

Oxygen Uptake Kinetics and Time Limit at Maximal Aerobic Workload in Tethered Swimming

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Maximal Force Testing in Tethered Swimming
2.4. Incremental Tethered Test (ITT)
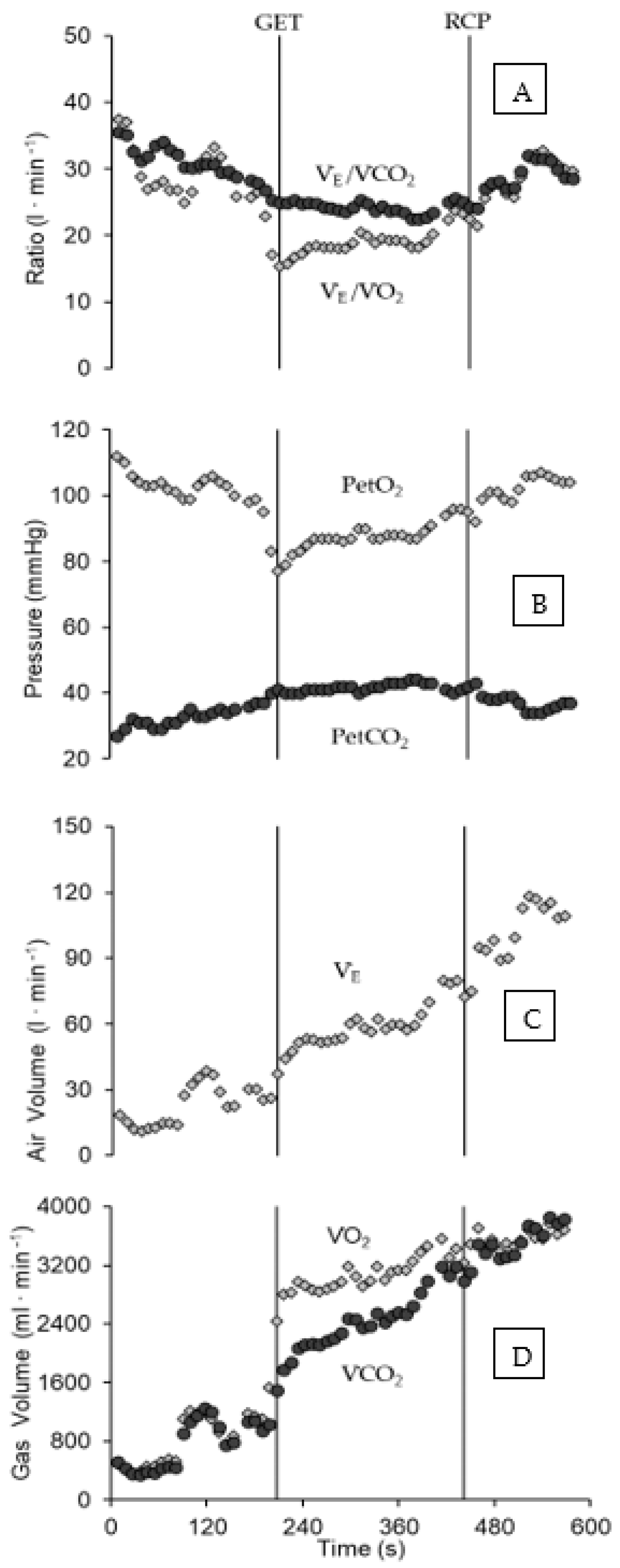
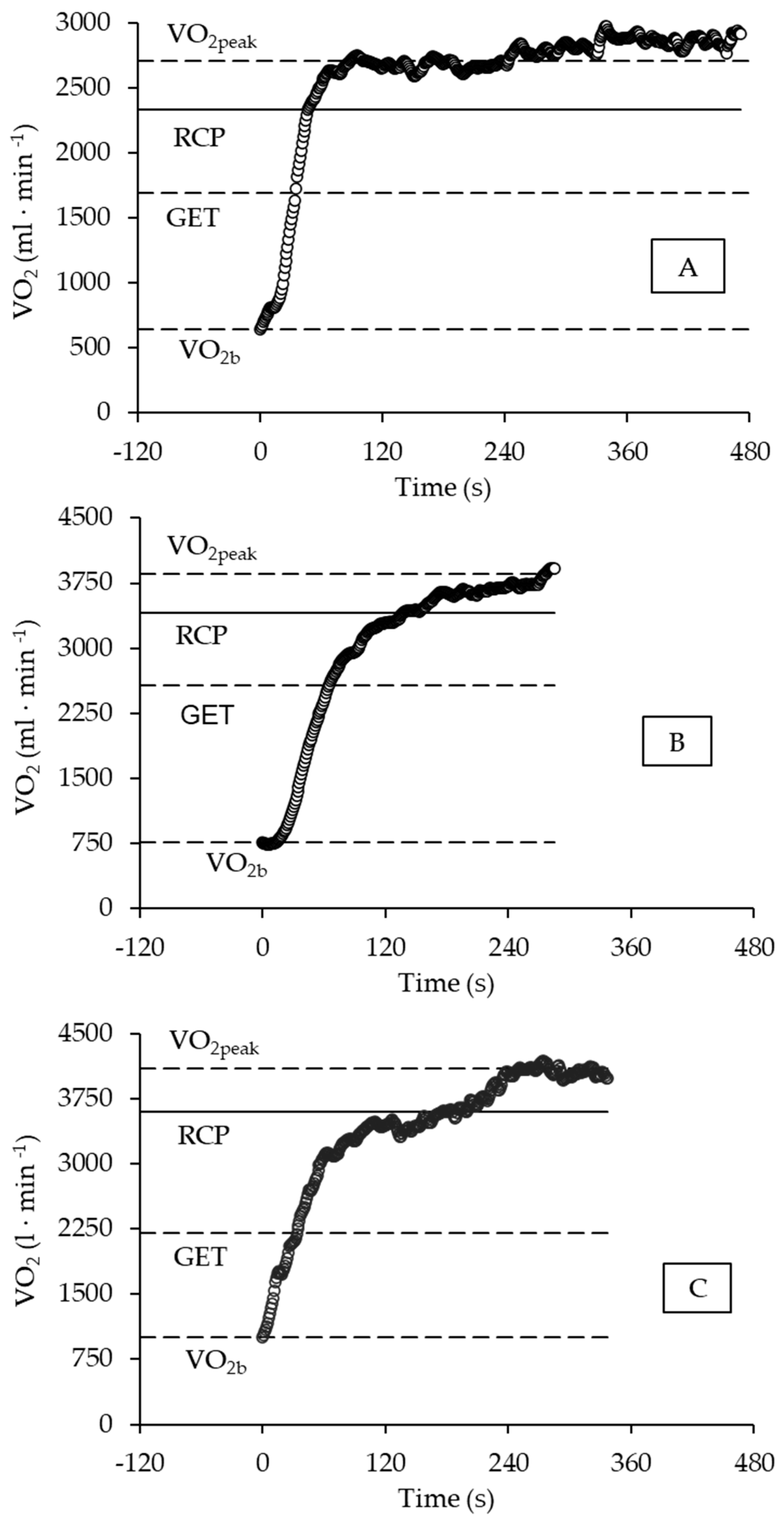
2.5. Analysis of VO2 Kinetics during Exercise
2.6. Statistical Analysis
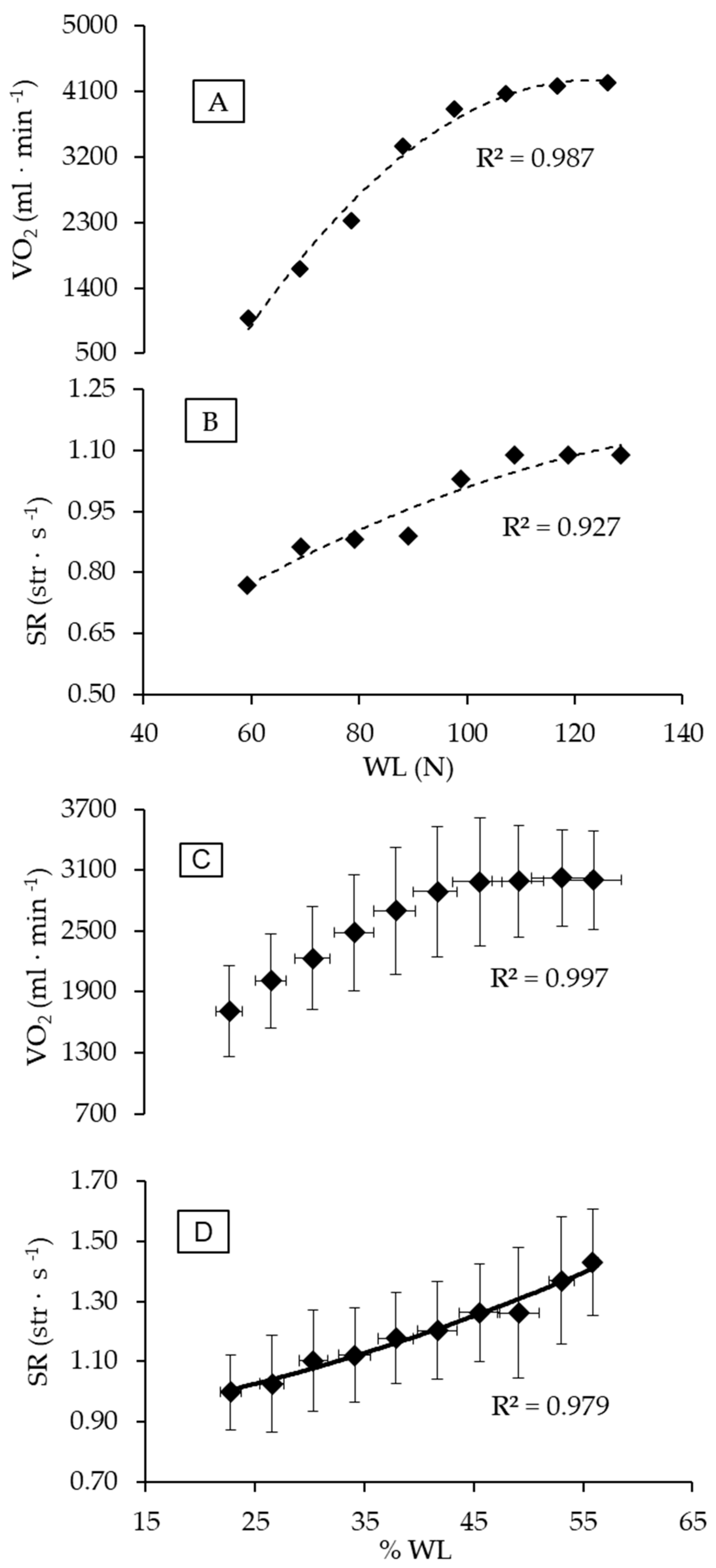
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samson, M.; Monnet, T.; Bernard, A.; Lacouture, P.; David, L. Comparative study between fully tethered and free swimming at different paces of swimming in front crawl. Sport. Biomech. 2019, 18, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Morouço, P.; Keskinen, K.L.; Vilas-Boas, J.P.; Fernandes, R.J. Relationship between tethered forces and the four swimming techniques performance. J. Appl. Biomech. 2011, 27, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro, N.; Marinho, D.A.; Batalha, N.; Marques, M.C.; Morouço, P. Reliability of tethered swimming evaluation in age group swimmers. J. Hum. Kinet. 2014, 41, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeater, R.A.; Martin, R.B.; White, M.K.; Gilson, K.H. Tethered swimming forces in the crawl, breast and back strokes and their relationship to competitive performance. J. Biomech. 1981, 14, 527–537. [Google Scholar] [CrossRef]
- Dopsaj, M.; Matkovic, I.; Zdravkovic, I. The relationship between 50 m—Freestyle results and characteristics of tethered forces in male sprint swimmers: A new approach to tethered swimming test. Facta Univ. Phys. Edu. Sport. 2000, 1, 15–22. [Google Scholar]
- Pinna, M.; Milia, R.; Roberto, S.; Marongiu, E.; Olla, S.; Loi, A.; Ortu, M.; Migliaccio, G.M.; Tocco, F.; Concu, A.; et al. Assessment of the specificity of cardiopulmonary response during tethered swimming using a new snorkel device. J. Physiol. Sci. 2013, 63, 7–16. [Google Scholar] [CrossRef]
