | bacteriohopane-32,33,34-triol-35-cyclitolguanine | 750.5594 | 0.006071 | 0.024858 | 3.100748 | control |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Zhao, M.; Mao, Z. Lipidomic Analysis of Hand Skin Surface Lipids Reveals Smoking-Related Skin Changes. Metabolites 2023, 13, 254. https://doi.org/10.3390/metabo13020254
Chen T, Zhao M, Mao Z. Lipidomic Analysis of Hand Skin Surface Lipids Reveals Smoking-Related Skin Changes. Metabolites. 2023; 13(2):254. https://doi.org/10.3390/metabo13020254
Chicago/Turabian StyleChen, Tian, Mengzhen Zhao, and Zhenxing Mao. 2023. "Lipidomic Analysis of Hand Skin Surface Lipids Reveals Smoking-Related Skin Changes" Metabolites 13, no. 2: 254. https://doi.org/10.3390/metabo13020254
APA StyleChen, T., Zhao, M., & Mao, Z. (2023). Lipidomic Analysis of Hand Skin Surface Lipids Reveals Smoking-Related Skin Changes. Metabolites, 13(2), 254. https://doi.org/10.3390/metabo13020254