- Pessôa Filho, D.M.; Greco, C.C.; Denadai, B.S. Tether-power at maximal lactate steady-state and endurance indexes of swimming performance. Rev. Bras. Med. Esporte 2014, 20, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Papoti, M.; Da Silva, A.S.R.; Araújo, G.G.; Santiago, V.; Martins, L.E.B.; Cunha, S.A.; Gobatto, C.A. Aerobic and anaerobic performances in tethered swimming. Int. J. Sport. Med. 2013, 34, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Pessôa Filho, D.M.; SiqueiraI, L.O.; Simionato, A.R.; Espada, M.A.; Pestana, D.S.; DiMenna, F.J. A rapidly-incremented tethered-swimming test for defining domain-specific training zones. J. Hum. Kinet. 2017, 57, 117–128. [Google Scholar] [CrossRef]
- Holmér, I. Oxygen uptake during swimming in man. J. Appl. Physiol. 1972, 33, 502–509. [Google Scholar] [CrossRef]
- Bonen, A.; Wilson, B.; Yarkony, M.; Belcastro, A.N. Maximal oxygen uptake during free, tethered, and flume swimming. J. Appl. Physiol. 1980, 48, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Rinehardt, K.F.; Kraemer, R.R.; Gormely, S.; Colan, S. Comparison of maximal oxygen uptakes from the tethered, the 183- and 457-meter unimpeded supramaximal freestyle swims. Int. J. Sport. Med. 1991, 12, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Pessôa Filho, D.M.; Massini, D.A.; SiqueiraI, L.O.; Santos, L.G.A.; Vasconcelos, C.M.T.; Almeida, T.A.F.; Espada, M.A.; Reis, J.F.; Alves, F.B.; DiMenna, F.J. A rapidly incremented tethered-swimming maximal protocol for cardiorespiratory assessment of swimmers. J. Vis. Exp. 2020, 155, e60630. [Google Scholar] [CrossRef]
- Sousa, A.; Vilas-Boas, J.P.; Fernandes, R. VO2 kinetics and metabolic contributions whilst swimming at 95, 100, and 105% of the velocity at VO2max. Biomed Res. Int. 2014, 2014, 675363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessôa Filho, D.; Alves, F.; Reis, J.; Greco, C.; Denadai, B. VO2 kinetics during heavy and severe exercise in swimming. Int. J. Sport. Med. 2012, 33, 744–748. [Google Scholar] [CrossRef]
- Almeida, T.A.F.; Pessôa Filho, D.M.; Espada, M.A.C.; Reis, J.F.; Simionato, A.R.; Siqueira, L.O.C.; Alves, F.B. VO2 kinetics kinetics and energy contribution in simulated maximal performance during short and middle distance-trials in swimming. Eur. J. Appl. Physiol. 2020, 120, 1097–1109. [Google Scholar] [CrossRef]
- Rodríguez, F.; Lätt, E.; Jürimäe, J.; Maestu, J.; Purge, P.; Rämson, R.; Haljaste, K.; Keskinen, K.; Jürimäe, T. VO2 Kinetics in all-out arm stroke, leg kick and whole stroke front crawl 100-m swimming. Int J Sport. Med. 2015, 37, 191–196. [Google Scholar] [CrossRef]
- Reis, J.; Alves, F.; Bruno, P.; Vleck, V.; Millet, G. Oxygen uptake kinetics and middle-distance swimming performance. J. Sci. Med. Sport. 2012, 15, 58–63. [Google Scholar] [CrossRef]
- Espada, M.C.; Reis, J.F.; Almeida, T.F.; Bruno, P.M.; Vleck, V.E.; Alves, F.B. Ventilatory and physiological responses in swimmers below and above their maximal lactate steady state. J. Strength Cond. Res. 2015, 29, 2836–2843. [Google Scholar] [CrossRef]
- Demarie, S.; Sardella, F.; Billat, V.L.; Magini, W.; Faina, M. The VO2 slow component in swimming. Eur. J. Appl. Physiol. 2001, 84, 95–99. [Google Scholar] [CrossRef]
- Fernandes, R.J.; Cardoso, C.S.; Soares, S.M.; Ascensao, A.; Colaço, P.J.; Vilas-Boas, J.P. Time limit and VO2 slow component at intensities corresponding to VO2max in swimmers. Int. J. Sport. Med. 2003, 24, 576–581. [Google Scholar] [CrossRef]
- Billat, V.L.; Morton, R.H.; Blondel, N.; Berthoin, S.; Bocquet, V.; Koralsztein, J.P.; Barstow, T.J. Oxygen kinetics and modelling of time to exhaustion whilst running at various velocities at maximal oxygen uptake. Eur. J. Appl. Physiol. 2000, 82, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Faina, M.; Billat, V.; Squadrone, R.; Angelis, M. Anaerobic contribution to the time to exhaustion at the minimal exercise intensity at which maximal oxygen uptake occurs in elite cyclist, kaykits and swimmers. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 76, 13–20. [Google Scholar] [CrossRef]
- Almeida, T.A.F.; Pessôa Filho, D.M.; Espada, M.C.; Reis, J.F.; Sancassani, A.; Massini, D.A.; Santos, F.J.; Alves, F.B. Physiological responses during high-intensity interval training in young swimmers. Front. Physiol. 2021, 12, 662029. [Google Scholar] [CrossRef] [PubMed]
- Almeida, T.A.F.; Massini, D.A.; Silva Júnior, O.T.; Venditti Júnior, R.; Espada, M.A.C.; Macedo, A.G.; Reis, J.F.; Alves, F.B.; Pessôa Filho, D.M. Time limit and VO2 kinetics at maximal aerobic velocity: Continuous vs. intermittent swimming trials. Front. Physiol. 2022, 13, 982874. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Figueiredo, P.; Zamparo, P.; Pyne, D.B.; Vilas-Boas, J.P.; Fernandes, R.J. Exercise modality effect on bioenergetical performance at ⩒O2max intensity. Med. Sci. Sport. Exerc. 2015, 47, 1705–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldari, C.; Fernandes, R.J.; Ribeiro, J.; Meucci, M.; Vilas-Boas, J.P.; Guidetti, L. Is the New AquaTrainer® Snorkel Valid for VO2 assessment in swimming? Int. J. Sport. Med. 2013, 34, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Amann, M.; Subudhi, A.; Foster, C. Influence of testing protocol on ventilatory thresholds and cycling performance. Med. Sci. Sport. Exerc. 2004, 36, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Whipp, B.J. Physiological mechanisms dissociating pulmonary CO2 and O2 exchange dynamics during exercise in humans. Exp. Physiol. 2007, 92, 347–355. [Google Scholar] [CrossRef]
- Özyener, F.; Rossiter, H.B.; Ward, S.A.; Whipp, B.J. Influence of exercise intensity on the on- and off-transients kinetics of pulmonary oxygen uptake in humans. J. Physiol. 2001, 533, 891–902. [Google Scholar] [CrossRef]
- DiMenna, F.J.; Jones, A.M. “Linear” versus “Nonlinear” VO2 responses to exercise: Reshaping traditional beliefs. J. Exerc. Sci. Fit. 2009, 7, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Whipp, B.J.; Ward, S.A.; Rossiter, H.B. Pulmonary O2 uptake during exercise: Conflating muscular and cardiovascular responses. Med. Sci. Sport. Exerc. 2005, 37, 1574–1585. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, be-havioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Fritz, C.O.; Morris, P.E.; Richler, J.J. Effect size estimates: Current use, calculations, and interpretation. J. Exp. Psychol. Gen. 2012, 141, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Murgatroyd, S.R.; Ferguson, C.; Ward, S.A.; Whipp, B.J.; Rossiter, H.B. Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J. Appl. Physiol. 2011, 110, 1598–1606. [Google Scholar] [CrossRef] [Green Version]
- Billat, V.; Faina, M.; Sardella, F.; Marini, C.; Fanton, F.; Lupo, S.; Faccini, P.; de Angelis, M.; Koralsztein, J.P.; Dalmonte, A. A Comparison of time to exhaustion at VO2max in élite cyclists, kayak paddlers, swimmers and runners. Ergonomics 1996, 39, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.J.; Keskinen, K.L.; Colaço, P.; Querido, A.J.; Machado, L.; Morais, P.A. Time limit at VO2max velocity in elite crawl swimmers. Int. J. Sport. Med. 2008, 29, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.J.; Billat, V.L.; Cruz, A.C.; Colaço, P.J.; Cardoso, C.S.; Vilas-Boas, J.P. Does net energy cost of swimming affect time to exhaustion at the individual’s maximal oxygen consumption velocity? J. Sport. Med. Phys. Fit. 2006, 46, 373. [Google Scholar]
- di Prampero, P.E. Factors limiting maximal performance in humans. Eur. J. Appl. Physiol. 2003, 90, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Vanhatalo, A.; Burnley, M.; Morton, R.H.; Poole, D.C. Critical power: Implications for determination of VO2max and exercise tolerance. Med. Sci. Sport. Exerc. 2010, 42, 1876–1890. [Google Scholar] [CrossRef]
- Billat, V.L.; Lepretre, P.M.; Heubert, R.P.; Koralsztein, J.P.; Gazeau, F.P. Influence of acute moderate hypoxia on time to exhaustion at vVO2max in unacclimatized runners. Int. J. Sport. Med. 2003, 24, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.D. New ideas on limitations to VO2max. Exerc. Sport. Sci. Rev. 2000, 28, 10–14. [Google Scholar] [PubMed]
- Poole, D.C.; Burnley, M.; Vanhatalo, A.; Rossiter, H.B.; Jones, A.M. Critical power: An important fatigue threshold in exercise physiology. Med. Sci. Sport. Exerc. 2016, 48, 2320–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toussaint, H.M.; Hollander, A.P. Energetics of competitive swimming. Sport. Med. 1994, 18, 384–405. [Google Scholar] [CrossRef]
- Sousa, A.; Figueiredo, P.; Pendergast, D.; Kjendlie, P.L.; Vilas-Boas, J.P.; Fernandes, R.J. critical evaluation of oxygen-uptake assessment in swimming. Int. J. Sport. Physiol. Perf. 2014, 9, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Billat, V.L.; Mille-Hamard, L.; Demarle, A.; Koralsztein, J.P. Effect of Training in humans on off- and on-transient oxygen uptake kinetics after severe exhausting intensity runs. Eur. J. Appl. Physiol. 2002, 87, 496–505. [Google Scholar] [CrossRef]
- Burnley, M.; Jones, A.M. Oxygen uptake kinetics as a determinant of sports performance. Eur. J. Sport. Sci. 2007, 7, 63–79. [Google Scholar] [CrossRef]
- Massini, D.A.; Almeida, T.A.F.; Vasconcelos, C.M.T.; Macedo, A.G.; Espada, M.A.C.; Reis, J.F.; Alves, F.J.B.; Fernandes, R.J.P.; Pessôa Filho, D.M. Are young swimmers short and middle distances energy cost sex-specific? Front. Physiol. 2021, 12, 796886. [Google Scholar] [CrossRef]
- Sokołowski, K.; Strzała, M.; Stanula, A.; Kryst, Ł.; Radecki-Pawlik, A.; Krężałek, P.; Rosemann, T.; Knechtle, B. Biological age in relation to somatic, physiological, and swimming kinematic indices as predictors of 100 m front crawl performance in young female swimmers. Int. J. Environ. Res. Public Health 2021, 18, 6062. [Google Scholar] [CrossRef]
- Sokołowski, K.; Bartolomeu, R.F.; Barbosa, T.M.; Strzała, M. VO2 kinetics and tethered strength influence the 200-m front crawl stroke kinematics and speed in young male swimmers. Front. Physiol. 2022, 13, 1045178. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Men | Women | |
|---|---|---|---|
| VO2b (ml·min−1) | 665.8 ± 148.7 | 684.2 ± 146.5 | 625.2 ± 162.1 |
| TD1 (s) | 17.7 ± 5.1 | 17.9 ± 5.1 | 17.2 ± 5.5 |
| τ1 (s) | 24.4 ± 9.8 | 25.5 ± 11.7 | 22.2 ± 3.9 |
| A1′ (ml·min−1) | 3115.2 ± 497.4 | 3368.6 ± 360.7 | 2557.9 ± 193.6 |
| R2 | 0.98 ± 0.02 | 0.97 ± 0.02 | 0.98 ± 0.00 |
| TD2 (s) | 149.3 ± 29.1 | 141.7 ± 26.5 | 166.0 ± 30.1 |
| SC (ml·min−1) | 333.6 ± 211.2 | 414.1 ± 215.6 | 172.6 ± 53.6 |
| SC (%) | 9.4 ± 5.1 | 11.0 ± 5.5 | 6.4 ± 2.4 |
| EEVO2 (ml·min−1) | 3427.6 ± 565.4 | 3744.5 ± 341.1 | 2730.5 ± 155.5 |
| VO2peak (%) | 100.4 ± 3.8 | 100.6 ± 4.2 | 100.1 ± 3.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massini, D.A.; Espada, M.C.; Macedo, A.G.; Santos, F.J.; Castro, E.A.; Ferreira, C.C.; Robalo, R.A.M.; Dias, A.A.P.; Almeida, T.A.F.; Pessôa Filho, D.M. Oxygen Uptake Kinetics and Time Limit at Maximal Aerobic Workload in Tethered Swimming. Metabolites 2023, 13, 773. https://doi.org/10.3390/metabo13070773
Massini DA, Espada MC, Macedo AG, Santos FJ, Castro EA, Ferreira CC, Robalo RAM, Dias AAP, Almeida TAF, Pessôa Filho DM. Oxygen Uptake Kinetics and Time Limit at Maximal Aerobic Workload in Tethered Swimming. Metabolites. 2023; 13(7):773. https://doi.org/10.3390/metabo13070773
Chicago/Turabian StyleMassini, Danilo A., Mário C. Espada, Anderson G. Macedo, Fernando J. Santos, Eliane A. Castro, Cátia C. Ferreira, Ricardo A. M. Robalo, Amândio A. P. Dias, Tiago A. F. Almeida, and Dalton M. Pessôa Filho. 2023. "Oxygen Uptake Kinetics and Time Limit at Maximal Aerobic Workload in Tethered Swimming" Metabolites 13, no. 7: 773. https://doi.org/10.3390/metabo13070773
APA StyleMassini, D. A., Espada, M. C., Macedo, A. G., Santos, F. J., Castro, E. A., Ferreira, C. C., Robalo, R. A. M., Dias, A. A. P., Almeida, T. A. F., & Pessôa Filho, D. M. (2023). Oxygen Uptake Kinetics and Time Limit at Maximal Aerobic Workload in Tethered Swimming. Metabolites, 13(7), 773. https://doi.org/10.3390/metabo13070773